
Membrane Trafficking Proteins: A New Target to Identify Resistance to Viruses in Plants



Abstract
1. Introduction
2. Plant Proteins Involved in Backward Vesicle Mechanisms
3. Plant Proteins Involved in Inward Vesicle Mechanisms
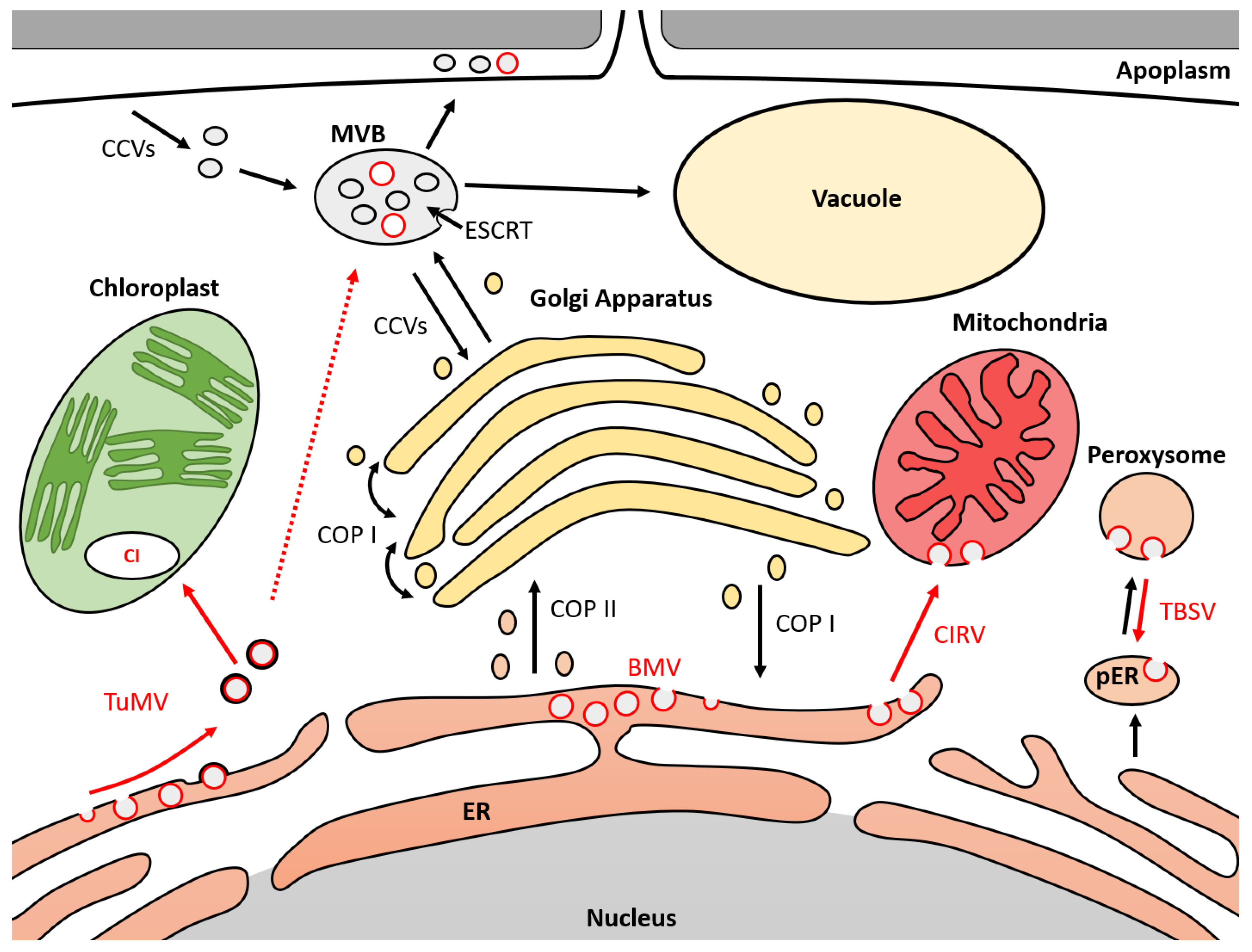
4. Host Intracellular Membrane Association with Viral Replication Complexes
5. Membrane Deformation and Involved Proteins during Replication of Tombusviruses
6. Membrane Deformation and Involved Proteins during Replication of Potyviruses
7. Membrane Deformation and Involved Proteins during Replication of Bromoviruses
8. Challenges and Opportunities for the Future
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dinar, A.; Zilberman, D. Impact of Pesticides on Farmer Health and the Rice Environment. J. Petrol. 1995, 369, 1689–1699. [Google Scholar] [CrossRef]
- Purdue, M.P.; Hoppin, J.A.; Blair, A.; Dosemeci, M.; Alavanja, M.C.R. Occupational Exposure to Organochlorine Insecticides and Cancer Incidence in the Agricultural Health Study. Int. J. Cancer 2007, 120, 642–649. [Google Scholar] [CrossRef]
- Hoisington, D.; Khairallah, M.; Reeves, T.; Ribaut, J.-M.; Skovmand, B.; Taba, S.; Warburton, M. Plant Genetic Resources: What Can They Contribute toward Increased Crop Productivity? Proc. Natl. Acad. Sci. USA 1998, 96, 5937–5943. [Google Scholar] [CrossRef]
- Desbiez, C.; Wipf-Scheibel, C.; Millot, P.; Berthier, K.; Girardot, G.; Gognalons, P.; Hirsch, J.; Moury, B.; Nozeran, K.; Piry, S.; et al. Distribution and Evolution of the Major Viruses Infecting Cucurbitaceous and Solanaceous Crops in the French Mediterranean Area. Virus Res. 2020, 286, 198042. [Google Scholar] [CrossRef] [PubMed]
- Revers, F.; Nicaise, V. Plant Resistance to Infection by Viruses. eLS 2014, 1–10. [Google Scholar] [CrossRef]
- Lecoq, H.; Moury, B.; Desbiez, C.; Palloix, A.; Pitrat, M. Durable Virus Resistance in Plants through Conventional Approaches: A Challenge. Virus Res. 2004, 100, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Fabre, F.; Rousseau, E.; Mailleret, L.; Moury, B. Durable Strategies to Deploy Plant Resistance in Agricultural Landscapes. New Phytol. 2012, 193, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Wang, A. Dissecting the Molecular Network of Virus-Plant Interactions: The Complex Roles of Host Factors. Annu. Rev. Phytopathol. 2015, 53, 45–66. [Google Scholar] [CrossRef]
- Hyodo, K.; Okuno, T. Pathogenesis Mediated by Proviral Host Factors Involved in Translation and Replication of Plant Positive-Strand RNA Viruses. Curr. Opin. Virol. 2016, 17, 11–18. [Google Scholar] [CrossRef]
- Bastet, A.; Zafirov, D.; Giovinazzo, N.; Guyon-Debast, A.; Nogué, F.; Robaglia, C.; Gallois, J.L. Mimicking Natural Polymorphism in EIF4E by CRISPR-Cas9 Base Editing Is Associated with Resistance to Potyviruses. Plant Biotechnol. J. 2019, 17, 1736–1750. [Google Scholar] [CrossRef]
- Piron, F.; Nicolaï, M.; Minoïa, S.; Piednoir, E.; Moretti, A.; Salgues, A.; Zamir, D.; Caranta, C.; Bendahmane, A. An Induced Mutation in Tomato EiF4E Leads to Immunity to Two Potyviruses. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Ouibrahim, L.; Mazier, M.; Estevan, J.; Pagny, G.; Decroocq, V.; Desbiez, C.; Moretti, A.; Gallois, J.L.; Caranta, C. Cloning of the Arabidopsis Rwm1 Gene for Resistance to Watermelon Mosaic Virus Points to a New Function for Natural Virus Resistance Genes. Plant J. 2014, 79, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Bednarek, S.Y.; Backues, S.K. Plant Dynamin-Related Protein Families DRP1 and DRP2 in Plant Development. Biochem. Soc. Trans. 2010, 38, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Cui, X.; Dai, Z.; He, R.; Li, Y.; Yu, K.; Bernards, M.; Chen, X.; Wang, A. A Plant RNA Virus Hijacks Endocytic Proteins to Establish Its Infection in Plants. Plant J. 2020, 101, 384–400. [Google Scholar] [CrossRef] [PubMed]
