
Cannabis sativa: From Therapeutic Uses to Micropropagation and Beyond


Abstract
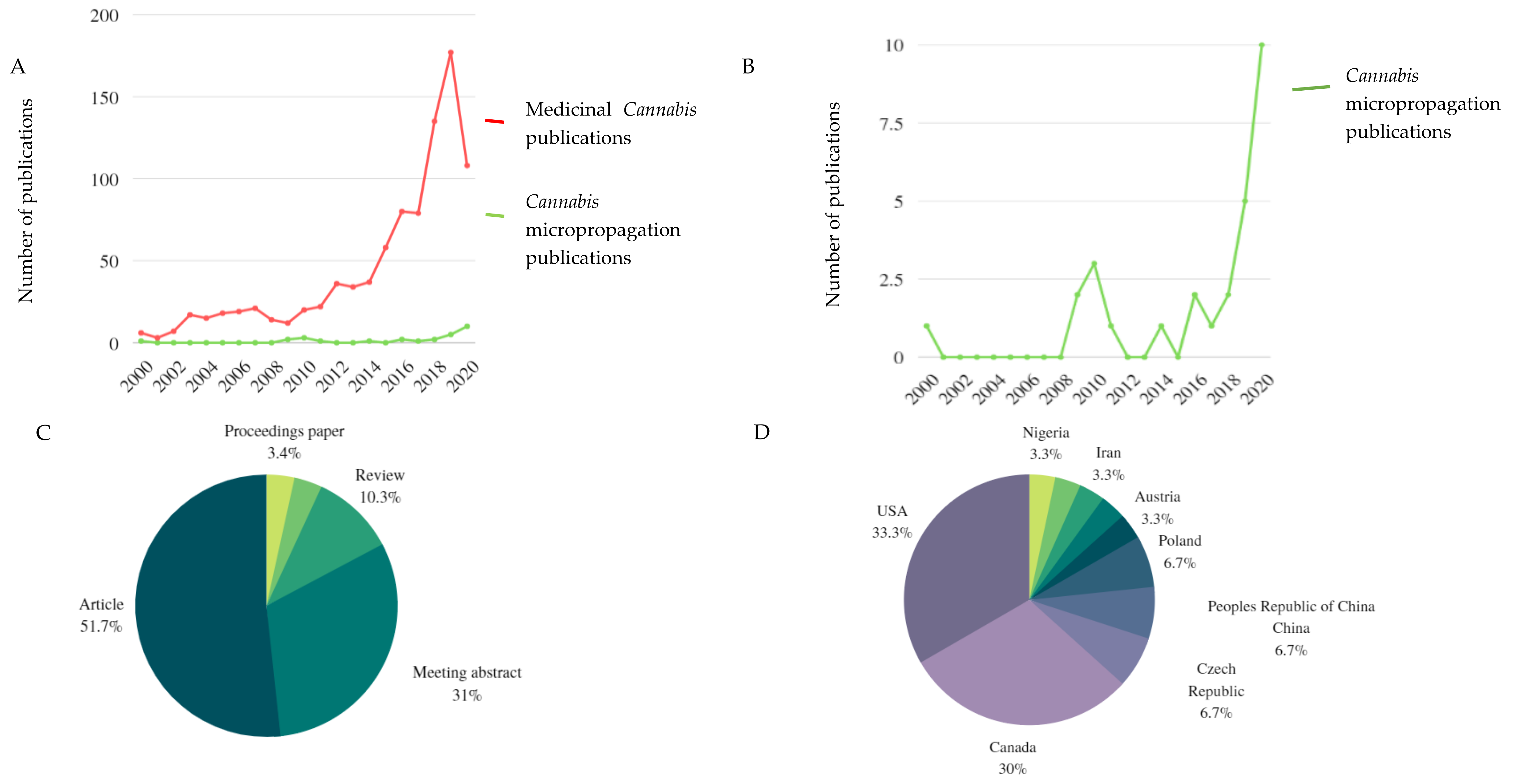
:1. Introduction
2. General Morphology of Cannabis sativa
2.1. Plant Morphology (C. sativa Type and C. indica Type)
2.2. Plant Parts Producing Cannabinoids (C. sativa Type and C. indica Type)
2.3. Chemical and Biosynthesis of Cannabis sativa Constituents
2.4. Therapeutic Uses of Cannabis sativa
3. Methodology
4. Legalisation and Propagation Strategies of Cannabis sativa
4.1. Conventional Propagation
4.2. Micropropagation of Cannabis sativa
4.2.1. Direct Organogenesis of Cannabis sativa
4.2.2. Indirect Organogenesis of Cannabis sativa
4.2.3. In Vitro and Ex Vitro Rooting of Cannabis sativa
4.2.4. Commercial Micropropagation of Cannabis sativa
4.3. In vitro Germplasm and Conservation
4.3.1. Cryopreservation
4.3.2. Synthetic Seed Technology
4.4. Genetic Engineering and Gene Editing
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Booth, J.K.; Bohlmann, J. Terpenes in Cannabis sativa—From plant genome to humans. Plant Sci. 2019, 284, 67–72. [Google Scholar] [CrossRef]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis sativa: The plant of the thousand and one molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Kisková, T.; Mungenast, F.; Suváková, M.; Jäger, W.; Thalhammer, T. Future aspects for cannabinoids in breast cancer therapy. Int. J. Mol. Sci. 2019, 20, 1673. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-L. An archaeological and historical account of Cannabis in China. Econ. Bot. 1974, 28, 437–448. [Google Scholar] [CrossRef]
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef]
- Jäger, A.K.; Hutchings, A.; van Staden, J. Screening of Zulu medicinal plants for prostaglandin-synthesis inhibitors. J. Ethnopharmacol. 1996, 52, 95–100. [Google Scholar] [CrossRef]
- Kalant, O.J. Report of the Indian hemp drugs commission, 1893–1894: A Critical Review. Int. J. Addict. 1972, 7, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.; Rueda, S.; Room, R.; Tyndall, M.; Fischer, B. Routes of administration for Cannabis use—Basic prevalence and related health outcomes: A scoping review and synthesis. Int. J. Drug Policy 2018, 52, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Kalant, H. Medicinal use of Cannabis: History and current status. Pain Res. Manag. 2001, 6, 80–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ElSohly, M.A.; Slade, D. Chemical constituents of marijuana: The complex mixture of natural cannabinoids. Life Sci. 2005, 78, 539–548. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Ross, S.A.; Slade, D.; Radwan, M.; Khan, I.A.; ElSohly, M.A. Minor oxygenated cannabinoids from high potency Cannabis sativa L. Phytochemistry 2015, 117, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Ligresti, A.; Petrosino, S.; Di Marzo, V. From Endocannabinoid profiling to “endocannabinoid therapeutics”. Curr. Opin. Chem. Biol. 2009, 13, 321–331. [Google Scholar] [CrossRef]
- Romero, P.; Peris, A.; Vergara, K.; Matus, J. Comprehending and improving Cannabis specialized metabolism in the systems biology era. Plant Sci. 2020, 298, 110571. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Lata, H.; Khan, I.A.; ElSohly, M.A. Cannabis sativa L.—Botany and Biotechnology; Springer: Cham, Switzerland, 2017; pp. 79–100. [Google Scholar]
- Thomas, B.F.; ElSohly, M.A. The botany of Cannabis sativa L. In The Analytical Chemistry of Cannabis; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–26. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Mehmedic, Z.; Khan, I.A.; Elsohly, M.A. Assessment of cannabinoids content in micropropagated plants of Cannabis sativa and their comparison with conventionally propagated plants and mother plant during developmental stages of growth. Planta Med. 2010, 76, 743–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Sanchez, I.J.; Verpoorte, R. Secondary metabolism in Cannabis. Phytochem. Rev. 2008, 7, 615–639. [Google Scholar] [CrossRef]
- Sirikantaramas, S.; Taura, F.; Morimoto, S.; Shoyama, Y. Recent Advances in Cannabis sativa Research: Biosynthetic studies and its potential in biotechnology. Curr. Pharm. Biotechnol. 2007, 8, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, N.; Gorelick, J.; Koch, S. Interplay between chemistry and morphology in medical Cannabis (Cannabis sativa L.). Ind. Crop. Prod. 2018, 129, 185–194. [Google Scholar] [CrossRef]
- Khakimov, B.; Bak, S.; Engelsen, S.B. High-throughput cereal metabolomics: Current analytical technologies, challenges, and perspectives. J. Cereal Sci. 2014, 59, 393–418. [Google Scholar] [CrossRef]
- Booth, J.K.; Page, J.E.; Bohlmann, J. Terpene synthases from Cannabis sativa. PLoS ONE 2017, 12, e0173911. [Google Scholar] [CrossRef] [Green Version]
- Barrales-cureño, H.J. Chemical characteristics, therapeutic uses, and legal aspects of the cannabinoids of Cannabis sativa: A Review. Braz. Arch. Biol. Technol. 2020, 63, 1–14. [Google Scholar] [CrossRef]
