
Flavonoids Accumulation in Fruit Peel and Expression Profiling of Related Genes in Purple (Passiflora edulis f. edulis) and Yellow (Passiflora edulis f. flavicarpa) Passion Fruits

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
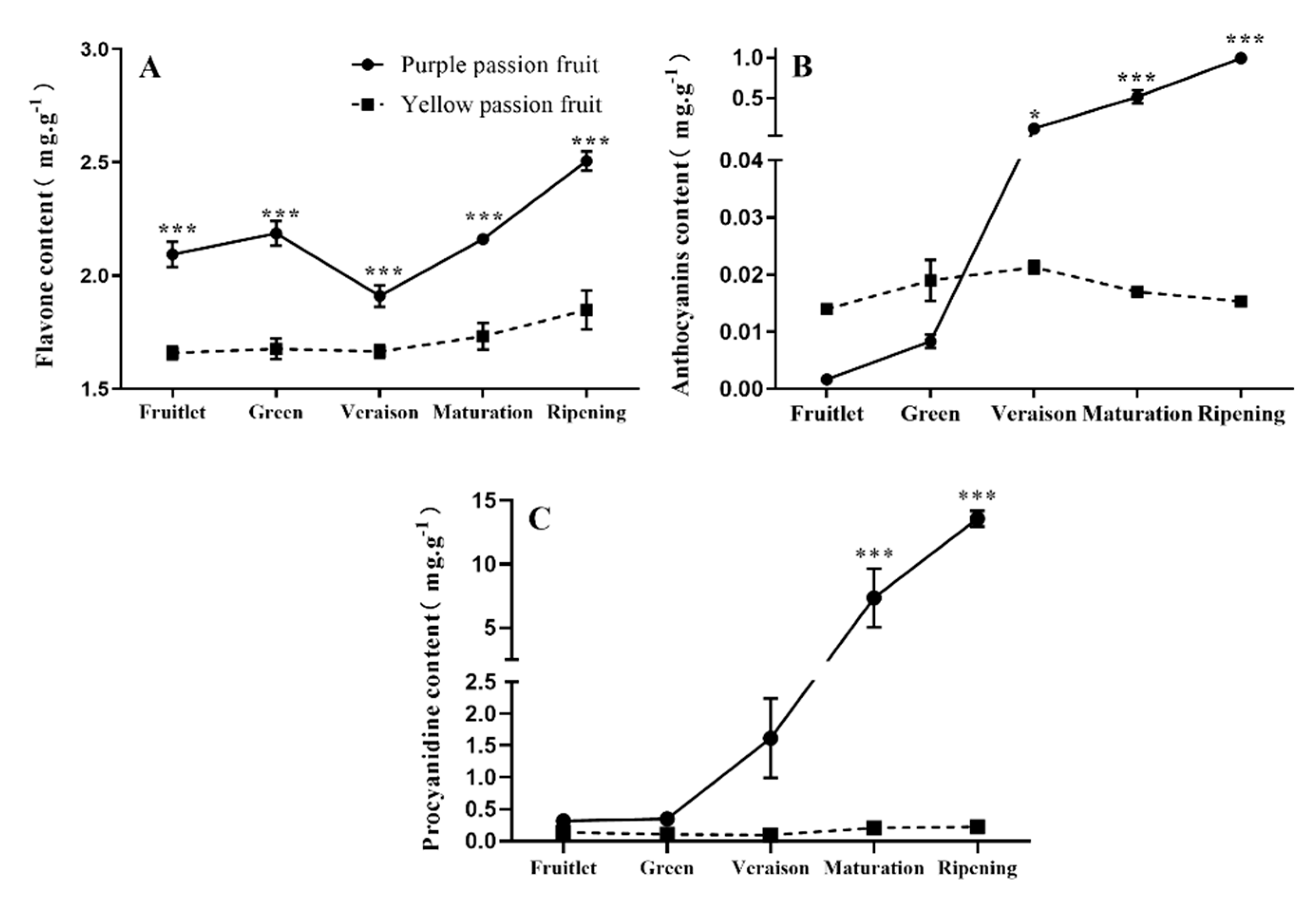
2.1. Total Flavonoids, Anthocyanins, and Procyanidins
2.2. Flavonoid and Anthocyanin Metabolites
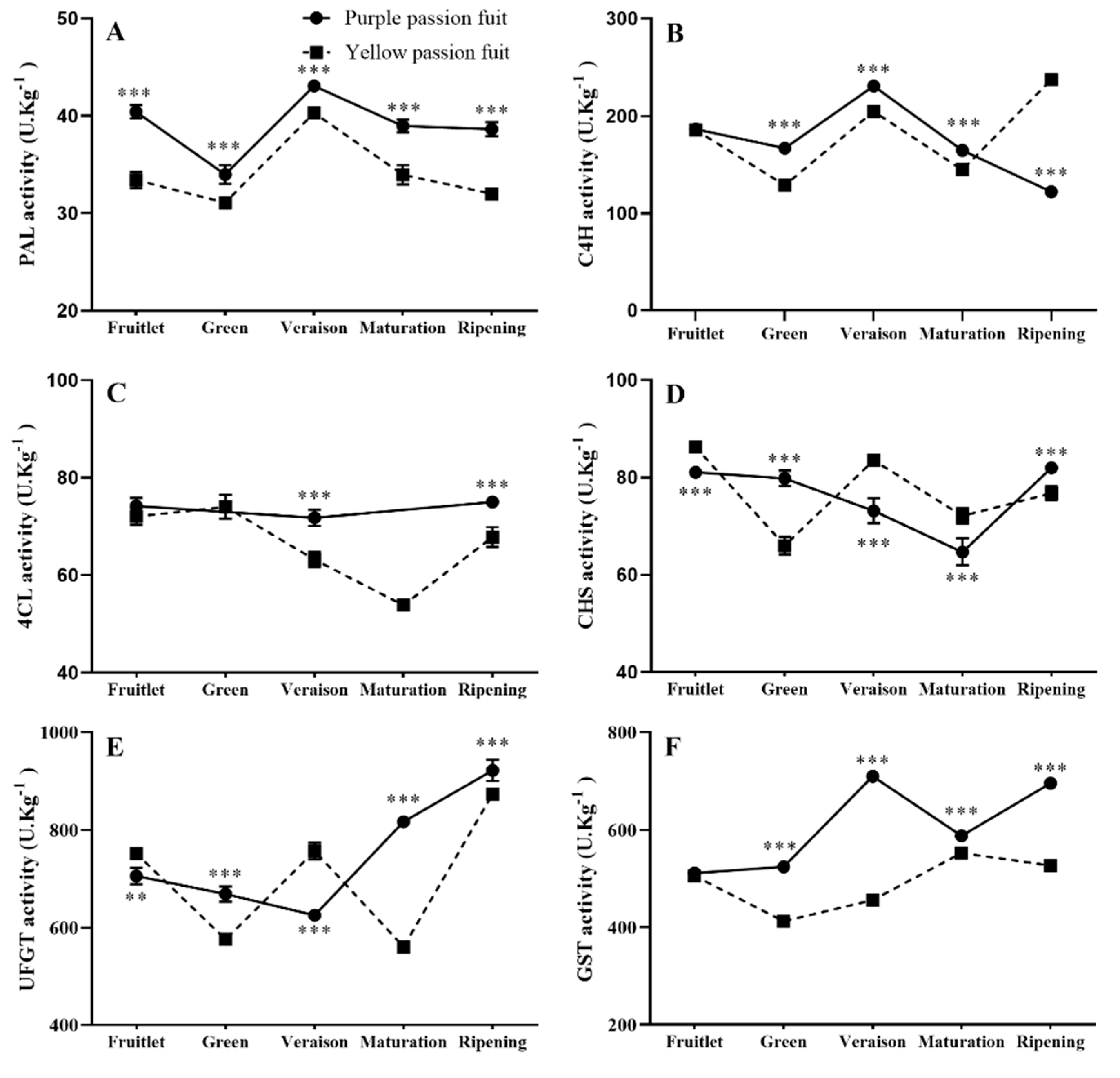
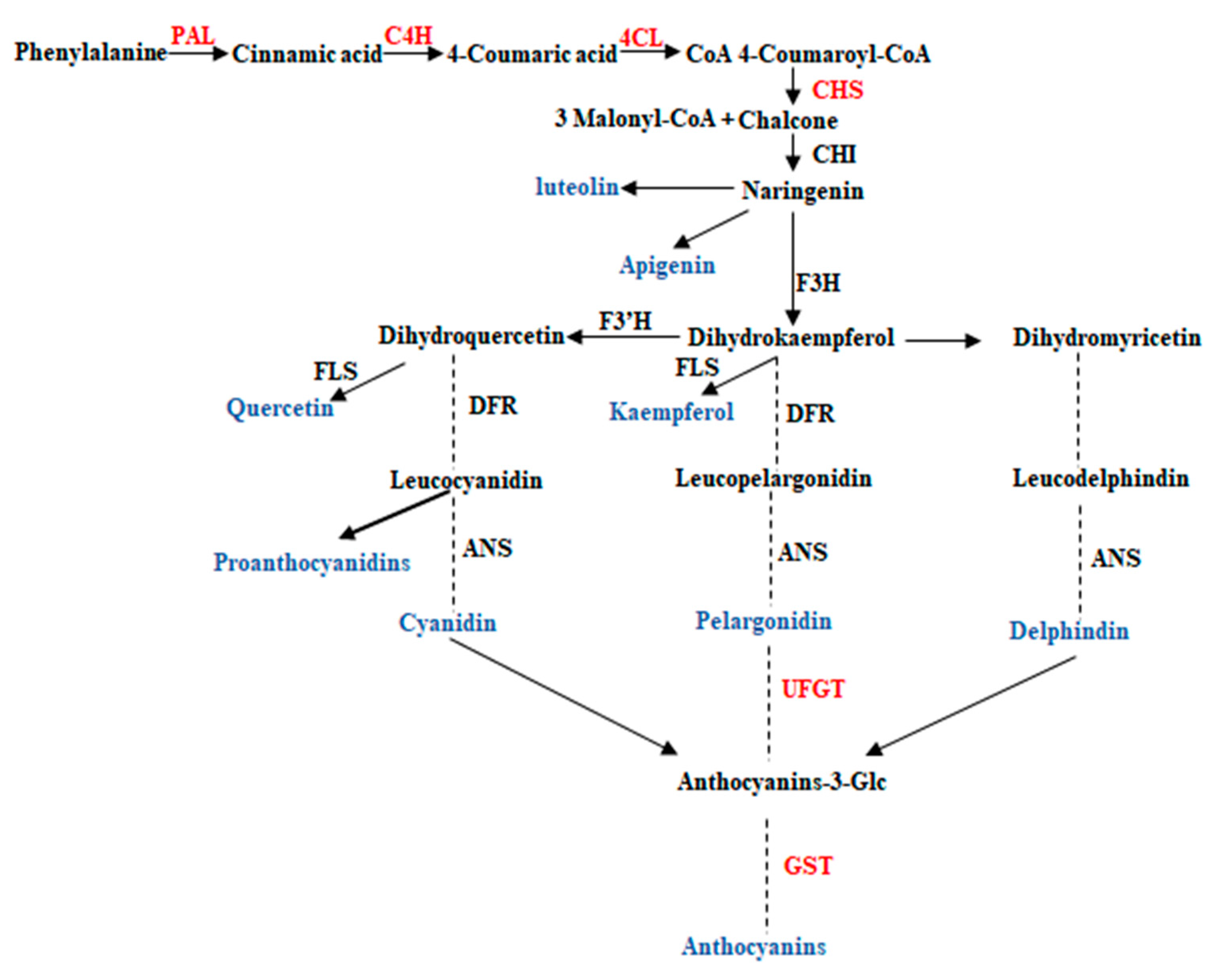
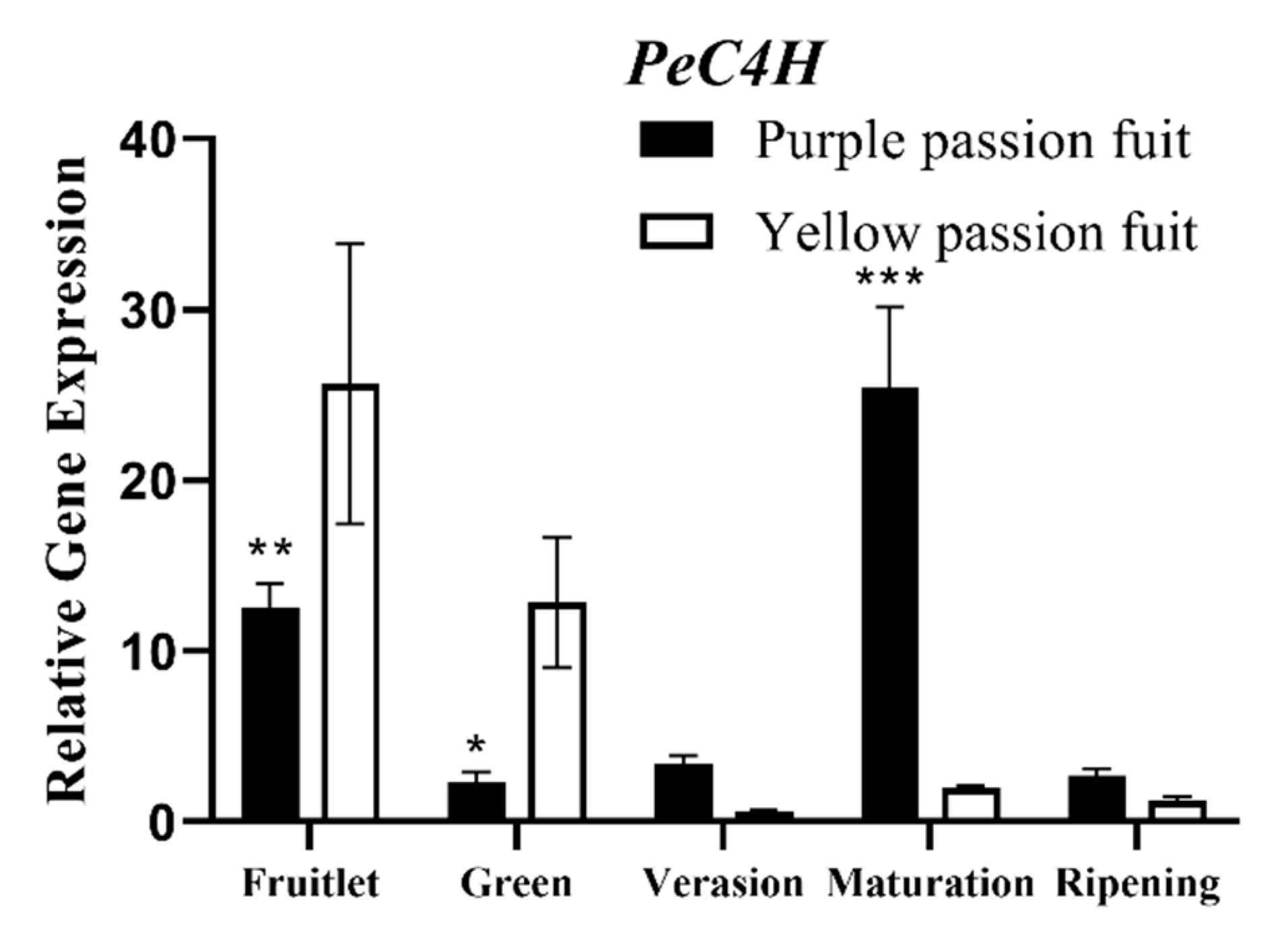
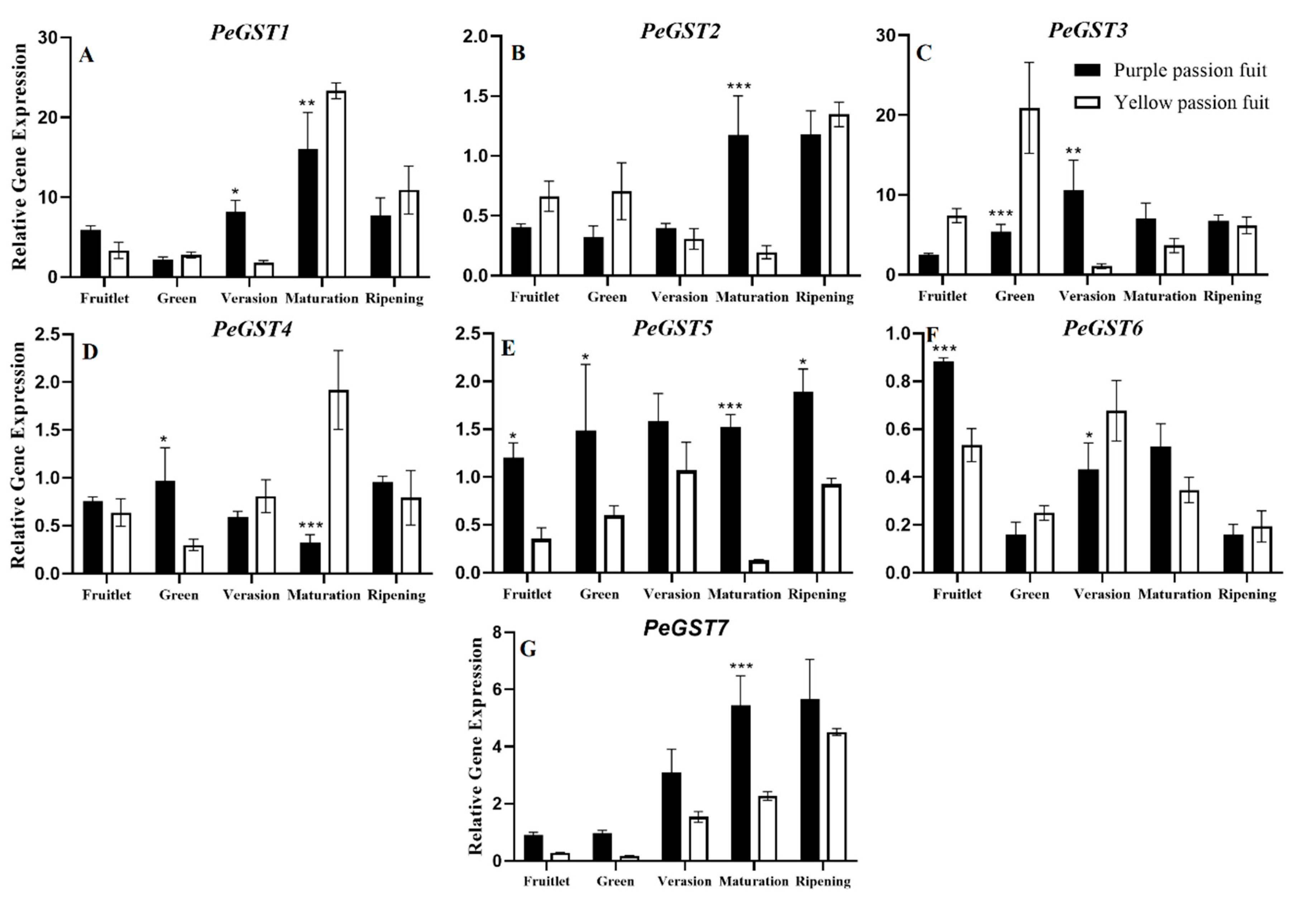
2.3. Key Enzymes Involved in Flavonoids Metabolism
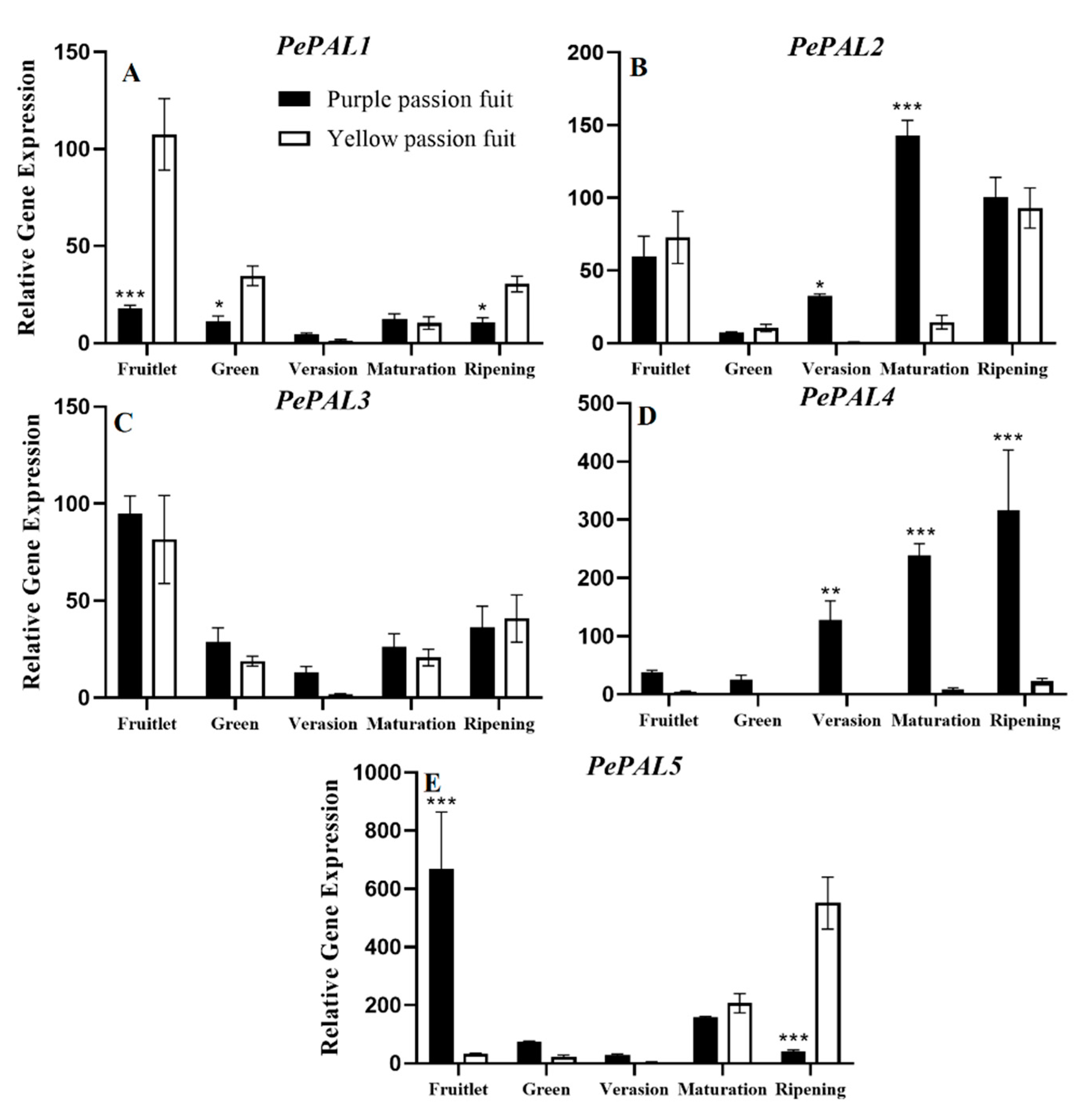
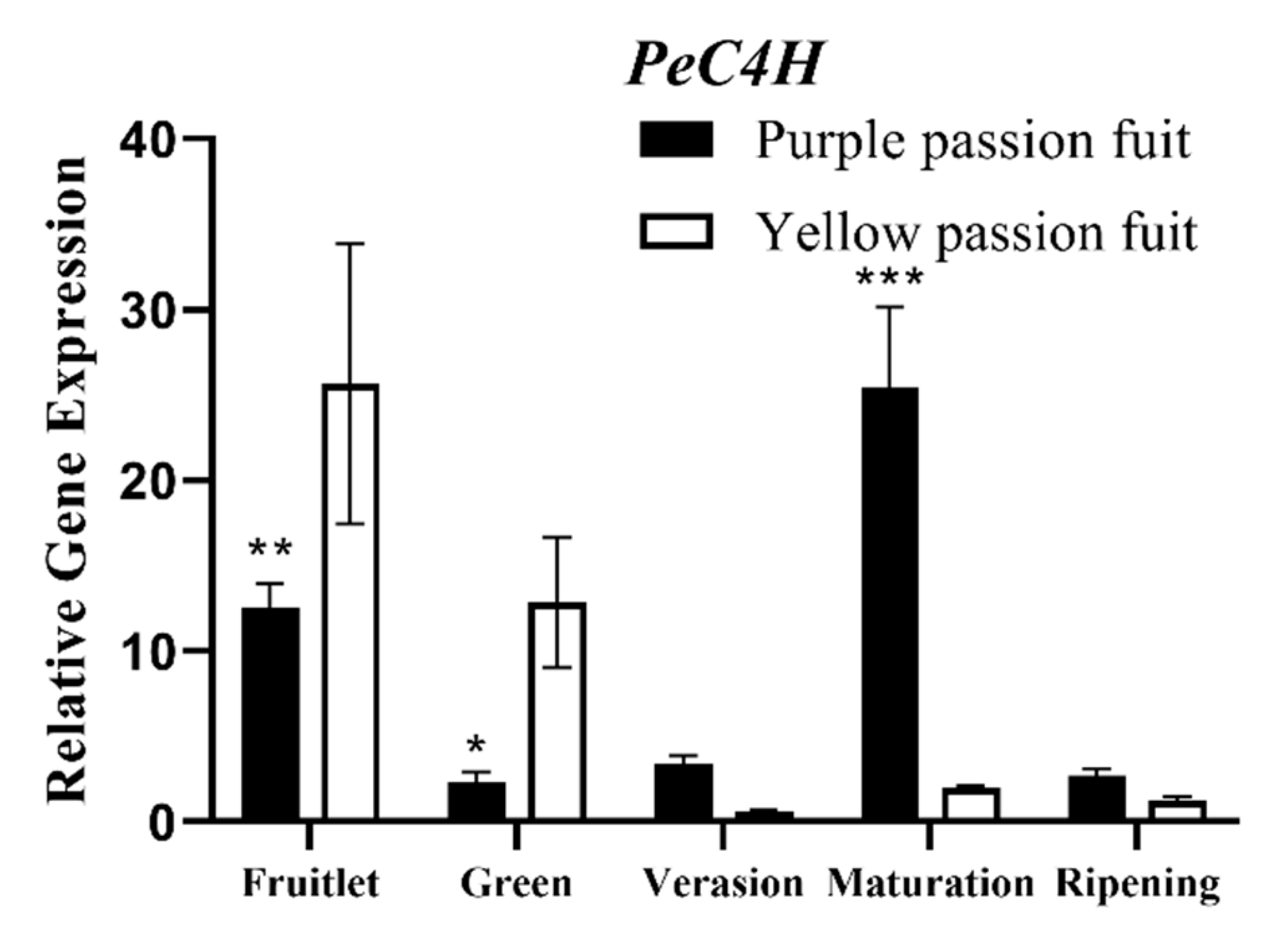
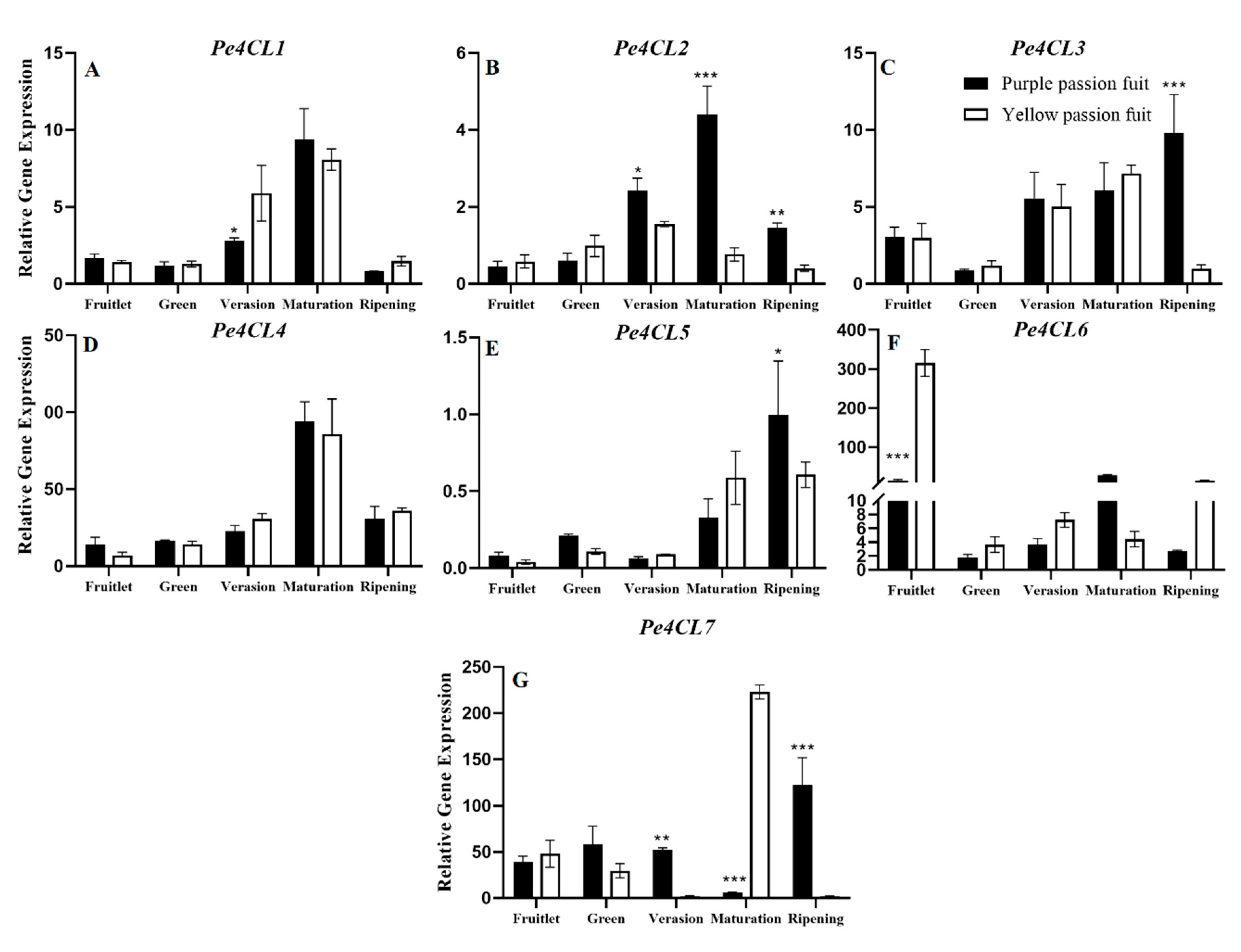
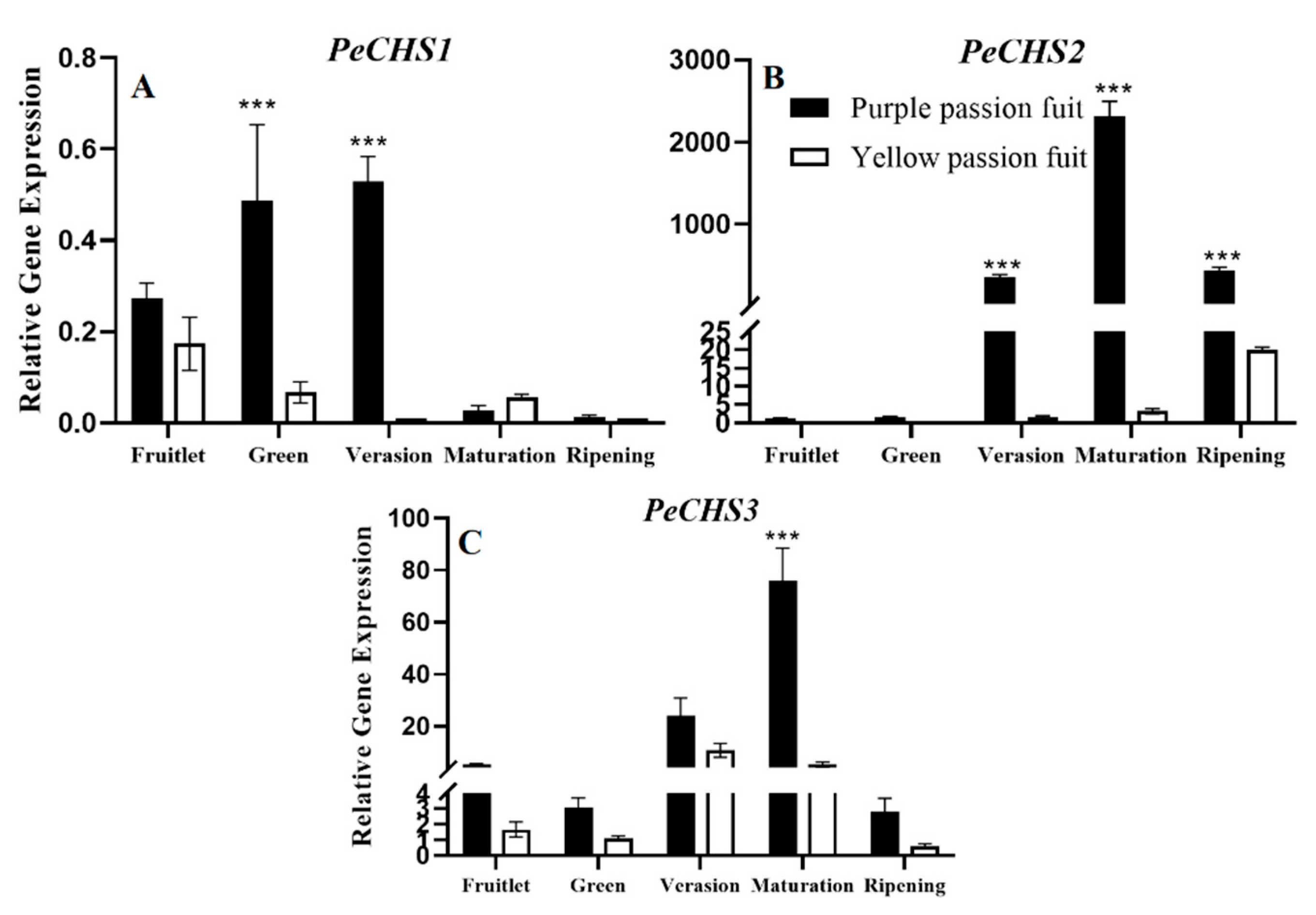
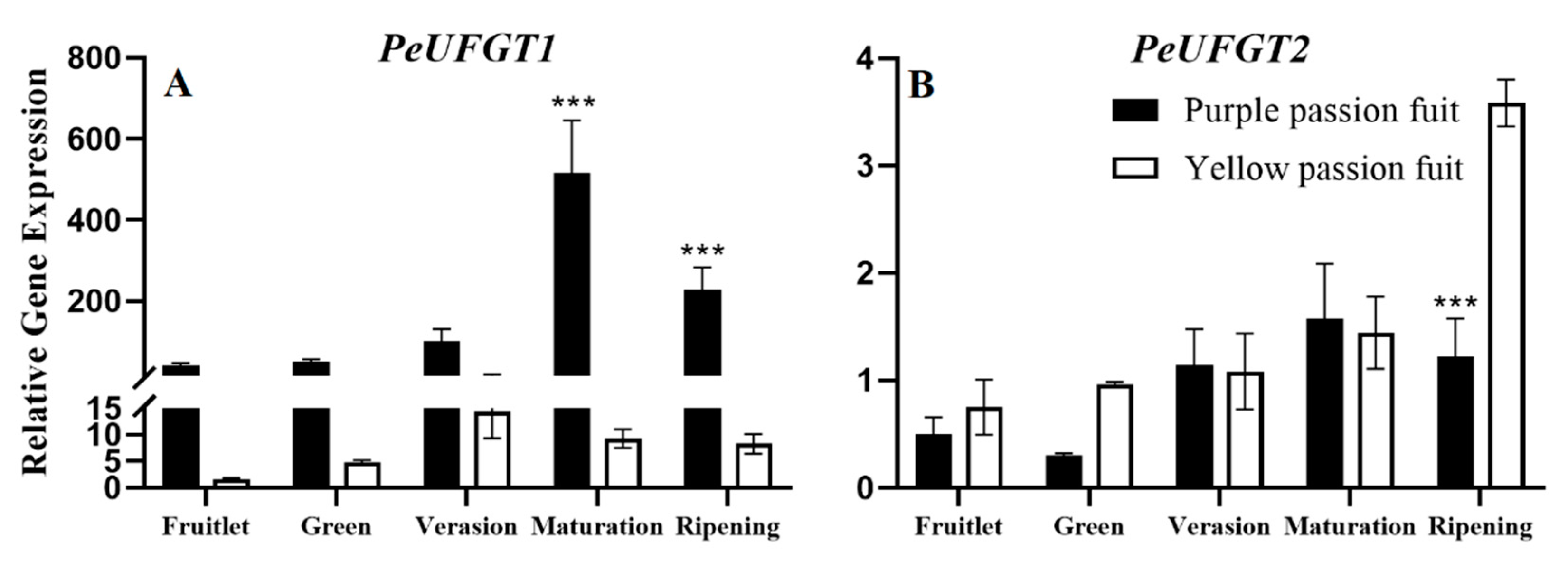
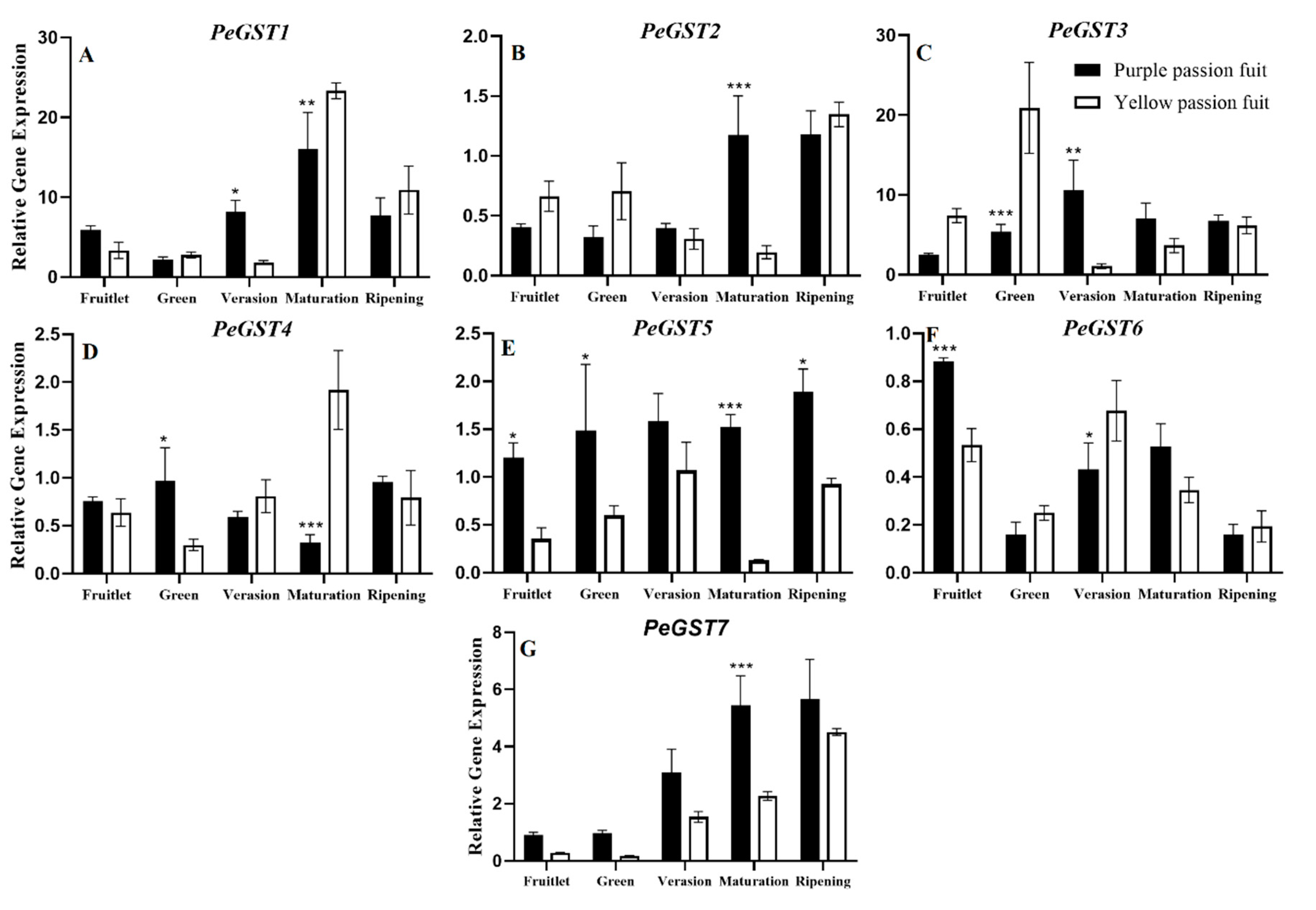
2.4. Expression Profiling of Genes Encoding Key Enzymes for Flavonoid Metabolism
2.5. Principle Component Analysis
2.5.1. Purple Passion Fruit
2.5.2. Yellow Passion Fruit
3. Discussion
4. Materials and Methods
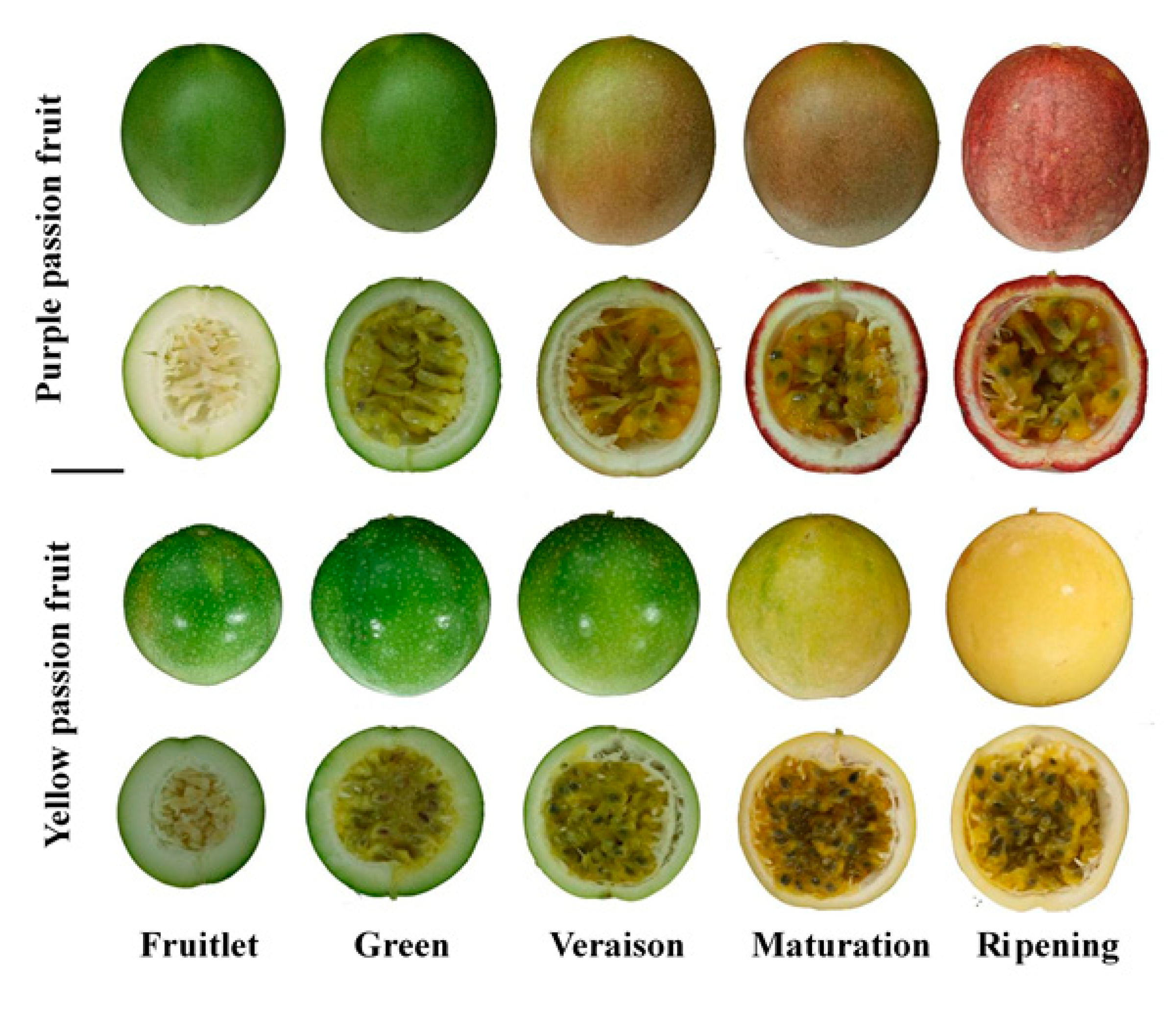
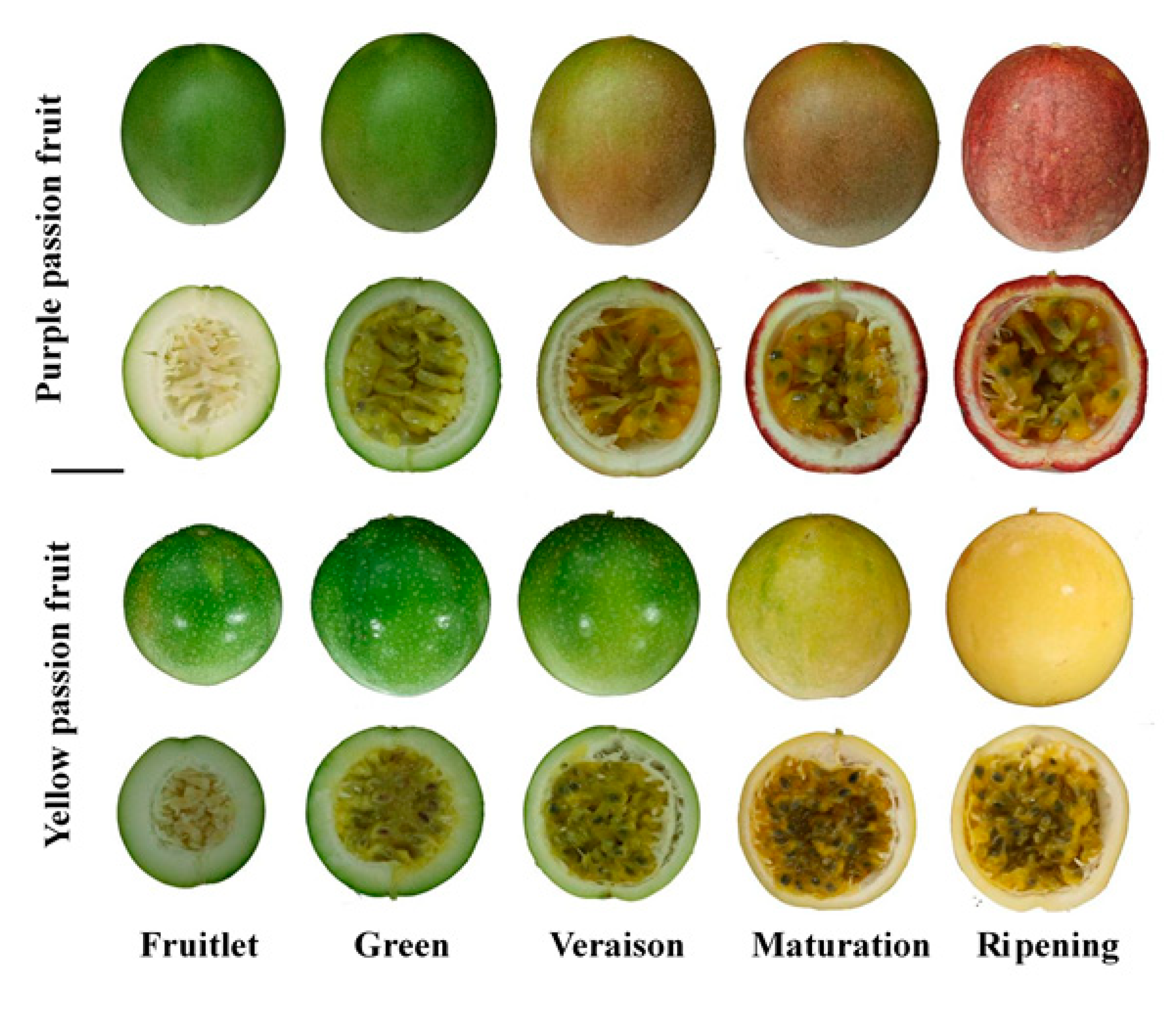
4.1. Plant Material
4.2. Determination of Total Flavonoids, Anthocyanins, and Proanthocyanins
4.3. Determination of Flavonoid and Anthocyanin Metabolites
4.4. Enzymes Extraction and Activity Assay
4.5. RNA Extraction and Real-Time Quantitative PCR
4.6. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rodriguez-Amaya, D.B. Passion Fruits. In Encyclopedia of Food Sciences and Nutrition; Academic Press: Cambridge, MA, USA, 2003; pp. 4368–4373. [Google Scholar]
- Huo, D.; Lan, J.; Ma, L.-L.; Hou, C.-J.; Yang, P. Function of Passiflor and its comprehensive processing utility. Sci. Technol. Food Ind. 2012, 19, 391–395. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, X.; Ali, M.M.; Rizwan, H.M.; Li, B.; Li, H.; Jia, K.; Yang, X.; Ma, S.; Li, S.; et al. Changes in the Content of Organic Acids and Expression Analysis of Citric Acid Accumulation-Related Genes during Fruit Development of Yellow (Passiflora edulis f. flavicarpa) and Purple (Passiflora edulis f. edulis) Passion Fruits. Int. J. Mol. Sci. 2021, 22, 5765. [Google Scholar] [CrossRef]
- Sousa, D.F.; Veras, V.S.; Freire, V.E.C.S.; Paula, M.L.; Serra, M.A.A.O.; Costa, A.C.P.J.; da Conceição, S.O. Cunha, M.; Queiroz, M.V.O.; Damasceno, M.M.C.; et al. Effectiveness of Passion Fruit Peel Flour (Passiflora edulis L.) versus Turmeric Flour (Curcuma longa L.) on Glycemic Control: Systematic Review and Meta-Analysis. Curr. Diabetes Rev. 2020, 16, 450–456. [Google Scholar] [CrossRef]