- Grangeon, R.; Agbeci, M.; Chen, J.; Grondin, G.; Zheng, H.; Laliberte, J.-F. Impact on the Endoplasmic Reticulum and Golgi Apparatus of Turnip Mosaic Virus Infection. J. Virol. 2012, 86, 9255–9265. [Google Scholar] [CrossRef] [PubMed]
- Sasvari, Z.; Kovalev, N.; Gonzalez, P.A.; Xu, K.; Nagy, P.D. Assembly-Hub Function of ER-Localized SNARE Proteins in Biogenesis of Tombusvirus Replication Compartment. PLoS Pathog. 2018, 14, e1007028. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fuchs, S.; Zhang, J.; Wellford, S.; Schuldiner, M.; Wang, X. An Unrecognized Function for COPII Components in Recruiting the Viral Replication Protein BMV 1a to the Perinuclear ER. J. Cell Sci. 2016, 129, 3597–3608. [Google Scholar] [CrossRef]
- Verchot, J. Wrapping Membranes around Plant Virus Infection. Curr. Opin. Virol. 2011, 1, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Feng, Z. Tombusviruses Orchestrate the Host Endomembrane System to Create Elaborate Membranous Replication Organelles. Curr. Opin. Virol. 2021, 48, 30–41. [Google Scholar] [CrossRef]
- Jensen, D.; Schekman, R. COPII-Mediated Vesicle Formation at a Glance. J. Cell Sci. 2011, 124, 1–4. [Google Scholar] [CrossRef]
- Duden, R. ER-to-Golgi Transport: COP I and COP II Function. Mol. Membr. Biol. 2003, 20, 197–207. [Google Scholar] [CrossRef]
- Matsuoka, K.; Orci, L.; Amherdt, M.; Bednarek, S.Y.; Hamamoto, S.; Schekman, R.; Yeung, T. COPII-Coated Vesicle Formation Reconstituted with Purified Coat Proteins and Chemically Defined Liposomes. Cell 1998, 93, 263–275. [Google Scholar] [CrossRef]
- Bor, L.T.; Wang, Y.; Yan, S.O.; Hong, W. COPII and Exit from the Endoplasmic Reticulum. Biochim. Biophys. Acta Mol. Cell Res. 2005, 1744, 293–303. [Google Scholar] [CrossRef]
- Lord, C.; Ferro-Novick, S.; Miller, E.A. The Highly Conserved COPII Coat Complex Sorts Cargo from the Endoplasmic Reticulum and Targets It to the Golgi. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef]
- Béthune, J.; Wieland, F.T. Assembly of COPI and COPII Vesicular Coat Proteins on Membranes. Annu. Rev. Biophys. 2018, 47, 1–21. [Google Scholar] [CrossRef]
- Barlowe, C. COPII: A Membrane Coat That Forms Endoplasmic Reticulum-Derived Vesicles. FEBS Lett. 1995, 369, 93–96. [Google Scholar] [CrossRef]
- Faso, C.; Chen, Y.-N.; Tamura, K.; Held, M.; Zemelis, S.; Marti, L.; Saravanan, R.; Hummel, E.; Kung, L.; Miller, E.; et al. A Missense Mutation in the Arabidopsis COPII Coat Protein Sec24A Induces the Formation of Clusters of the Endoplasmic Reticulum and Golgi Apparatus. Plant Cell 2009, 21, 3655–3671. [Google Scholar] [CrossRef] [PubMed]
- Hughson, F.M. Copy Coats: COPI Mimics Clathrin and COPII. Cell 2010, 142, 19–21. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Glick, B.S.; Nakano, A. Membrane Traffic within the Golgi Apparatus. Annu. Rev. Cell Dev. Biol. 2009, 25, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Seaman, M.N.J.; Michael McCaffery, J.; Emr, S.D. A Membrane Coat Complex Essential for Endosome-to-Golgi Retrograde Transport in Yeast. J. Cell Biol. 1998, 142, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Huotari, J.; Helenius, A. Endosome Maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef]
- Pearse, B.M.F. Clathrin: A Unique Protein Associated with Intracellular Transfer of Membrane by Coated Vesicles. Proc. Natl. Acad. Sci. USA 1976, 73, 1255–1259. [Google Scholar] [CrossRef]
- Roth, T.F.; Porter, K.R. Yolk Protein Uptake in the Oocyte of the Mosquito Aedes aegypti L. J. Cell Biol. 1964, 20, 313–332. [Google Scholar] [CrossRef]