- Guzmán, M. Cannabinoids: Potential anticancer agents. Nat. Rev. Cancer 2003, 3, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Sarfaraz, S.; Adhami, V.M.; Syed, D.N.; Afaq, F.; Mukhtar, H. Cannabinoids for cancer treatment: Progress and promise. Cancer Res. 2008, 68, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Chakravarti, B.; Ravi, J.; Ganju, R.K. Cannabinoids as therapeutic agents in cancer: Status and future implications. Oncotarget 2014, 5, 5852–5872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertwee, R.G. Endocannabinoids; Springer: Cham, Switzerland, 2015; pp. 1–472. [Google Scholar] [CrossRef]
- Bifulco, M.; Laezza, C.; Portella, G.; Vitale, M.; Orlando, P.; De Petrocellis, L.; Di Marzo, V. Control by the endogenous cannabinoid system of ras oncogene-dependent tumor growth. FASEB J. 2001, 15, 2745–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munson, A.E.; Harris, L.S.; Friedman, M.A.; Dewey, W.L.; Carchman, R.A. Antineoplastic activity of cannabinoids. J. Natl. Cancer Inst. 1975, 55, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; Galve-Roperh, I.; Canova, C.; Brachet, P.; Guzmán, M. Δ9-Tetrahydrocannabinol induces apoptosis in C6 glioma cells. FEBS Lett. 1998, 436, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Jacobsson, S.O.; Rongård, E.; Stridh, M.; Tiger, G.; Fowler, C.J. Serum-dependent effects of tamoxifen and cannabinoids upon C6 glioma cell viability. Biochem. Pharmacol. 2000, 60, 1807–1813. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Melck, D.; Palmisano, A.; Bisogno, T.; Laezza, C.; Bifulco, M.; Di Marzo, V. The endogenous cannabinoid anandamide inhibits human breast cancer cell proliferation. Proc. Natl. Acad. Sci. USA 1998, 95, 8375–8380. [Google Scholar] [CrossRef] [Green Version]
- Ligresti, A.; Bisogno, T.; Matias, I.; De Petrocellis, L.; Cascio, M.G.; Cosenza, V.; D’Argenio, G.; Scaglione, G.; Bifulco, M.; Sorrentini, I.; et al. Possible endocannabinoid control of colorectal cancer growth. Gastroenterology 2003, 125, 677–687. [Google Scholar] [CrossRef]
- Mimeault, M.; Pommery, N.; Wattez, N.; Bailly, C.; Hénichart, J.-P. Anti-proliferative and apoptotic effects of anandamide in human prostatic cancer cell lines: Implication of epidermal growth factor receptor down-regulation and ceramide production. Prostate 2003, 56, 1–12. [Google Scholar] [CrossRef]
- Mechoulam, R.; Hanuš, L. A historical overview of chemical research on cannabinoids. Chem. Phys. Lipids 2000, 108, 1–13. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; ElSohly, M.A. Propagation of Cannabis for clinical research: An approach towards a modern herbal medicinal products development. Front. Plant Sci. 2020, 11, 958. [Google Scholar] [CrossRef]
- Naville, S. 340 Billion: The Global Cannabis Market. Available online: https://www.gbnews.ch/340-billion-the-global-cannabis-market/ (accessed on 15 December 2020).
- Nunley, K. New Analysis Values Global Cannabis Market at $344 Billion. Available online: https://www.medicalmarijuanainc.com/news/new-analysis-values-global-cannabis-market-at-344-billion/ (accessed on 15 December 2020).
- Backer, R.; Schwinghamer, T.; Rosenbaum, P.; McCarty, V.; Bilodeau, S.E.; Lyu, D.; Ahmed, B.; Robinson, G.; Lefsrud, M.; Wilkins, O.; et al. Closing the yield gap for Cannabis: A Meta-analysis of factors determining Cannabis yield. Front. Plant Sci. 2019, 10, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, K.; Struik, P.C.; Yin, X.; Calzolari, D.; Musio, S.; Thouminot, C.; Bjelková, M.; Stramkale, V.; Magagnini, G.; Amaducci, S. A comprehensive study of planting density and nitrogen fertilization effect on dual-purpose hemp (Cannabis sativa L.) cultivation. Ind. Crops. Prod. 2017, 107, 427–438. [Google Scholar] [CrossRef]
- García-Tejero, I.; Zuazo, V.H.D.; Sánchez-Carnenero, C.; Hernández, A.; Ferreiro-Vera, C.; Casano, S. Seeking suitable agronomical practices for industrial hemp (Cannabis sativa L.) cultivation for biomedical applications. Ind. Crop. Prod. 2019, 139, 111524. [Google Scholar] [CrossRef]
- Baldini, M.; Ferfuia, C.; Zuliani, F.; Danuso, F. Suitability assessment of different hemp (Cannabis sativa L.) varieties to the cultivation environment. Ind. Crop. Prod. 2019, 143, 111860. [Google Scholar] [CrossRef]
- Jin, D.; Jin, S.; Chen, J. Cannabis indoor growing conditions, management practices, and post-harvest treatment: A review. Am. J. Plant Sci. 2019, 10, 925–946. [Google Scholar] [CrossRef] [Green Version]
- Pepe, M.; Hesami, M.; Small, F.; Maxwell, A.; Jones, P.; Maxwell, A.; Jones, P. Comparative analysis of machine learning and evolutionary optimization algorithms for precision tissue culture of Cannabis sativa: Prediction and validation of in vitro shoot growth and development based on the optimization of light and carbohydrate sources. bioRvix 2021. [Google Scholar] [CrossRef]
- Adhikary, D.; Kulkarni, M.; El-Mezawy, A.; Mobini, S.; Elhiti, M.; Gjuric, R.; Ray, A.; Polowick, P.; Slaski, J.J.; Jones, M.P.; et al. Medical Cannabis and industrial hemp tissue culture: Present status and future potential. Front. Plant Sci. 2021, 12, 275. [Google Scholar] [CrossRef]
- Monthony, A.S.; Kyne, S.T.; Grainger, C.M.; Jones, A.M.P. Recalcitrance of Cannabis sativa to de novo regeneration; a multi-genotype replication study. PLoS ONE 2021, 16, e0235525. [Google Scholar] [CrossRef]
- Rense, S. Here Are All the States That Have Legalized Weed in the U.S. Available online: https://www.esquire.com/lifestyle/a21719186/all-states-that-legalized-weed-in-us/ (accessed on 15 December 2020).