- Konta, E.M.; Almeida, M.R.; do Amaral, C.L.; Darin, J.D.C.; de Rosso, V.V.; Mercadante, A.Z.; Antunes, L.M.G.; Bianchi, M.L.P. Evaluation of the Antihypertensive Properties of Yellow Passion Fruit Pulp (Passiflora edulis Sims f. flavicarpa Deg.) in Spontaneously Hypertensive Rats. Phytother. Res. 2014, 28, 28–32. [Google Scholar] [CrossRef]
- Lewis, B.J.; Herrlinger, K.A.; Craig, T.A.; Mehring-Franklin, C.E.; DeFreitas, Z.; Hinojosa-Laborde, C. Antihypertensive effect of passion fruit peel extract and its major bioactive components following acute supplementation in spontaneously hypertensive rats. J. Nutr. Biochem. 2013, 24, 1359–1366. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima, G.C.; Batista, Â.G.; Carazin, C.B.B.; Cintra, D.E.; Prado, M.A.; Júnior, M.R.M. Passion fruit peel intake decreases inflammatory response and reverts lipid peroxidation and adiposity in diet-induced obese rats. Nutr. Res. 2020, 76, 106–117. [Google Scholar] [CrossRef]
- Nerdy, N.; Ritarwan, K. Hepatoprotective Activity and Nephroprotective Activity of Peel Extract from Three Varieties of the Passion Fruit (Passiflora Sp.) in the Albino Rat. Open Access Maced. J. Med. Sci. 2019, 7, 536–542. [Google Scholar] [CrossRef] [Green Version]
- Prasertsri, P.; Booranasuksakul, U.; Naravoratham, K.; Trongtosak, P. Acute Effects of Passion Fruit Juice Supplementation on Cardiac Autonomic Function and Blood Glucose in Healthy Subjects. Prev. Nutr. Food Sci. 2019, 24, 245–253. [Google Scholar] [CrossRef]
- Garcia, M.V.; Milani, M.S.; Ries, E.F. Production optimization of passion fruit peel flour and its incorporation into dietary food. Food Sci. Technol. Int. 2020, 26, 132–139. [Google Scholar] [CrossRef]
- Viganó, J.; Meirelles, A.A.D.; Náthia-Neves, G.; Baseggio, A.M.; Cunha, R.L.; Maróstica, M.R., Jr.; Meireles, M.A.A.; Gurikov, P.; Smirnova, I.; Martínez, J. Impregnation of passion fruit bagasse extract in alginate aerogel microparticles. Int. J. Biol. Macromol. 2020, 155, 1060–1068. [Google Scholar] [CrossRef]
- Liang, D.; Ali, M.M.; Yousef, A.F.; He, Y.; Huang, X.; Li, J.; Yang, Q.; Chen, F. Root Colonization of Piriformospora indica Improves Phyto-Nutritional Composition of Leaves, Stems, Tendrils and Fruits of Passiflora edulis f. edulis. Adv. Food Sci. 2021, 43, 142–149. [Google Scholar]
- Ali, M.M.; Anwar, R.; Yousef, A.F.; Li, B.; Luvisi, A.; de Bellis, L.; Aprile, A.; Chen, F. Influence of Bagging on the Development and Quality of Fruits. Plants 2021, 10, 358. [Google Scholar] [CrossRef]