- Paul, M.J.; Frigerio, L. Coated Vesicles in Plant Cells. Semin. Cell Dev. Biol. 2007, 18, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Drake, M.T.; Kornfeld, S. ADP-Ribosylation Factor 1 Dependent Clathrin-Coat Assembly on Synthetic Liposomes. Proc. Natl. Acad. Sci. USA 1999, 96, 5013–5018. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake, G.; LaMontagne, E.D.; Heese, A. Never Walk Alone: Clathrin-Coated Vesicle (CCV) Components in Plant Immunity. Annu. Rev. Phytopathol. 2019, 57, 387–409. [Google Scholar] [CrossRef] [PubMed]
- Kaksonen, M.; Roux, A. Mechanisms of Clathrin-Mediated Endocytosis. Nat. Rev. Mol. Cell Biol. 2018, 19, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, E.; Greene, L.E. Multiple Roles of Auxilin and Hsc70 in Clathrin-Mediated Endocytosis. Traffic 2007, 8, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, L.; Hong, W. SNARE Proteins in Membrane Trafficking. Traffic 2017, 18, 767–775. [Google Scholar] [CrossRef]
- Jahn, R.; Scheller, R.H. SNAREs—Engines for Membrane Fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef]
- Bonifacino, J.S.; Glick, B.S. Review: The Mechanisms of Vesicle Budding and Fusion. Cell 2004, 116, 153–166. [Google Scholar] [CrossRef]
- Takemoto, K.; Ebine, K.; Askani, J.C.; Krüger, F.; Gonzalez, Z.A.; Ito, E.; Goh, T.; Schumacher, K.; Nakano, A.; Ueda, T. Distinct Sets of Tethering Complexes, SNARE Complexes, and Rab GTPases Mediate Membrane Fusion at the Vacuole in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 201717839. [Google Scholar] [CrossRef]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and Secretion of Exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef]
- Sahu, R.; Kaushik, S.; Clement, C.C.; Cannizzo, E.S.; Scharf, B.; Follenzi, A.; Potolicchio, I.; Nieves, E.; Cuervo, A.M.; Santambrogio, L. Microautophagy of Cytosolic Proteins by Late Endosomes. Dev. Cell 2011, 20, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Zhuang, X.; Shen, J.; Jiang, L. Plant ESCRT Complexes: Moving Beyond Endosomal Sorting. Trends Plant Sci. 2017, 22, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Fader, C.M.; Colombo, M.I. Autophagy and Multivesicular Bodies: Two Closely Related Partners. Cell Death Differ. 2009, 16, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Christ, L.; Raiborg, C.; Wenzel, E.M.; Campsteijn, C.; Stenmark, H. Cellular Functions and Molecular Mechanisms of the ESCRT Membrane-Scission Machinery. Trends Biochem. Sci. 2017, 42, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Emr, S.D. THE ESCRT COMPLEXES: Structure and Mechanism of a Membrane-Trafficking Network. Annu. Rev. Biophys. Biomol. Struct. 2006, 35, 277–298. [Google Scholar] [CrossRef]
- Bowers, K.; Lottridge, J.; Helliwell, S.B.; Goldthwaite, L.M.; Luzio, J.P.; Stevens, T.H. Protein-Protein Interactions of ESCRT Complexes in the Yeast Saccharomyces Cerevisiae. Traffic 2004, 5, 194–210. [Google Scholar] [CrossRef] [PubMed]
- Reyes, F.C.; Buono, R.A.; Roschzttardtz, H.; Di Rubbo, S.; Yeun, L.H.; Russinova, E.; Otegui, M.S. A Novel Endosomal Sorting Complex Required for Transport (ESCRT) Component in Arabidopsis Thaliana Controls Cell Expansion and Development. J. Biol. Chem. 2014, 289, 4980–4988. [Google Scholar] [CrossRef]
- Spitzer, C.; Reyes, F.C.; Buono, R.; Sliwinski, M.K.; Haas, T.J.; Otegui, M.S. The ESCRT-Related CHMP1A and B Proteins Mediate Multivesicular Body Sorting of Auxin Carriers in Arabidopsis and Are Required for Plant Development. Plant Cell 2009, 21, 749–766. [Google Scholar] [CrossRef]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The ESCRT Pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef]