- Galán-Ávila, A.; García-Fortea, E.; Prohens, J.; Herraiz, F.J. Development of a direct in vitro plant regeneration protocol from Cannabis sativa L. seedling explants: Developmental morphology of shoot regeneration and ploidy level of regenerated plants. Front. Plant Sci. 2020, 11, 645. [Google Scholar] [CrossRef]
- Piunno, K.F.; Golenia, G.; Boudko, E.A.; Downey, C.; Jones, A.M.P. Regeneration of shoots from immature and mature inflorescences of Cannabis sativa. Can. J. Plant Sci. 2019, 99, 556–559. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Thidiazuron-induced high-frequency direct shoot organogenesis of Cannabis sativa L. Vitr. Cell. Dev. Biol. 2009, 45, 12–19. [Google Scholar] [CrossRef]
- Wang, R.; He, L.S.; Xia, B.; Tong, J.F.; Li, N.; Peng, F. A Micropropagation system for cloning of hemp (Cannabis sativa L.) by shoot tip culture. Pak. J. Bot. 2009, 41, 603–608. [Google Scholar]
- Chaohua, C.; Gonggu, Z.; Lining, Z.; Chunsheng, G.; Qing, T.; Jianhua, C.; Xinbo, G.; Dingxiang, P.; Jianguang, S. A rapid shoot regeneration protocol from the cotyledons of hemp (Cannabis sativa L.). Ind. Crop. Prod. 2016, 83, 61–65. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; De Klerk, G.J. Plant Propagation by Tissue Culture, 3rd ed.; Springer: Dodrecht, The Netherlands, 2008; p. 501. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Techen, N.; Khan, I.A.; ElSohly, M.A. In vitro mass propagation of Cannabis sativa L.: A protocol refinement using novel aromatic cytokinin meta-topolin and the assessment of eco-physiological, biochemical and genetic fidelity of micropropagated plants. J. Appl. Res. Med. Aromat. Plants 2016, 3, 18–26. [Google Scholar] [CrossRef]
- Boonsnongcheep, P.; Pongkitwitoon, B. Factors affecting micropropagation of Cannabis sativa L.: A review. Pharm. Sci. Asia 2020, 47, 21–29. [Google Scholar] [CrossRef]
- Kodym, A.; Leeb, C.J. Back to the roots: Protocol for the photoautotrophic micropropagation of medicinal Cannabis. Plant Cell, Tissue Organ Cult. 2019, 138, 399–402. [Google Scholar] [CrossRef] [Green Version]
- Grace, D.J.; Roberts, W.J. Method of Producing Pathogen-Free Cannabis Plants and Pathogen-Free Plants and Clones Produced Therefrom. U.S. Patent US20200352124A1, 12 November 2020. Available online: https://www.freepatentsonline.com/y2020/0352124.html (accessed on 16 September 2020).
- Hari, V. Generation of new varieties of Cannabis by chemical mutagenesis of Cannabis cell suspensions. U.S. Patent US20200107510A1, 4 April 2020. Available online: https://www.freepatentsonline.com/y2020/0107510.html (accessed on 17 September 2020).
- Hari, V. Selection of New Varieties of Cannabis Plants Expressing Cannabinoids by Cell Culture (Patent). 2020. Available online: https://www.freepatentsonline.com/y2020/0107511.html (accessed on 16 September 2020).
- Codesido, V.; Meyer, S.; Casano, S. Influence of media composition and genotype for successful Cannabis sativa L. in vitro introduction. Acta Hortic. 2020, 1285, 75–80. [Google Scholar] [CrossRef]
- Wróbel, T.; Dreger, M.; Wielgus, K.; Słomski, R. Modified nodal cuttings and shoot tips protocol for rapid regeneration of Cannabis sativa L. J. Nat. Fibers 2020, 1–10. [Google Scholar] [CrossRef]
- Rojas-martinez, L.; Klerk, G. Hyperhydricity in plant tissue culture. Drowing from within. In Prophyta Annual 2010; Agro Business Solutions: Grootebroek, The Netherlands, 2010; pp. 22–25. [Google Scholar]
- Grulichová, M.; Mendel, P. Effect of Different Phytohormones on Growth and Development of Micropropagated Cannabis sativa L.; MendelNet: Brno, Czechia, 2017; pp. 618–623. [Google Scholar]
- Pan, M.; Van Staden, J. The use of charcoal in in vitro culture—A review. Plant Growth Regul. 1998, 26, 155–163. [Google Scholar] [CrossRef]
- Parsons, J.L.; Martin, S.L.; James, T.; Golenia, G.; Boudko, E.A.; Hepworth, S.R. Polyploidization for the genetic improvement of Cannabis sativa. Front. Plant Sci. 2019, 10, 476. [Google Scholar] [CrossRef]
- Mubi, S.M.; Svetik, S.; FlaJ¡Man, M.; Murovec, J. In Vitro tissue culture and genetic analysis of two high-cbd medical Cannabis (Cannabis sativa L.) breeding lines. Genetika 2020, 52, 925–941. [Google Scholar] [CrossRef]
- Murphy, R.; Adelberg, J. Physical factors increased quantity and quality of micropropagated shoots of Cannabis sativa L. in a repeated harvest system with ex vitro rooting. Vitr. Cell. Dev. Biol. 2021, 1–9. [Google Scholar] [CrossRef]
- Slusarkiewicz-Jarzina, A.; Ponitka, A.; Kaczmarek, Z. Influence of cultivar, explant source and plant growth regulator on callus induction and plant regeneration of Cannabis sativa L. Acta Biol. Crac. Ser. Bot. 2005, 47, 145–151. [Google Scholar]
- Wielgus, K.; Luwanska, A.; Lassocinski, W.; Kaczmarek, Z. Estimation of Cannabis sativa L. tissue culture conditions essential for callus induction and plant regeneration. J. Nat. Fibers 2008, 5, 199–207. [Google Scholar] [CrossRef]
- Movahedi, M.; Ghasemi-Omran, V.; Torabi, S. The effect of different concentrations of TDZ and BA on in vitro regeneration of Iranian Cannabis (Cannabs sativa) using cotyledon and epicotyl explants. J. Plant Mol. Breed. 2015, 3, 20–27. [Google Scholar]
- Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. High frequency plant regeneration from leaf derived callus of high Δ9-Tetrahydrocannabinol yielding Cannabis sativa L. Planta Med. 2010, 76, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I. Studies on the Development of Callus Cultures of Cannabis sativa L. Regarding Plant Regeneration. Available online: https://fenix.tecnico.ulisboa.pt/downloadFile/1970719973967673/extended%20abstract%20final.pdf (accessed on 12 December 2020).