- Caltagirone, S.; Rossi, C.; Poggi, A.; Ranelletti, F.O.; Natali, P.G.; Brunetti, M.; Aiello, F.B.; Piantelli, M. Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int. J. Cancer 2000, 87, 595–600. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Cao, Y.; Han, Y.; Li, D.; Lin, Y.; Cai, Y. Systematic Analysis of the 4-Coumarate:Coenzyme A Ligase (4CL) Related Genes and Expression Profiling during Fruit Development in the Chinese Pear. Genes 2016, 7, 89. [Google Scholar] [CrossRef] [Green Version]
- Haskill, J.S.; Häyry, P.; Radov, L.A. Systemic and local immunity in allograft and cancer rejection. Contemp. Top. Immunobiol. 1978, 8, 107–170. [Google Scholar] [CrossRef]
- Fu, H.; Qiao, Y.; Wang, P.; Mu, X.; Zhang, J.; Fu, B.; Du, J. Changes of bioactive components and antioxidant potential during fruit development of Prunus humilis Bunge. PLoS ONE 2021, 16, e0251300. [Google Scholar] [CrossRef]
- Lister, C.E.; Lancaster, J.E.; Sutton, K.H.; Walker, J.R.L. Developmental changes in the concentration and composition of flavonoids in skin of a red and a green apple cultivar. J. Sci. Food Agric. 1994, 64, 155–161. [Google Scholar] [CrossRef]
- Zoratti, L.; Karppinen, K.; Luengo Escobar, A.; Häggman, H.; Jaakola, L. Light-controlled flavonoid biosynthesis in fruits. Front. Plant Sci. 2014, 5, 534. [Google Scholar] [CrossRef]
- Qian, Y.; Lynch, J.H.; Guo, L.; Rhodes, D.; Morgan, J.A.; Dudareva, N. Completion of the cytosolic post-chorismate phenylalanine biosynthetic pathway in plants. Nat. Commun. 2019, 10, 15. [Google Scholar] [CrossRef]
- Kong, J.-Q. Phenylalanine ammonia-lyase, a key component used for phenylpropanoids production by metabolic engineering. RSC Adv. 2015, 5, 62587–62603. [Google Scholar] [CrossRef]
- Singh, R.; Rastogi, S.; Dwivedi, U.N. Phenylpropanoid Metabolism in Ripening Fruits. Compr. Rev. Food Sci. Food Saf. 2010, 9, 398–416. [Google Scholar] [CrossRef]
- Lu, Y.; Song, Y.; Zhu, J.; Xu, X.; Pang, B.; Jin, H.; Jiang, C.; Liu, Y.; Shi, J. Potential application of CHS and 4CL genes from grape endophytic fungus in production of naringenin and resveratrol and the improvement of polyphenol profiles and flavour of wine. Food Chem. 2021, 347, 128972. [Google Scholar] [CrossRef]
- Flores-Sanchez, I.J.; Verpoorte, R. Plant Polyketide Synthases: A fascinating group of enzymes. Plant Physiol. Biochem. 2009, 47, 167–174. [Google Scholar] [CrossRef]
- Li, X.-J.; Zhang, J.-Q.; Wu, Z.-C.; Lai, B.; Huang, X.-M.; Qin, Y.-H.; Wang, H.-C.; Hu, G.-B. Functional characterization of a glucosyltransferase gene, LcUFGT1, involved in the formation of cyanidin glucoside in the pericarp of Litchi chinensis. Physiol. Plant. 2016, 156, 139–149. [Google Scholar] [CrossRef]