- Hurley, J.H. The ESCRT Complexes. Crit. Rev. Biochem. Mol. Biol. 2011, 45, 463–487. [Google Scholar] [CrossRef] [PubMed]
- Piper, R.C.; Katzmann, D.J. Biogenesis and Function of Multivesicular Bodies. Annu. Rev. Cell Dev. Biol. 2007, 23, 519–547. [Google Scholar] [CrossRef]
- Shahriari, M.; Richter, K.; Keshavaiah, C.; Sabovljevic, A.; Huelskamp, M.; Schellmann, S. The Arabidopsis ESCRT Protein-Protein Interaction Network. Plant Mol. Biol. 2011, 76, 85–96. [Google Scholar] [CrossRef]
- Scott, A.; Chung, H.-Y.; Gonciarz-Swiatek, M.; Hill, G.C.; Whitby, F.G.; Gaspar, J.; Holton, J.M.; Viswanathan, R.; Ghaffarian, S.; Hill, C.P.; et al. Structural and Mechanistic Studies of VPS4 Proteins. EMBO J. 2005, 24, 3658–3669. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Hanson, P.I. Membrane Budding and Scission by ESCRT Machinery. Nat. Rev. Mol. Cell Biol. 2010, 11, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Ettema, T.J.G.; Bernander, R. Cell Division and the ESCRT Complex: A Surprise from the Archaea. Commun. Integr. Biol. 2009, 2, 86–88. [Google Scholar] [CrossRef]
- Luhtala, N.; Odorizzi, G. Bro1 Coordinates Deubiquitination in the Multivesicular Body Pathway by Recruiting Doa4 to Endosomes. J. Cell Biol. 2004, 166, 717–729. [Google Scholar] [CrossRef]
- Boualem, A.; Dogimont, C.; Bendahmane, A. The Battle for Survival between Viruses and Their Host Plants. Curr. Opin. Virol. 2016, 17, 32–38. [Google Scholar] [CrossRef]
- Elena, S.F.; Rodrigo, G. Towards an Integrated Molecular Model of Plant-Virus Interactions. Curr. Opin. Virol. 2012, 2, 713–718. [Google Scholar] [CrossRef]
- Kovalev, N.; Inaba, J.I.; Li, Z.; Nagy, P.D. The Role of Co-Opted ESCRT Proteins and Lipid Factors in Protection of Tombusviral Double-Stranded RNA Replication Intermediate against Reconstituted RNAi in Yeast. PLoS Pathog. 2017, 13, 1–26. [Google Scholar] [CrossRef]
- Nagy, P.D.; Strating, J.R.P.M.; van Kuppeveld, F.J.M. Building Viral Replication Organelles: Close Encounters of the Membrane Types. PLoS Pathog. 2016, 12, 6–11. [Google Scholar] [CrossRef]
- Paul, D.; Bartenschlager, R. Architecture and Biogenesis of Plus-Strand RNA Virus Replication Factories. World J. Virol. 2013, 2, 32–48. [Google Scholar] [CrossRef]
- Jiang, J.; Laliberte, J.-F. Membrane Association for Plant Virus Replication and Movement. Curr. Res. Top. Plant Virol. 2016, 1–335. [Google Scholar] [CrossRef]
- Weber-Lofti, F.; Dietrich, A.; Russo, M.; Rubino, L. Mitochondrial Targeting and Membrane Anchoring of a Viral Replicase in Plant and Yeast Cells. J. Virol. 2002, 76, 10485–10496. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, Y.; Komoda, K.; Yamanaka, T.; Tamai, A.; Meshi, T.; Funada, R.; Tsuchiya, T.; Naito, S. Subcellular Localization of Host and Viral Proteins Associated with Tobamovirus RNA Replication. EMBO J. 2003, 22, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Laliberté, J.-F.; Zheng, H. Viral Manipulation of Plant Host Membranes. Annu. Rev. Virol. 2014, 1, 237–259. [Google Scholar] [CrossRef] [PubMed]
- Laliberté, J.-F.; Sanfaçon, H. Cellular Remodeling During Plant Virus Infection. Annu. Rev. Phytopathol. 2010, 48, 69–91. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Pogany, J.; Lin, J.Y. How Yeast Can Be Used as a Genetic Platform to Explore Virus-Host Interactions: From “omics” to Functional Studies. Trends Microbiol. 2014, 22, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Panavas, T.; Serviene, E.; Brasher, J.; Nagy, P.D. Yeast Genome-Wide Screen Reveals Dissimilar Sets of Host Genes Affecting Replication of RNA Viruses. Proc. Natl. Acad. Sci. USA 2005, 102, 7326–7331. [Google Scholar] [CrossRef]