- Raharjo, T.J.; Eucharia, O.; Chang, W.-T.; Verpoorte, R. Callus induction and phytochemical characterization of Cannabis sativa cell suspension cultures. Indones. J. Chem. 2006, 6, 70–74. [Google Scholar] [CrossRef]
- Page, S.R.G.; Monthony, A.S.; Jones, A.M.P. Basal media optimization for the micropropagation and callogenesis of Cannabis sativa L. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Page, S.R.; Monthony, A.S.; Jones, A.M.P. DKW basal salts improve micropropagation and callogenesis compared with MS basal salts in multiple commercial cultivars of Cannabis sativa. Botany 2021, 99, 269–279. [Google Scholar] [CrossRef]
- Monthony, A.S.; Bagheri, S.; Zheng, Y.; Jones, A.M.P. Flower power: Floral reversion as a viable alternative to nodal micropropagation in Cannabis sativa. Vitr. Cell. Dev. Biol. 2021, in press. [Google Scholar] [CrossRef]
- Monthony, A.S.; Page, S.R.; Hesami, M.; Jones, A.M.P. The Past, Present and Future of Cannabis sativa Tissue Culture. Plants 2021, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Zarei, A.; Behdarvandi, B.; Dinani, E.T.; Maccarone, J. Cannabis sativa L. photoautotrophic micropropagation: A powerful tool for industrial scale in vitro propagation. Vitr. Cell. Dev. Biol. 2021, in press. [Google Scholar] [CrossRef]
- Aremu, A.O.; Bairu, M.W.; Dolezal, K.; Finnie, J.; Van Staden, J. Topolins: A panacea to plant tissue culture challenges? Plant Cell, Tiss Org Cult. (PCTOC) 2012, 108, 1–16. [Google Scholar] [CrossRef]
- Donnelly, D.J.; Tisdall, L. Acclimatization strategies for micropropagated plants. In Micropropagation of Woody Plants; Springer: Dodrecht, The Netherlands, 1993; Volume 41, pp. 153–166. [Google Scholar] [CrossRef]
- Hazarika, B. Acclimatization of tissue-cultured plants. Curr. Sci. 2003, 85, 1704–1712. [Google Scholar]
- Mr Grow It. Soil vs Coco Coir—What’s Better for Growing Cannabis? Available online: https://www.mrgrowit.com/post/2018/03/13/soil-vs-coco-coir-whats-better-for-growing-cannabis (accessed on 16 December 2020).
- Zhang, X.; Xu, G.; Cheng, C.; Lei, L.; Sun, J.; Xu, Y.; Deng, C.; Dai, Z.; Yang, Z.; Chen, X.; et al. Establishment of an Agrobacterium-mediated genetic transformation and CRISPR/Cas9-mediated targeted mutagenesis in hemp (Cannabis sativa L.). Plant Biotechnol. J. 2021. [Google Scholar] [CrossRef]
- Smýkalová, I.; Vrbová, M.; Cvečková, M.; Plačková, L.; Žukauskaitė, A.; Zatloukal, M.; Hrdlička, J.; Plíhalová, L.; Doležal, K.; Griga, M. The effects of novel synthetic cytokinin derivatives and endogenous cytokinins on the in vitro growth responses of hemp (Cannabis sativa L.) explants. Plant Cell Tissue Organ Cult. 2019, 139, 381–394. [Google Scholar] [CrossRef]
- Downey, C.D.; Golenia, G.; Boudko, E.A.; Jones, A.M.P. Cryopreservation of 13 commercial Cannabis sativa genotypes using in vitro nodal explants. Plants 2021, 10, 1794. [Google Scholar] [CrossRef]
- Uchendu, E.; Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Cryopreservation of shoot tips of elite cultivars of Cannabis sativa L. by droplet vitrification. Med. Cannabis Cannabinoids 2019, 2, 29–34. [Google Scholar] [CrossRef]
- Qahtan, A.A.; Abdel-Salam, E.; Alatar, A.A.; Wang, Q.-C.; Faisal, M. An Introduction to Synthetic Seeds: Production, Techniques, and Applications; Springer: Cham, The Netherlands, 2019; pp. 1–20. [Google Scholar] [CrossRef]
- Chandrasekhara, R.M. Synthetic seeds: A review in agriculture and forestry. Afr. J. Biotechnol. 2012, 11, 14254–14275. [Google Scholar] [CrossRef]
- Holland, E. Tissue Culture & Synthetic Seeds: Buds without Borders? Available online: https://cannabisnow.com/tissue-culture-synthetic-seeds-buds-without-borders/ (accessed on 15 September 2021).
- Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Propagation through alginate encapsulation of axillary buds of Cannabis sativa L.—An important medicinal plant. Physiol. Mol. Biol. Plants 2009, 15, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lata, H.; Chandra, S.; Techen, N.; Khan, I.A.; ElSohly, M.A. Molecular analysis of genetic fidelity in Cannabis sativa L. plants grown from synthetic (encapsulated) seeds following in vitro storage. Biotechnol. Lett. 2011, 33, 2503–2508. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Jones, A.M.P. Modeling and optimizing callus growth and development in Cannabis sativa using random forest and support vector machine in combination with a genetic algorithm. Appl. Microbiol. Biotechnol. 2021, 105, 5201–5212. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Pepe, M.; Monthony, A.S.; Baiton, A.; Maxwell, A.; Jones, P. Modeling and optimizing in vitro seed germination of industrial hemp (Cannabis sativa L.). Ind. Crops. Prod. 2021, 170, 113753. [Google Scholar] [CrossRef]
- Feeney, M.; Punja, Z.K. Tissue culture and Agrobacterium-mediated transformation of hemp (Cannabis sativa L.). Vitr. Cell. Dev. Biol. 2003, 39, 578–585. [Google Scholar] [CrossRef]
- Wahby, I.; Caba, J.; Ligero, F. Agrobacterium infection of hemp (Cannabis sativa L.): Establishment of hairy root cultures. J. Plant Interact. 2013, 8, 312–320. [Google Scholar] [CrossRef]
- Berahmand, F.; Beizaee, N.; Nayyeri, M.D.; Sharafi, A.; Manjili, H.K.; Danafar, H.; Sohi, H.H. Cannabis sativa L. genetically transformed root-based culture via Agrobacterium rhizogenes. Pharm. Biomed. Res. 2016, 2, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, A.; Yadav, N.S.; Gaudet, D.; Kovalchuk, I. Transient expression of the β-glucuronidase gene in Cannabis sativa varieties. Plant Signal. Behav. 2020, 15, 1780037. [Google Scholar] [CrossRef]
- Deguchi, M.; Kane, S.; Potlakayala, S.; George, H.; Proano, R.; Sheri, V.; Curtis, W.R.; Rudrabhatla, S. Metabolic engineering strategies of industrial hemp (Cannabis sativa L.): A brief review of the advances and challenges. Front. Plant Sci. 2020, 11, 580621. [Google Scholar] [CrossRef] [PubMed]
- Beard, K.M.; Boling, A.W.; Bargmann, B.O. Protoplast isolation, transient transformation, and flow-cytometric analysis of reporter-gene activation in Cannabis sativa L. Ind. Crop. Prod. 2021, 164, 113360. [Google Scholar] [CrossRef]
- Ahmed, S.; Gao, X.; Jahan, A.; Adams, M.; Wu, N.; Kovinich, N. Nanoparticle-based genetic transformation of Cannabis sativa. J. Biotechnol. 2021, 326, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Galán-Ávila, A.; Gramazio, P.; Ron, M.; Prohens, J.; Herraiz, F.J. A novel and rapid method for Agrobacterium-mediated production of stably transformed Cannabis sativa L. plants. Ind. Crop. Prod. 2021, 170, 113691. [Google Scholar] [CrossRef]
- Matchett-Oates, L.; Mohamaden, E.; Spangenberg, G.C.; Cogan, N.O.I. Development of a robust transient expression screening system in protoplasts of Cannabis. Vitr. Cell. Dev. Biol. 2021, in press. [Google Scholar] [CrossRef]
- Kurtz, L.E.; Brand, M.H.; Lubell-Brand, J.D. Production of tetraploid and triploid hemp. HortScience 2020, 55, 1703–1707. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Explant | Explant/Decontamination | Steps and Culture Medium | Experimental Outcome | Pros | Cons | References |
|---|---|---|---|---|---|---|
| Seeds | Seeds: sterilised in 75% (v/v) EtOH for 1 min, rinsed in 5% (v/v) active NaCl for 15 min | Culture initiation PGR-free MS medium | Best explant response (59–70%) and highest number of shoots per explant recorded for shoot tip explants cultured on medium supplemented with TDZ | Did not utilise PGRs with cytokinin activity, which minimised the risk of soma clonal variation | Regeneration was low, 74% of nodal segments and 82% of shoot tips not growing | [61] |
| In vitro shoot tips and nodal segments with one axillary bud without leaves (seedlings) | Shoot induction MS medium + BAP (0.5–2.0 mg/L), TDZ (0.1–0.5 mg/L), mT (0.1–1.0 mg/L) | Best regeneration rate obtained from TDZ at 0.2 mg/L. Nodal segments less responsive and growth of only one shoot per explant regardless of the tested PGR | Shorter micropropagation duration time. Does not require elongation step | TDZ use related to phenotypic vitrification, leaf rolling, leaf narrowing and supressed growth of shoots. High BAP and mT concentrations also related to phenotypic changes in regenerated plants | [61] | |
| In vitro plantlets | Rooting ½ MS medium + IAA (0.25–0.75 mg/L) and or IBA (0.25–0.75 mg/L) | No significant difference observed in tested auxins in terms of rooting rates | It can be used for germplasm conservation and breeding. Rooting limited to 21 days due to rapid growth of shoots in culture. Plantlets obtained within 66–70 days | Number of plantlets from single explant was low. Protocol thus not suitable for industrial application | [61] | |
| Seeds | Seeds: surface sterilised in 75% (v/v) ethanol for 2 min and 30 s, soaked in NaClO for 25 min | Culture initiation ½ MS medium | Hypocotyl was significantly better than cotyledon leaves in terms of shoot organogenic potential | This is the first report of direct in vitro regeneration of plants from hypocotyls | Leaves displayed a poor ability to promote shoot organogenesis | [47] |
| In vitro cotyledons, hypocotyls and true leaves | Shoot induction Medium + TDZ (0.4–1.0 mg/L), NAA (0.02–0.2 mg/L), BAP (0.5–2.0 mg/L), IBA (0.5 mg/L), 2,4-D (0.1 mg/L), ZEARIB (1.0–2.0 mg/L), BAPRIB (1.0 mg/L), 4-CPPU (1.0 mg/L) | Medium containing (TDZ 0.4 mg/L + NAA 0.2 mg/L) was the best, achieving the highest shoot induction rate of 22.32% | None | Medium without PGRs and ZEARIB 1 mg/L + NAA 0.02 mg/L were the worst treatments, without any explant showing response in terms of shoot organogenesis | [47] | |
| Seeds | Seeds: surface sterilised by washing under running water with a few drops of detergent, 0.2% mercury chloride for 13 min | Culture initiation MS medium | Plantlets were grown from seeds | None | None | [63] |
| In vitro shoot tips | Shoot induction MS medium + BAP (0.4 mg/L)/TDZ (0.1 mg/L)/mT (0.5 mg/L) + NAA (0.1 mg/L)/IAA (0.1 mg/L)/GA3 (2.3 mg/L) | In both varieties, the highest stem was observed when cultured on medium supplemented with TDZ and GA3, and the shortest stem recorded on medium supplemented with TDZ and NAA | None | The presence of NAA strongly influenced callus formation and general shoot architecture. Difficult to tell which extent longer stems are a genotypic trait | [63] | |
| In vitro plantlets | Rooting MS medium + IBA (0.5 mg/L) + activated charcoal | The most vital plantlets of both genotypes with the highest number of roots were observed on medium where phytohormones were not present or on medium supplemented with mT (0.5 mg/L) | Culture media supplemented with mT without any phytohormones produced the best overall appearance of plantlets | None | [63] | |
| Seeds | Seeds: soaked with H2SO4 for 20 s, sterilised in 75% ethanol for 2 min, 3% (v/v) NaClO for 20 min | Culture initiation MS medium | Seeds grew up to seedlings and cotyledons were excised as explants to induce in vitro shoots | None | None | [52] |