- Vilperte, V.; Boehm, R.; Debener, T. A highly mutable GST is essential for bract colouration in Euphorbia pulcherrima Willd. Ex Klotsch. BMC Genom. 2021, 22, 208. [Google Scholar] [CrossRef]
- Rinaldi, M.M.; Costa, A.M.; Faleiro, F.G.; Junqueira, N.T.V. Conservação pós-colheita de frutos de Passiflora setacea DC. submetidos a diferentes sanitizantes e temperaturas de armazenamento. Braz. J. Food Technol. 2017, 20, e2016046. [Google Scholar] [CrossRef]
- Maniwara, P.; Nakano, K.; Boonyakiat, D.; Ohashi, S.; Hiroi, M.; Tohyama, T. The use of visible and near infrared spectroscopy for evaluating passion fruit postharvest quality. J. Food Eng. 2014, 143, 33–43. [Google Scholar] [CrossRef]
- Oluoch, P.; Nyaboga, E.N.; Bargul, J.L. Analysis of genetic diversity of passion fruit (Passiflora edulis Sims) genotypes grown in Kenya by sequence-related amplified polymorphism (SRAP) markers. Ann. Agrar. Sci. 2018, 16, 367–375. [Google Scholar] [CrossRef]
- Hyson, D.A. A Comprehensive Review of Apples and Apple Components and Their Relationship to Human Health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef]
- Wang, C.-H.; Yu, J.; Cai, Y.-X.; Zhu, P.-P.; Liu, C.-Y.; Zhao, A.-C.; Lü, R.-H.; Li, M.-J.; Xu, F.-X.; Yu, M.-D. Characterization and Functional Analysis of 4-Coumarate:CoA Ligase Genes in Mulberry. PLoS ONE 2016, 11, e0155814. [Google Scholar] [CrossRef]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef]
- Falginella, L.; Di Gaspero, G.; Castellarin, S.D. Expression of flavonoid genes in the red grape berry of “Alicante Bouschet” varies with the histological distribution of anthocyanins and their chemical composition. Planta 2012, 236, 1037–1051. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Ferreres, F.; Sousa, C.; Valentão, P.; Andrade, P.B.; Seabra, R.M.; Gil-Izquierdo, Á. New C-Deoxyhexosyl Flavones and Antioxidant Properties of Passiflora edulis Leaf Extract. J. Agric. Food Chem. 2007, 55, 10187–10193. [Google Scholar] [CrossRef]
- Domínguez-Rodríguez, G.; García, M.C.; Plaza, M.; Marina, M.L. Revalorization of Passiflora species peels as a sustainable source of antioxidant phenolic compounds. Sci. Total Environ. 2019, 696, 134030. [Google Scholar] [CrossRef]
- Zeraik, M.L.; Serteyn, D.; Deby-Dupont, G.; Wauters, J.-N.; Tits, M.; Yariwake, J.H.; Angenot, L.; Franck, T. Evaluation of the antioxidant activity of passion fruit (Passiflora edulis and Passiflora alata) extracts on stimulated neutrophils and myeloperoxidase activity assays. Food Chem. 2011, 128, 259–265. [Google Scholar] [CrossRef]