- Diaz, A.; Zhang, J.; Ollwerther, A.; Wang, X.; Ahlquist, P. Host ESCRT Proteins Are Required for Bromovirus RNA Replication Compartment Assembly and Function. PLoS Pathog. 2015, 11, e1004742. [Google Scholar] [CrossRef] [PubMed]
- Rapaport, D. How Does the TOM Complex Mediate Insertion of Precursor Proteins into the Mitochondrial Outer Membrane? J. Cell Biol. 2005, 171, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Cross, L.L.; Ebeed, H.T.; Baker, A. Peroxisome Biogenesis, Protein Targeting Mechanisms and PEX Gene Functions in Plants. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Barajas, D.; Jiang, Y.; Nagy, P.D. A Unique Role for the Host ESCRT Proteins in Replication of Tomato Bushy Stunt Virus. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.G.L.; Clendening, E.A.; Sheen, H.; Gidda, S.K.; White, K.A.; Mullen, R.T. A Unique N-Terminal Sequence in the Carnation Italian Ringspot Virus P36 Replicase-Associated Protein Interacts with the Host Cell ESCRT-I Component Vps23. J. Virol. 2014, 88, 6329–6344. [Google Scholar] [CrossRef] [PubMed]
- Barajas, D.; Martín, I.F.; Pogany, J.; Risco, C.; Nagy, P.D. Noncanonical Role for the Host Vps4 AAA+ ATPase ESCRT Protein in the Formation of Tomato Bushy Stunt Virus Replicase. PLoS Pathog. 2014, 10, 24–32. [Google Scholar] [CrossRef]
- Barajas, D.; Nagy, P.D. Ubiquitination of Tombusvirus P33 Replication Protein Plays a Role in Virus Replication and Binding to the Host Vps23p ESCRT Protein. Virology 2010, 397, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Xu, K.; Kovalev, N.; Nagy, P.D. Recruitment of Vps34 PI3K and Enrichment of PI3P Phosphoinositide in the Viral Replication Compartment Is Crucial for Replication of a Positive-Strand RNA Virus. PLoS Pathog. 2019, 15, 1–30. [Google Scholar] [CrossRef]
- Hyodo, K.; Mine, A.; Taniguchi, T.; Kaido, M.; Mise, K.; Taniguchi, H.; Okuno, T. ADP Ribosylation Factor 1 Plays an Essential Role in the Replication of a Plant RNA Virus. J. Virol. 2013, 87, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, D.G.; Jiang, J.; Movahed, N.; Germain, H.; Yamaji, Y.; Zheng, H.; Laliberté, J.-F.F. Turnip Mosaic Virus Uses the Snare Protein Vti11 in an Unconventional Route for Replication Vesicle Trafficking. Plant Cell 2018, 30, 2594–2615. [Google Scholar] [CrossRef]
- Wei, T.; Zhang, C.; Hou, X.; Sanfaçon, H.; Wang, A. The SNARE Protein Syp71 Is Essential for Turnip Mosaic Virus Infection by Mediating Fusion of Virus-Induced Vesicles with Chloroplasts. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef]
- Jiang, J.; Patarroyo, C.; Garcia Cabanillas, D.; Zheng, H.; Laliberté, J.-F. The Vesicle-Forming 6K 2 Protein of Turnip Mosaic Virus Interacts with the COPII Coatomer Sec24a for Viral Systemic Infection. J. Virol. 2015, 89, 6695–6710. [Google Scholar] [CrossRef]
- Wei, T.; Wang, A. Biogenesis of Cytoplasmic Membranous Vesicles for Plant Potyvirus Replication Occurs at Endoplasmic Reticulum Exit Sites in a COPI- and COPII-Dependent Manner. J. Virol. 2008, 82, 12252–12264. [Google Scholar] [CrossRef]
- Amano, M.; Mochizuki, A.; Kawagoe, Y.; Iwahori, K.; Niwa, K.; Svoboda, J.; Maeda, T.; Imura, Y. High-Resolution Mapping of Zym, a Recessive Gene for Zucchini Yellow Mosaic Virus Resistance in Cucumber. Theor. Appl. Genet. 2013, 126, 2983–2993. [Google Scholar] [CrossRef]
- Diaz, A.; Wang, X.; Ahlquist, P. Membrane-Shaping Host Reticulon Proteins Play Crucial Roles in Viral RNA Replication Compartment Formation and Function. Proc. Natl. Acad. Sci. USA 2010, 107, 16291–16296. [Google Scholar] [CrossRef]
- Giner, A.; Pascual, L.; Bourgeois, M.; Gyetvai, G.; Rios, P.; Picó, B.; Troadec, C.; Bendahmane, A.; Garcia-Mas, J.; Martín-Hernández, A.M. A Mutation in the Melon Vacuolar Protein Sorting 41 Prevents Systemic Infection of Cucumber Mosaic Virus. Sci. Rep. 2017, 7, 10471. [Google Scholar] [CrossRef]