| Cotyledons excised from seedlings (aseptic seedlings obtained from sterilised seeds) | Shoot induction MS medium + TDZ (0.1–0.4 mg/L), BA (4–8 mg/L), ZT (0.5–1.5 mg/L) with or without NAA (0.1–0.6 mg/L) | Cotyledon cultured in medium containing TDZ with or without the addition of NAA were capable of inducing formation of a nodular callus. Induction rate lower when using only TDZ. Peak of 51.7% induction frequency in MS medium + TDZ (0.4 mg/L) + NAA (0.2 mg/L) | Rapid shoot regeneration. No limitation of cultural season due to the use of cotyledons | This regeneration protocol is genotype dependent | [52] | |
| In vitro shoots | Rooting ½ MS Medium with IBA (0.2–2 mg/L) | IBA (0.5–2 mg/L) had 80% root induction | None | None | [52] | |
| Seeds | Seeds: washed for 20 min with 0.1% antiseptic APSA80 liquid detergent, sterilised in 75% (v/v) ethanol for 30 s and 0.1% mercuric chloride for 10–15 min | Culture initiation ½ MS medium with 10 g/L sucrose and 5.5 g/L agar | Shoot tips were harvested from 20-day-old sterile plantlets | None | None | [51] |
| Shoot tips harvested from 20-day-old sterile plantlets (aseptic seedlings obtained from sterilised seeds) | Regeneration MS medium + BA (1–5 mg/L), KT (1–5 mg/L), TDZ (1–5 mg/L) with or without NAA (1–5 mg/L) | TDZ (0.2 mg/L) provided the best bud induction, producing an average of 3.22 buds. 0.1 mg/L NAA was optimal concentration for auxiliary bud induction | CKs stimulated shoot formation and stem enlargement in each explant | Type of CK affected plantlet morphology | [51] | |
| In vitro plantlets | Rooting MS medium + IBA (0.1–0.5 mg/L) + NAA (0.05–0.25 mg/L), IAA (0.05–0.25 mg/L) | 85% rooting response in IBA (0.1 mg/L) and NAA (0.05 mg/L) explants | None | None | [51] | |
| Axillary buds | Axillary buds: surface disinfected by maintaining them under stirred tap water for 1 h; 30 min immersion in 15% (v/v) bleach, stirred solution | Culture initiation and shoot induction MS medium with or without vitamins/Formula βH/Formula βA + 0.48 mg/L mT or 0.37 mg/L NAA + 0.41 mg/L IBA with or without MS basal salts, Formula βH basal salts, Formula βA basal salts, with or without MS vitamins | 100% survival of axillary buds was observed for all cultivars at least under one studied media. Most of the varieties survived and reacted better without the addition of MS vitamins. Use of PGRs was variety dependent: some cultivars responded better to the addition of mT instead to NAA+IBA | This study confirmed that the success of in vitro introduction of C. sativa is cultivar dependent | Different cultivars of the same species have a completely different response to the same medium | [60] |
| Nodal segments with axillary buds | Nodal segments containing young axillary buds: sterilised in 2% NaOCl, 0.1% (v/v) Tween 20 for 5 min | Culture initiation MS medium + activated charcoal | None | None | None | [65] |
| In vitro explants | Shoot induction MS medium + 0.1 mg/L NAA + 0.4 mg/L kinetin | None | None | None | [65] | |
| In vitro plantlets | Rooting MS medium + 0.1 mg/L NAA + 0.4 mg/L kinetin + 1.0 mg/L IBA | None | None | None | [65] | |
| Disinfected axillary buds | Oryzalin treatments Shoot induction medium + 17.32, 34.62, 51.95 mg/L oryzalin or MS medium + 6.93, 13.85, 20.78 mg/L oryzalin | 62.5% to 87.5% survival rate for explants treated with 6.92 mg/L oryzalin | The treatment of axillary buds with oryzalin is an effective method for chromosome doubling | Poor survival rate of explants treated with high oryzalin concentrations with 0% of explants surviving the 51.95 mg/L | [65] | |
| Shoot tips | In vitro shoot tip cuttings | Maintenance of stock plants in ventilated glass jars ¼ Rockwool block placed onto glass preservation jars (3 shoot tip cuttings for each block) | The self-built preservation jars were more suited for the culture of Cannabis as they provided more head space | The stock cultures could be maintained for at least 6 months. Excellent-quality plantlets | Wilting plants (blocks too dry/humidity too low). Deterioration of plants due to the blocks being too wet | [56] |
| In vitro shoot tip cuttings | Maintenance of stock plants using RITA® system Nutrient solution (20 mL), Canna Aqua Vega Fertiliser. RITA container with 3 rockwool blocks each (2 shoot tip cuttings in each block), nutrient solution (75 mL), jars connected via tubing to a 1 bar pressure pump | The RITA® system was more practicable in terms of handling because of the wide opening | Relies on industry-based fertiliser, rockwool blocks and forced ventilation. No requirement of growth regulators. No sugar or vitamins required | Stunted plants or yellow leaves (nutrient deficiency) | [56] | |
| In vitro shoot tips | Rooting Glass vessel, 2 rockwool blocks, nutrient solution (20 mL) | 97.5% of in vitro shoot tip cuttings were rooted and acclimatised within 3 weeks inside the growth chamber | None | None | [56] | |
| Shoots | Shoots from immature and mature inflorescences: surface sterilised in ethanol for 1 min, followed by 10% v/v bleach for 10 min, washed in sterile water for 50 s | Culture initiation MS medium + TDZ (0–2.2 mg/L) | TDZ was shown to be among the most effective PGRs for shoot proliferation and de novo regeneration | First known report of shoot regeneration from floral tissues | None | [48] |
| In vitro explants with regenerating shoots | Shoot regeneration/rooting MS medium + KIN (0.40 mg/L) + NAA (0.10 mg/L) + activated charcoal | Regeneration was occurring from existing meristematic tissue, but this was not specifically determined | First report of shoot regeneration or plant propagation at reproductive phase | Further work needed to refine the protocol | [48] | |
| Nodal segments with axillary buds | Nodal segments containing axiliary buds: disinfected with 0.5% NaOCl for 20 min | Shoot induction MS medium + TDZ (0.01–1.10 mg/L) + 500 mg/L activated charcoal | In TDZ, of the different concentrations tested, the highest average number of shoots was obtained in MS + 0.5 µM TDZ | One step protocol for promoting shoot formation and root induction in the same medium | None | [54] |
| In vitro explants with regenerating shoots | Shoot formation/Rooting ½ MS medium + IBA (0.01–1.01 mg/L), mT (0.01–1.21 mg/L) | 100% of explants exposed to with 0.48 mg/L mT produced shoots. Shoot number and shoot length was higher when using mT compared to TDZ. The best concentration for rooting was 0.05 mg/L mT | High shoot proliferation rate. Proof of the safety of mT for large-scale production. 96% of regenerated shoots were able to develop roots | mT concentrations higher than 0.97 mg/L were inhibitory to rooting | [54] | |
| Nodal segments with axillary buds | Nodal segments containing auxiliary buds: sterilised using 0.5% NaOCl for 20 min | Shoot induction MS medium + BA (0.01–2.03 mg/L), KN (0.01–1.94 mg/L), TDZ (0.01–1.98 mg/L) with or without GA3 (2.42 mg/L) | TDZ was the most effective PGR for shoot proliferation. 100% culture response when using TDZ (0.11 mg/L), with an average of 13 shoots per explant | Regeneration of many plants in a short period of time. GA3 can act as a replacement for auxins in shoot induction | TDZ concentrations higher than 1.1 mg/L supressed shoot formation | [50] |