- Memon, A.F.; Solangi, A.R.; Memon, S.Q.; Mallah, A.; Memon, N.; Memon, A.A. Simultaneous Determination of Quercetin, Rutin, Naringin, and Naringenin in Different Fruits by Capillary Zone Electrophoresis. Food Anal. Methods 2017, 10, 83–91. [Google Scholar] [CrossRef]
- Balbaa, S.I.; Zaki, A.Y.; El Shamy, A.M. Total Flavonoid and Rutin Content of the Different Organs of Sophora japonica L. J. Assoc. Off. Anal. Chem. AOAC Int. 1974, 57, 752–755. [Google Scholar] [CrossRef]
- Yang, Z.; Zhai, W. Optimization of microwave-assisted extraction of anthocyanins from purple corn (Zea mays L.) cob and identification with HPLC–MS. Innov. Food Sci. Emerg. Technol. 2010, 11, 470–476. [Google Scholar] [CrossRef]
- Jaiswal, V.; DerMarderosian, A.; Porter, J.R. Anthocyanins and polyphenol oxidase from dried arils of pomegranate (Punica granatum L.). Food Chem. 2010, 118, 11–16. [Google Scholar] [CrossRef]
- Argentieri, M.P.; Levi, M.; Guzzo, F.; Avato, P. Phytochemical analysis of Passiflora loefgrenii Vitta, a rich source of luteolin-derived flavonoids with antioxidant properties. J. Pharm. Pharmacol. 2015, 67, 1603–1612. [Google Scholar] [CrossRef]
- Waki, T.; Mameda, R.; Nakano, T.; Yamada, S.; Terashita, M.; Ito, K.; Tenma, N.; Li, Y.; Fujino, N.; Uno, K.; et al. A conserved strategy of chalcone isomerase-like protein to rectify promiscuous chalcone synthase specificity. Nat. Commun. 2020, 11, 870. [Google Scholar] [CrossRef]
- Liou, G.; Chiang, Y.-C.; Wang, Y.; Weng, J.-K. Mechanistic basis for the evolution of chalcone synthase catalytic cysteine reactivity in land plants. J. Biol. Chem. 2018, 293, 18601–18612. [Google Scholar] [CrossRef] [Green Version]
- Jinxia, H.; Lijia, Q.; Ji, Y.; Hao, Y.; Hongya, G. A preliminary study on the origin and evolution of chalcone synthase (CHS) gene in angiosperms. Acta Bot. Sin. 2004, 46, 10–19. [Google Scholar]
- Holton, T.A.; Cornish, E.C. Genetics and Biochemistry of Anthocyanin Biosynthesis. Plant Cell 1995, 7, 1071–1083. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä, K.; Pirttilä, A.M.; Törrönen, R.; Kärenlampi, S.; Hohtola, A. Expression of Genes Involved in Anthocyanin Biosynthesis in Relation to Anthocyanin, Proanthocyanidin, and Flavonol Levels during Bilberry Fruit Development. Plant Physiol. 2002, 130, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.; Jiang, H.-L.; Lei, B.; Li, J.-N.; Chai, Y.-R. Molecular cloning, characterization and expression of two rapeseed (Brassica napus L.) cDNAs orthologous to Arabidopsis thaliana phenylalanine ammonia-lyase 1. Euphytica 2008, 159, 1–16. [Google Scholar] [CrossRef]