- Navarro, B.; Russo, M.; Pantaleo, V.; Rubino, L. Cytological Analysis of Saccharomyces Cerevisiae Cells Supporting Cymbidium Ringspot Virus Defective Interfering RNA Replication. J. Gen. Virol. 2006, 87, 705–714. [Google Scholar] [CrossRef]
- Panavas, T.; Hawkins, C.M.; Panaviene, Z.; Nagy, P.D. The Role of the P33:P33/P92 Interaction Domain in RNA Replication and Intracellular Localization of P33 and P92 Proteins of Cucumber Necrosis Tombusvirus. Virology 2005, 338, 81–95. [Google Scholar] [CrossRef] [PubMed]
- McCartney, A.W.; Greenwood, J.S.; Fabian, M.R.; White, K.A.; Mullen, R.T. Localization of the Tomato Bushy Stunt Virus Replication Protein P33 Reveals a Peroxisome-to-Endoplasmic Reticulum Sorting Pathway. Plant Cell 2005, 17, 3513–3531. [Google Scholar] [CrossRef] [PubMed]
- Titorenko, V.I.; Rachubinski, R.A. The Peroxisome: Orchestrating Important Developmental Decisions from inside the Cell. J. Cell Biol. 2004, 164, 641–645. [Google Scholar] [CrossRef] [PubMed]
- Gunawardene, C.D.; Donaldson, L.W.; White, K.A. Tombusvirus Polymerase: Structure and Function. Virus Res. 2017, 234, 74–86. [Google Scholar] [CrossRef]
- Fernández de Castro, I.; Fernández, J.J.; Barajas, D.; Nagy, P.D.; Risco, C. Three-Dimensional Imaging of the Intracellular Assembly of a Functional Viral RNA Replicase Complex. J. Cell Sci. 2017, 130, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Jonczyk, M.; Pathak, K.B.; Sharma, M.; Nagy, P.D. Exploiting Alternative Subcellular Location for Replication: Tombusvirus Replication Switches to the Endoplasmic Reticulum in the Absence of Peroxisomes. Virology 2007, 362, 320–330. [Google Scholar] [CrossRef]
- Turner, K.A.; Sit, T.L.; Callaway, A.S.; Allen, N.S.; Lommel, S.A. Red Clover Necrotic Mosaic Virus Replication Proteins Accumulate at the Endoplasmic Reticulum. Virology 2004, 320, 276–290. [Google Scholar] [CrossRef]
- Sanfaçon, H. Replication of Positive-Strand RNA Viruses in Plants: Contact Points between Plant and Virus Components. Can. J. Bot. 2005, 83, 1529–1549. [Google Scholar] [CrossRef]
- Hwang, Y.T.; McCartney, A.W.; Gidda, S.K.; Mullen, R.T. Localization of the Carnation Italian Ringspot Virus Replication Protein P36 to the Mitochondrial Outer Membrane Is Mediated by an Internal Targeting Signal and the TOM Complex. BMC Cell Biol. 2008, 9, 1–26. [Google Scholar] [CrossRef]
- Oana, D.; Ziegler, A.; Torrance, L.; Gasemi, S.; Danci, M. Potyviridae Family—Short Review. J. Hortic. For. Biotechnol. 2009, 13, 410–421. [Google Scholar]
- Wei, T.; Huang, T.-S.; McNeil, J.; Laliberte, J.-F.; Hong, J.; Nelson, R.S.; Wang, A. Sequential Recruitment of the Endoplasmic Reticulum and Chloroplasts for Plant Potyvirus Replication. J. Virol. 2010, 84, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Schaad, M.C.; Jensen, P.E.; Carrington, J.C. Formation of Plant RNA Virus Replication Complexes on Membranes: Role of an Endoplasmic Reticulum-Targeted Viral Protein. EMBO J. 1997, 16, 4049–4059. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, C.; Laliberte, J.-F. The Poly(A) Binding Protein Is Internalized in Virus-Induced Vesicles or Redistributed to the Nucleolus during Turnip Mosaic Virus Infection. J. Virol. 2007, 81, 10905–10913. [Google Scholar] [CrossRef] [PubMed]
- Cotton, S.; Grangeon, R.; Thivierge, K.; Mathieu, I.; Ide, C.; Wei, T.; Wang, A.; Laliberte, J.-F. Turnip Mosaic Virus RNA Replication Complex Vesicles Are Mobile, Align with Microfilaments, and Are Each Derived from a Single Viral Genome. J. Virol. 2009, 83, 10460–10471. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Jiang, Z.; Zhang, K.; Wang, P.; Cao, X.; Yue, N.; Wang, X.; Zhang, X.; Li, Y.; Li, D.; et al. Three-Dimensional Analysis of Chloroplast Structures Associated with Virus Infection. Plant Physiol. 2018, 176, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, E.; Roingeard, P. Virus-Induced Double-Membrane Vesicles. Cell. Microbiol. 2015, 17, 45–50. [Google Scholar] [CrossRef]