| In vitro shoots | Rooting MS medium + IAA (0.44–0.88 mg/L), IBA (0.51–1.02 mg/L), NAA (0.47–0.93 mg/L) with or without 500 mg/L activated charcoal | 94% response of cultures in IBA (0.51 mg/L) with an average of 4.8 roots per explant | Addition of activated charcoal was effective in root induction | Profuse callus formation was observed when using IAA and IBA | [50] | |
| Nodal segments with axillary buds | Apical nodal segments containing axillary bud: sterilised using 0.5% NaOCl for 20 min | Shoot initiation MS medium + BA, KN, TDZ (concentrations not mentioned) | Quality and quantity of shoot regenerants in cultures were best with 0.11 mg/L TDZ | None | None | [64] |
| Apical nodal segments containing axillary bud: sterilised using 0.5% NaOCl for 20 min | Rooting ½ MS medium + activated charcoal + IAA + IBA + NAA (concentrations not mentioned) | Highest percentage of rooting achieved in ½ MS with 500 mg/dm3 activated charcoal supplemented with 0.51 mg/L IBA | None | None | [64] | |
| Nodal segments with axillary buds | Nodal segments containing axillary buds: sterilised using 1.67% (C(O)NCl)₂ + Tween 20 for 8 min | Shoot initiation: MS + TDZ (0.011– 1.76 mg/L), mT (0.012–1.93 mg/L), BAP (1–5 mg/L), IAA (0.1 mg/L) | MS medium + 0.1 mg/L TDZ resulted in the highest regeneration of shoots. Tissue culture responsiveness was genotype dependent | None | Results demonstrated the recalcitrance of Cannabis in tissue culture and its poor multiplication rate | [66] |
| Apical shoot tip | Apical shoot tip+ node | Shoot initiation: DKW medium without PGRs | The highest number of harvested shoot tips was found in the 46 µmol/m2/s in non-vented vessels | Unlike traditional micropropagation, this method re-uses the same rooted basal stem section of the initial explant over several apical tip removal cycles, resulting in a higher number of shoot tips | None | [67] |
| Explant | Explant/Decontamination | Steps and Culture Medium | Experimental Outcome | Pros | Cons | References |
|---|---|---|---|---|---|---|
| Seeds | Seeds: Sterilised in 5% Ca (ClO)2 for 6, 8 and 15 min | Culture initiation MS medium | Best sterilisation time was achieved after 15 min (5% hypochlorite solution) | None | Hemp seeds were highly contaminated | [68] |
| In vitro young leaves, petioles, internodes and axillary buds | Callus induction/indirect regeneration MS medium + KN (1–4 mg/L), NAA (0.5–2 mg/L), 2,4-D (2–4 mg/L), DIC (2–3 mg/L) | Callus was obtained from all explant types. Petiole explants with 2–3 mg/L DIC had the highest frequency of callus formation with 82.7% of explants | Explants derived from plants growing in pots | Low frequency of callus from internodes and axillary buds. Efficiency of plant regeneration is low | [68] | |
| In vitro regenerated plantlets | Rooting MS medium + IAA (1 mg/L) and NAA (1.0 mg/L) | 69.95% of the plantlets formed roots | None | Further experiments needed to develop an efficient plant regeneration system | [68] | |
| Seeds | Seeds: sterilised in 70% ethanol for 10 s and in 1% NaClO for 20 min | Culture initiation DARIA medium | Explants of cotyledons, stems, and roots were excised from plantlets | None | None | [69] |
| In vitro cotyledons, stems, roots | Callus induction DARIA medium + KN (1 mg/L) + NAA (0.05 mg/L) | Callus was obtained from all explant types | The highest efficiency of morphogenic callus induction was noticed from cotyledon explants | Callus formed at root explants was incapable of morphogenesis and plant regeneration | [69] | |
| In vitro explants | Indirect regeneration DARIA medium + BA (0.2 mg/L) + NAA (0.03 mg/L) | Stem explants showed the highest regeneration rate percentage and cotyledon explants showed the highest efficiency in callus induction | The use of three media, DARIA ind+, DARIA pro +, and DARIA root +, supplemented with PGRs, enabled regeneration of plants with relatively high efficiency | None | [69] | |
| In vitro explants | Rooting DARIA medium + IAA (2 mg/L) | Rooted plants were transferred to soil | None | None | [69] | |
| Seeds | Seeds: sterilised with 70% ethanol for 30 s, 2% NaOCl for 20 min and 0.05% HgCl2 for 5 min | Culture initiation MS medium | Seeds produced seedlings for obtaining explants | None | None | [70] |
| In vitro cotyledon and epicotyl | Callus induction MS medium + BA (0.1–3 mg/L), TDZ (0.1–3 mg/L) with or without IBA 0.5 mg/L | Cotyledon explant showed better response compared to epicotyl explants in terms of the mass and size of the calli produced in various hormonal combination | The first response of explant to callus formation was observed after 11 days. The addition of IBA in various concentrations of BA had positive influence on callus induction | None | [70] | |
| In vitro calli | Shoot induction MS medium + BA (0.1–3 mg/L), TDZ (0.1–3 mg/L) with or without IBA 0.5 mg/L | Epicotyl explants showed better regeneration rate compared to cotyledon. Epicotyl explant callus treated with 2 mg/L BA and 0.5 mg/L IBA showed high shoot regeneration rate | None | None | [70] | |
| In vitro regenerated shoots | Rooting MS medium + NAA (0.1–1 mg/L), IBA (0.1–1 mg/L) | IBA (0.1 mg/L) showed highest rooting rate | None | Burning was observed in the shoots cultured in media supplemented with NAA hormone | [70] | |
| Young leaves | Young leaves: sterilised using 0.5% NaOCl, 15% (v/v) bleach | Culture initiation/callus induction MS medium + IAA (0.09–0.35 mg/L), IBA (0.1–0.41 mg/L), NAA (0.09–0.37 mg/L), 2,4-D (0.11–0.44 mg/L) with 0.22 mg/LTDZ | Optimum callus growth in 0.09 mg/L NAA + 0.22 mg/L μM TDZ | Rapid protocol for producing plantlets from young leaf tissue | The formation and growth of the callus was affected by the type of PGR and concentration applied | [71] |
| In vitro calli | Shoot induction MS medium + BAP (0.11–2.25 mg/L), KN (0.12–2.15 mg/L), TDZ (0.11–2.2 mg/L) | Highest shoot induction and proliferation was observed in 0.11 mg/L TDZ | None | None | [71] | |
| In vitro regenerated shoots | Rooting ½ MS medium + IAA (0.09–1.75 mg/L), IBA (0.10–2.03 mg/L), NAA (0.09–1.86 mg/L) | Shoots rooted best in ½ MS medium with 0.51 mg/L IBA. The presence of IBA resulted in significantly higher rooting percentage (80–96%) than IAA or NAA | None | None | [71] | |
| Leaves, flowers, 4-day-old seedlings | Leaves, flowers, and 4-day-old seedlings: washing with detergent, 70% EtOH for 3 min, sterilised distilled water for 10 min, 2% NaClO soak for 20 min | Culture initiation/callus induction MS medium + mesoinositol (100 mg/L), thiamine diHCl (10 mg/L), pyridoxine HCl (1 mg/L), nicotinic acid (1 mg/L), 2,4-D (1 mg/L), sucrose (30 g/L) and agar (10 g/L) | Flowers gave more callus while the leaves had less callus production | Callus was easily induced in standard medium | Cannabinoids were not produced in Cannabis cell cultures | [73] |