- Bai, C.; Xu, J.; Cao, B.; Li, X.; Li, G. Transcriptomic analysis and dynamic expression of genes reveal flavonoid synthesis in Scutellaria viscidula. Acta Physiol. Plant. 2018, 40, 161. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Li, C.; Duan, C.; Gu, C.; Zhang, Q. Integrated Metabolomic and Transcriptomic Analysis Reveals the Flavonoid Regulatory Network by Eutrema EsMYB90. Int. J. Mol. Sci. 2021, 22, 8751. [Google Scholar] [CrossRef]
- Zhao, X.; Yuan, Z.; Feng, L.; Fang, Y. Cloning and expression of anthocyanin biosynthetic genes in red and white pomegranate. J. Plant Res. 2015, 128, 687–696. [Google Scholar] [CrossRef]
- Boss, P.K.; Davies, C.; Robinson, S.P. Analysis of the Expression of Anthocyanin Pathway Genes in Developing Vitis vinifera L. cv Shiraz Grape Berries and the Implications for Pathway Regulation. Plant Physiol. 1996, 111, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Kondo, S.; Hiraoka, K.; Kobayashi, S.; Honda, C.; Terahara, N. Changes in the Expression of Anthocyanin Biosynthetic Genes during Apple Development. J. Am. Soc. Hortic. Sci. 2002, 127, 971–976. [Google Scholar] [CrossRef]
- Zhai, R.; Wang, Z.; Zhang, S.; Meng, G.; Song, L.; Wang, Z.; Li, P.; Ma, F.; Xu, L. Two MYB transcription factors regulate flavonoid biosynthesis in pear fruit (Pyrus bretschneideri Rehd.). J. Exp. Bot. 2016, 67, 1275–1284. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, Y.-Y.; Liu, H.; Zhang, X.-S.; Ni, R.; Wang, P.-Y.; Gao, S.; Lou, H.-X.; Cheng, A.-X. Functional characterization of a liverworts bHLH transcription factor involved in the regulation of bisbibenzyls and flavonoids biosynthesis. BMC Plant Biol. 2019, 19, 497. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Y.; Coulter, J.A.; Shen, B.; Li, Y.; Li, C.; Cao, Z.; Zhang, J. The WD40 Gene Family in Potato (Solanum Tuberosum L.): Genome-Wide Analysis and Identification of Anthocyanin and Drought-Related WD40s. Agronomy 2020, 10, 401. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Hwang, G.; Lee, S.; Zhu, J.-Y.; Paik, I.; Nguyen, T.T.; Kim, J.; Oh, E. High Ambient Temperature Represses Anthocyanin Biosynthesis through Degradation of HY5. Front. Plant Sci. 2017, 8, 1787. [Google Scholar] [CrossRef] [Green Version]
- Vinatoru, M.; Mason, T.J.; Calinescu, I. Ultrasonically assisted extraction (UAE) and microwave assisted extraction (MAE) of functional compounds from plant materials. TrAC Trends Anal. Chem. 2017, 97, 159–178. [Google Scholar] [CrossRef]