- Cui, Y.; Gao, J.; He, Y.; Jiang, L. Plant Extracellular Vesicles. Protoplasma 2020, 257, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Movahed, N.; Cabanillas, D.G.; Wan, J.; Vali, H.; Laliberté, J.F.; Zheng, H. Turnip Mosaic Virus Components Are Released into the Extracellular Space by Vesicles in Infected Leaves. Plant Physiol. 2019, 180, 1375–1388. [Google Scholar] [CrossRef]
- Wan, J.; Basu, K.; Mui, J.; Vali, H.; Zheng, H.; Laliberté, J.-F. Ultrastructural Characterization of Turnip Mosaic Virus-Induced Cellular Rearrangements Reveals Membrane-Bound Viral Particles Accumulating in Vacuoles. J. Virol. 2015, 89, 12441–12456. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Cervera, M.T.; Garcia, J.A. Processing of the Plum Pox Virus Polyprotein at the P3-6K1junction Is Not Required for Virus Viability. J. Gen. Virol. 1995, 76, 951–956. [Google Scholar] [CrossRef]
- Chung, B.Y.W.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An Overlapping Essential Gene in the Potyviridae. Proc. Natl. Acad. Sci. USA 2008, 105, 5897–5902. [Google Scholar] [CrossRef]
- Jiang, J. Signal-Directed Endoplasmic Reticulum and Golgi Exit of Turnip Mosaic Virus 6K 2 Protein for Replication Vesicle Cellular Biogenesis. Ph.D. Thesis, Université du Québec, Gatineau, QC, Canada, 2015. [Google Scholar]
- Wu, G.; Cui, X.; Chen, H.; Renaud, J.B.; Yu, K.; Chen, X.; Wang, A. Dynamin-Like Proteins of Endocytosis in Plants Are Coopted by Potyviruses To Enhance Virus Infection. J. Virol. 2018, 92, 1–22. [Google Scholar] [CrossRef]
- Bamunusinghe, D.; Seo, J.-K.; Rao, A.L.N. Subcellular Localization and Rearrangement of Endoplasmic Reticulum by Brome Mosaic Virus Capsid Protein. J. Virol. 2011, 85, 2953–2963. [Google Scholar] [CrossRef]
- He, G.; Zhang, Z.; Sathanatham, P.; Diaz, A.; Wang, X. Brome Mosaic Virus (Bromoviridae). Encycl. Virol. 2020, 3, 252–259. [Google Scholar] [CrossRef]
- den Boon, J.A.; Chen, J.; Ahlquist, P. Identification of Sequences in Brome Mosaic Virus Replicase Protein 1a That Mediate Association with Endoplasmic Reticulum Membranes. J. Virol. 2001, 75, 12370–12381. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Westler, W.M.; Den Boon, J.A.; Wang, X.; Diaz, A.; Steinberg, H.A.; Ahlquist, P. An Amphipathic α-Helix Controls Multiple Roles of Brome Mosaic Virus Protein 1a in RNA Replication Complex Assembly and Function. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.; Chen, J.; Janda, M.; Sullivan, M.; Den Boon, J.; Ahlquist, P. A Positive-Strand RNA Virus Replication Complex Parallels Form and Function of Retrovirus Capsids. Mol. Cell 2002, 9, 505–514. [Google Scholar] [CrossRef]
- Voeltz, G.K.; Prinz, W.A.; Shibata, Y.; Rist, J.M.; Rapoport, T.A. A Class of Membrane Proteins Shaping the Tubular Endoplasmic Reticulum. Cell 2006, 124, 573–586. [Google Scholar] [CrossRef]
- Diaz, A.; Ahlquist, P. Role of Host Reticulon Proteins in Rearranging Membranes for Positive-Strand RNA Virus Replication. Curr. Opin. Microbiol. 2012, 15, 519–524. [Google Scholar] [CrossRef]
- Radisky, D.C.; Snyder, W.B.; Emr, S.D.; Kaplan, J. Characterization of VPS41, a Gene Required for Vacuolar Trafficking and High-Affinity Iron Transport in Yeast. Cell Biol. 1997, 94, 5662–5666. [Google Scholar] [CrossRef]
- Winter, V.; Hauser, M.T. Exploring the ESCRTing Machinery in Eukaryotes. Trends Plant Sci. 2006, 11, 115–123. [Google Scholar] [CrossRef]