| In vitro calli | Suspension cultures Liquid MS medium after 2 weeks: one part was maintained in the MS medium while the other was maintained in B5 medium (B5 components, 2,4-D 2.0 mg/L, IAA 0.5 mg/L, NAA 0.5 mg/L, K 0.2 mg/L and sucrose 30 g/L) | Shoots from seedlings produced more callus than the stems and no callus was formed on the roots | None | None | [73,76] | |
| Immature embryo hypocotyls, true leaves, cotyledons and hypocotyls | Immature embryo hypocotyls, true leaves, cotyledons and hypocotyls: sterilised using 2% (v/v) NaClO for 25 min followed by 75% (v/v) EtOH for 5 min | Culture initiation: MS+ nicotinic acid (1 mg/L) + pyridoxine-HCl (1 mg/L) + thiamine-HCl (10 mg/L) + myo-inositol (0.1 g/L) + 3% sucrose + phytagel (2.5 g/L) + 2,4-D (1 mg/L) + KIN (0.25 mg/L) + casein (100 mg/L) Hydrolysate regeneration: 1/2 strength MS + 1.5% sucrose + phytagel (3.5 g/L) + TDZ (0.5 mg/L) + 6-BA (0.3 mg/L) + NAA (0.2 mg/L) + IAA (0.2 mg/L) Rooting: 1/2 strength MS + NAA (0.2 mg/L) + IBA (0.5 mg/L) + ZeaRIB (0.01 mg/L) | Over 20% of the immature embryo hypocotyls developed embryogenic calli within 5 days. Hypocotyls collected 15 days after anthesis produced more calli than those collected earlier or later | None | Genotype dependence of Cannabis | [83] |
| Leaf | Leaf material from in vitro shoots: no sterilisation mentioned | Culture initiation/callus induction MS + TDZ (1.0 μM) Shoot induction MS + TDZ (0.5 μM) | Callus was effectively induced in all 10 genotypes, yet the subsequent transfer of calli to shoot induction medium failed to initiate shoot organogenesis in any of the tested genotypes. Regeneration of Cannabis from somatic tissues is highly genotype specific | None | This method is not suitable for inducing de novo regeneration across different genotypes | [45] |
| Plantlet Growth Stage | Growth Conditions | Experimental Outcome | References |
|---|---|---|---|
| Direct Organogenesis | |||
| Plantlet (21 days old) | -Pots with sterilised soil -Under a plastic cover -25 ± 1° C (18/6 photoperiod, 60 µmol m−2 s−1) -Hardened for 2 weeks before transferring to the field | 95% survival rate in the growing chamber 90% survival rate in field conditions Plantlets maintained ability to synthesise cannabinoids | [61] |
| Spontaneously rooted plantlets | -Pots (2 L) with fertilised commercial substrate (black peat, granulated peat moss and perlite) - Regenerants received foliar pulverisation with water -Small plants were covered with plastic vessels and were progressively exposed to the environmental humidity - 22 ± 1 °C - 60% ± 1% relative humidity | After 1 week of progressive exposition of regenerants to the environmental humidity, the process of acclimatisation ended, and hypocotyl-derived plants displayed a vigorous growth Hypocotyl derived plants showed sexual functionality 8 weeks after in vitro explant inoculation | [47] |
| Plantlet (age not defined) | -Kept under controlled environmental conditions in an indoor cultivation facility -Well rooted plants washed with tap water to remove all traces of medium -Plants pre-incubated in coco natural growth medium for 10 days before transferring in sterile potting mix-fertilome in large pots -25–30 °C -Light, ∼700 µmol m−2 s−1 with 16 h photoperiod -60% relative humidity | Plants propagated with mT rooted better when transferred to soil than the shoots produced with TDZ 100% survival rate in acclimatised plants | [54] |
| Plantlet (age not defined) | -Kept in a greenhouse -Plantlets with well-developed roots removed from tissue culture vessel and washed under running water -Propagated in plastic cups containing sterilised organic manure, clay soil and sand (1:1:1) -22 °C -Cool white, fluorescent lights (16/8 h photoperiod, 36 µmol m−2 s−1) | 75% of rooted shoots survived after acclimation | [52] |
| Rooted shoots (age not defined) | -Kept in controlled environmental conditions grown in an indoor cultivation facility -Rooted shoots were carefully taken out of the medium and washed thoroughly running tap water -Plantlets were pre-incubated in coco natural growth medium thermocol cups for 10 days -Cups were covered with polythene bags to maintain humidity and later acclimatised in sterile potting mix-fertilome -A hot air suction fan was attached with approximately 1 m distance between plants -16 h photoperiod -25–30 °C -60% humidity | 95% survival of rooted plantlets transferred to soil New growth observed after 2 weeks Plants reached 14–16 cm in height within 6 weeks of transfer Plants showed normal development and no gross morphological variation | [50] |
| Plantlets | -Rooted shoots were carefully taken out of the medium and washed thoroughly in running tap water followed by washings with 0.2% (w/v) Bavistin1 and tap water -Washed plantlets were transferred to root trainers consisting of 20 cells, each of 200 cm3, filled with perlite and 10 mL water -Plantlets were transferred to plastic pots filled with vermiculite and plant ash, grown in a shade-house -After an acclimation period of 2 weeks, the plantlets were able to be transplanted to the field | 95% plants acclimatised 99% plantlet survival for 3 months after field transfer | [51] |
| Plantlets | -Rooted plantlets were placed in Grodan Gro-Smart Tray Insert (Indoor Growing Canada, Montreal, Canada) in the standard tray with transparent dome (Mondi, BC, Canada) with vents. -The plants were fertilised using SF vegetative fertiliser solution. -Rooted plants received photoperiod and light intensity conditions (150 μmol m−2 s −1 and 18/6 h light/dark). | Survival rate above 90% Up to 2260 rooted plantlets were produced per 10 m2 | [78] |
| Indirect Organogenesis | |||
| Rooted shoots | -Cultivated in pots containing equal ratio of perlite and pit moss -To avoid evaporation, the pots were covered with a transparent cover and placed in growth chambers -25 °C -Covers removed after two weeks and plants were transferred into the greenhouse | 70% of the seedlings produced in tissue culture conditions survived and showed normal growth | [70] |
| Plantlets | -Rooted shoots were carefully taken out of the medium and washed thoroughly in running tap water -Plantlets were pre-incubated in coco natural growth medium thermocol cups for 10 days -Growth cups were covered with polythene bags to maintain humidity, kept in a grow room, and later acclimatised in sterile potting mix (fertilome) in large pots -25 °C | 95% survival rate in indoor grow room | [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adams, T.K.; Masondo, N.A.; Malatsi, P.; Makunga, N.P. Cannabis sativa: From Therapeutic Uses to Micropropagation and Beyond. Plants 2021, 10, 2078. https://doi.org/10.3390/plants10102078
Adams TK, Masondo NA, Malatsi P, Makunga NP. Cannabis sativa: From Therapeutic Uses to Micropropagation and Beyond. Plants. 2021; 10(10):2078. https://doi.org/10.3390/plants10102078
Chicago/Turabian StyleAdams, Tristan K., Nqobile A. Masondo, Pholoso Malatsi, and Nokwanda P. Makunga. 2021. "Cannabis sativa: From Therapeutic Uses to Micropropagation and Beyond" Plants 10, no. 10: 2078. https://doi.org/10.3390/plants10102078
APA StyleAdams, T. K., Masondo, N. A., Malatsi, P., & Makunga, N. P. (2021). Cannabis sativa: From Therapeutic Uses to Micropropagation and Beyond. Plants, 10(10), 2078. https://doi.org/10.3390/plants10102078