- Kim, I.; Lee, J. Variations in Anthocyanin Profiles and Antioxidant Activity of 12 Genotypes of Mulberry (Morus spp.) Fruits and Their Changes during Processing. Antioxidants 2020, 9, 242. [Google Scholar] [CrossRef] [Green Version]
- Hellström, J.K.; Mattila, P.H. HPLC Determination of Extractable and Unextractable Proanthocyanidins in Plant Materials. J. Agric. Food Chem. 2008, 56, 7617–7624. [Google Scholar] [CrossRef]
- Henry-Kirk, R.A.; Plunkett, B.; Hall, M.; McGhie, T.; Allan, A.C.; Wargent, J.J.; Espley, R.V. Solar UV light regulates flavonoid metabolism in apple (Malus x domestica). Plant. Cell Environ. 2018, 41, 675–688. [Google Scholar] [CrossRef]
- Ribani, M.; Bottoli, C.B.G.; Collins, C.H.; Jardim, I.C.S.F.; Melo, L.F.C. Validation for chromatographic and electrophoretic methods. Quim. Nova 2004, 27, 771–780. [Google Scholar] [CrossRef]
- Jiang, Z.; Huang, Q.; Jia, D.; Zhong, M.; Tao, J.; Liao, G.; Huang, C.; Xu, X. Characterization of Organic Acid Metabolism and Expression of Related Genes During Fruit Development of Actinidia eriantha ‘Ganmi 6. ’ Plants 2020, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Du, J.; Du, L.; Luo, Q.; Xiong, J. Anti-fatigue activity of purified anthocyanins prepared from purple passion fruit (P. edulis Sim) epicarp in mice. J. Funct. Foods 2020, 65, 103725. [Google Scholar] [CrossRef]
- Lalitha, S. Primer Premier 5. Biotech Softw. Internet Rep. 2000, 1, 270–272. [Google Scholar] [CrossRef]
- Munhoz, C.F.; Santos, A.A.; Arenhart, R.A.; Santini, L.; Monteiro-Vitorello, C.B.; Vieira, M.L.C. Analysis of plant gene expression during passion fruit—Xanthomonas axonopodis interaction implicates lipoxygenase 2 in host defence. Ann. Appl. Biol. 2015, 167, 135–155. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Fruiting Stage | Rutin (ng/g) | Luteolin (ng/g) | Quercetin (ng/g) | Cyanidin-3-O-Glucoside Chloride (ng/g) | Peonidin-3-O-Glucoside (ng/g) | Pelargonidin-3-O-Glucoside (ng/g) |
|---|---|---|---|---|---|---|---|
| Fruitlet | 906.99 ± 79.10 c | 15.62 ± 0.48 b | 5.55 ± 2.31 c | 1.48 ± 0.58 d | 1.13 ± 0.85 c | 3.53 ± 0.53 c | |
| Green | 1386.36 ± 132.25 c | 13.55 ± 0.88 b | 4.63 ± 4.50 c | 3.91 ± 0.60 d | 2.02 ± 0.19 c | 4.15 ± 0.39 c | |
| Purple | Veraison | 2384.89 ± 1309.13 c | 13.50 ± 3.05 b | 9.01 ± 5.30 c | 3881.42 ± 572.51 c | 411.03 ± 310.05 c | 75.94 ± 32.73 c |
| Maturation | 9499.92 ± 692.22 b | 13.91 ± 1.20 b | 21.14 ± 3.24 b | 5927.90 ± 303.74 b | 3351.67 ± 667.85 b | 209.71 ± 25.81 b | |
| Ripening | 22,569.60 ± 3386.66 a | 29.19 ± 4.52 a | 35.25 ± 2.31 a | 7341.62 ± 639.87 a | 9793.08 ± 3045.70 a | 511.92 ± 109.69 a | |