- Restrepo-Hartwig, M.; Ahlquist, P. Brome Mosaic Virus RNA Replication Proteins 1a and 2a Colocalize and 1a Independently Localizes on the Yeast Endoplasmic Reticulum. J. Virol. 1999, 73, 10303–10309. [Google Scholar] [CrossRef]
- Lõhmus, A.; Varjosalo, M.; Mäkinen, K. Protein Composition of 6K2-Induced Membrane Structures Formed during Potato Virus A Infection. Mol. Plant Pathol. 2016, 17, 943–958. [Google Scholar] [CrossRef] [PubMed]
- Thaminy, S.; Miller, J.; Stagljar, I. The Split-Ubiquitin Membrane-Based Yeast Two-Hybrid System. Protein-Protein Interact. 2005, 261, 297–312. [Google Scholar] [CrossRef]
- Kurowska, M.; Daszkowska-Golec, A.; Gruszka, D.; Marzec, M.; Szurman, M.; Szarejko, I.; Maluszynski, M. TILLING—A Shortcut in Functional Genomics. J. Appl. Genet. 2011, 52, 371–390. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Virus | Viral Protein | Host | Host Protein | Mechanisms | Relation between Pathogen and Host | Ref. |
|---|---|---|---|---|---|---|
| Tombusvirus | ||||||
| TBSV | p33 | A. thaliana | VPS23, BRO1 | ESCRT | Protein interaction | [75] |
| TBSV | p33 | Yeast | VPS4, VPS24 | ESCRT | Protein interaction | [77] |
| TBSV | p33 | Yeast | VPS23 | ESCRT | Protein interaction | [78] |
| TBSV | p33 | Yeast | UFE1, USE1 | SNARE | Protein interaction | [16] |
| TBSV | p33 | Yeast | VPS34 | ESCRT | Protein interaction | [79] |
| TBSV | - | Yeast | VPS15, VPS30, VPS34 | ESCRT | KO reducing viral replication | [79] |
| TBSV | - | Yeast | VPS18, VPS32, VPS24, VPS29, VPS4, VPS41, DID2, VPS23, VPS28, VPS51, VPS61, VPS69 | ESCRT, SNARE | KO reducing viral replication | [77] |
| TBSV, CIRV | - | A. thaliana | VPS4 | ESCRT | Dominant-negative reducing viral replication | [76] |
| CIRV | p36 | Yeast | VPS23 | ESCRT | Protein interaction | [76] |
| TBSV | p33 | Yeast | PEX19 | Protein interaction | [74] | |
| RCNMV | p27 | In vitro | ARF1 | COPI | Protein interaction | [80] |
| Potyvirus | Protein interaction | |||||
| TuMV | 6K2 | In vitro | VTI11 | SNARE | Protein interaction | [81] |
| TuMV | 6K2 | Yeast | VAP27 | SNARE | Protein interaction | [82] |
| TuMV | 6K2 | Yeast | SEC24a | COPII | Protein interaction | [83] |
| TuMV | 6K2, VPg, CP, CI | A. thaliana | DRP1/2 | CCV | Protein interaction | [77,84] |
| ZYMV | - | Cucumis sativus | VPS4 | ESCRT | Substitution inducing resistance | [85] |
| Bromovirus and Cucumovirus | ||||||
| BMV | 1a | In vitro | SNF7 | ESCRT | Protein interaction | [72] |
| BMV | 1a | In vitro | RTN1p, RTN2p, YOP1p | RHP | KO reducing viral replication | [86] |
| BMV | 1a | Yeast | VPS23, VPS20, SNF7, VPS24, VPS2, VPS4, DID2, VPS60 | ESCRT | KO reducing viral replication | [72] |
| CMV | Cucumis melo | VPS41 | SNARE | Substitution inducing resistance | [87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agaoua, A.; Bendahmane, A.; Moquet, F.; Dogimont, C. Membrane Trafficking Proteins: A New Target to Identify Resistance to Viruses in Plants. Plants 2021, 10, 2139. https://doi.org/10.3390/plants10102139
Agaoua A, Bendahmane A, Moquet F, Dogimont C. Membrane Trafficking Proteins: A New Target to Identify Resistance to Viruses in Plants. Plants. 2021; 10(10):2139. https://doi.org/10.3390/plants10102139
Chicago/Turabian StyleAgaoua, Aimeric, Abdelhafid Bendahmane, Frédéric Moquet, and Catherine Dogimont. 2021. "Membrane Trafficking Proteins: A New Target to Identify Resistance to Viruses in Plants" Plants 10, no. 10: 2139. https://doi.org/10.3390/plants10102139
APA StyleAgaoua, A., Bendahmane, A., Moquet, F., & Dogimont, C. (2021). Membrane Trafficking Proteins: A New Target to Identify Resistance to Viruses in Plants. Plants, 10(10), 2139. https://doi.org/10.3390/plants10102139