| Yellow | Fruitlet | 779.55 ± 74.56 ab | 15.30 ± 4.40 a | 3.08 ± 0.06 a | 0.70 ± 0.55 c | 0 | 2.54 ± 0.05 c |
| Green | 865.34 ± 36.12 abc | 10.88 ± 0.71 ab | 3.09 ± 0.06 a | 5.88 ± 0.51 a | 0 | 3.77 ± 0.48 b | |
| Veraison | 565.93 ± 54.38 d | 8.41 ± 0.62 c | 0.65 ± 0.08 b | 5.76 ± 1.67 a | 0 | 4.88 ± 0.89 a | |
| Maturation | 664.39 ± 34.58 bc | 8.51 ± 0.62 c | 0 | 3.66 ± 1.38 b | 0.06 ± 0.11 b | 3.98 ± 0.09 ab | |
| Ripening | 705.82 ± 168.55 a | 14.84 ± 4.06 a | 0 | 2.90 ± 0.79 b | 0.96 ± 0.38 a | 4.59 ± 0.34 ab |
| Flavonoid/Anthocyanin Component | Linearity (r2) | Slope (y) | Response (Sy) | Sy/y | LOD * (µg·mL−1) | LOQ ** (µg·mL−1) |
|---|---|---|---|---|---|---|
| Rutin | 0.999303 | 0.2737 | 5.2262 | 19.0921 | 63.00 | 190.92 |
| Luteolin | 0.999692 | 0.2745 | 4.9727 | 18.1111 | 59.76 | 181.11 |
| Quercetin | 0.999667 | 0.2756 | 4.6358 | 16.8164 | 55.49 | 168.16 |
| Cyanidin-3-O-glucoside chloride | 0.998590 | 0.2767 | 4.3319 | 15.6526 | 51.65 | 156.52 |
| Peonidin-3-O-glucoside | 0.999506 | 0.2757 | 4.6096 | 16.7147 | 55.15 | 167.14 |
| Pelargonidin-3-O-glucoside | 0.998351 | 0.2754 | 4.7720 | 17.3254 | 57.17 | 173.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, M.; Ali, M.M.; He, Y.; Ma, S.; Rizwan, H.M.; Yang, Q.; Li, B.; Lin, Z.; Chen, F. Flavonoids Accumulation in Fruit Peel and Expression Profiling of Related Genes in Purple (Passiflora edulis f. edulis) and Yellow (Passiflora edulis f. flavicarpa) Passion Fruits. Plants 2021, 10, 2240. https://doi.org/10.3390/plants10112240
Shi M, Ali MM, He Y, Ma S, Rizwan HM, Yang Q, Li B, Lin Z, Chen F. Flavonoids Accumulation in Fruit Peel and Expression Profiling of Related Genes in Purple (Passiflora edulis f. edulis) and Yellow (Passiflora edulis f. flavicarpa) Passion Fruits. Plants. 2021; 10(11):2240. https://doi.org/10.3390/plants10112240
Chicago/Turabian StyleShi, Meng, Muhammad Moaaz Ali, Yinying He, Songfeng Ma, Hafiz Muhammad Rizwan, Qiang Yang, Binqi Li, Zhimin Lin, and Faxing Chen. 2021. "Flavonoids Accumulation in Fruit Peel and Expression Profiling of Related Genes in Purple (Passiflora edulis f. edulis) and Yellow (Passiflora edulis f. flavicarpa) Passion Fruits" Plants 10, no. 11: 2240. https://doi.org/10.3390/plants10112240
APA StyleShi, M., Ali, M. M., He, Y., Ma, S., Rizwan, H. M., Yang, Q., Li, B., Lin, Z., & Chen, F. (2021). Flavonoids Accumulation in Fruit Peel and Expression Profiling of Related Genes in Purple (Passiflora edulis f. edulis) and Yellow (Passiflora edulis f. flavicarpa) Passion Fruits. Plants, 10(11), 2240. https://doi.org/10.3390/plants10112240