Sterols and Triterpenes: Antiviral Potential Supported by In-Silico Analysis


 ,
,  ,
,  , and
, and 
Abstract
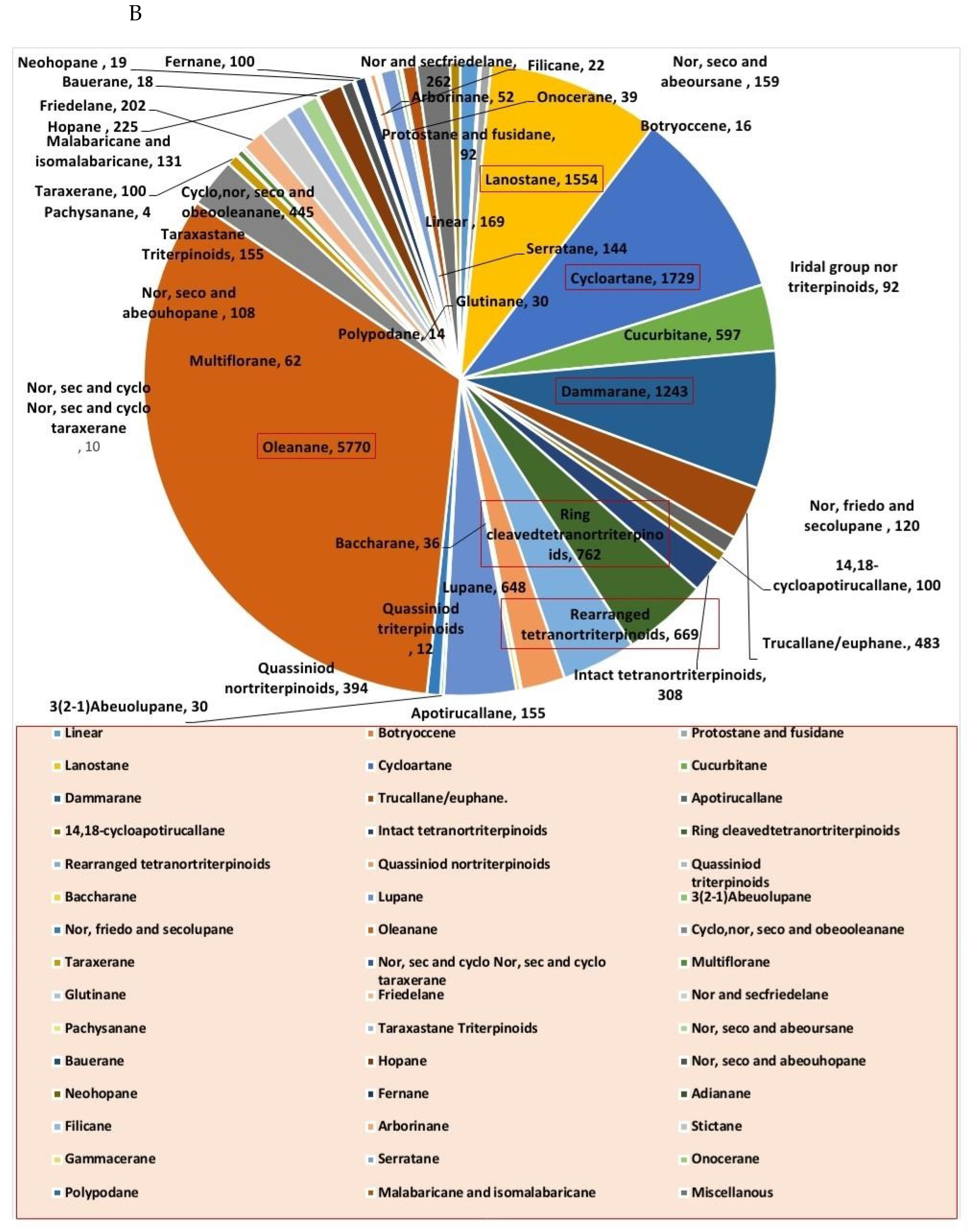
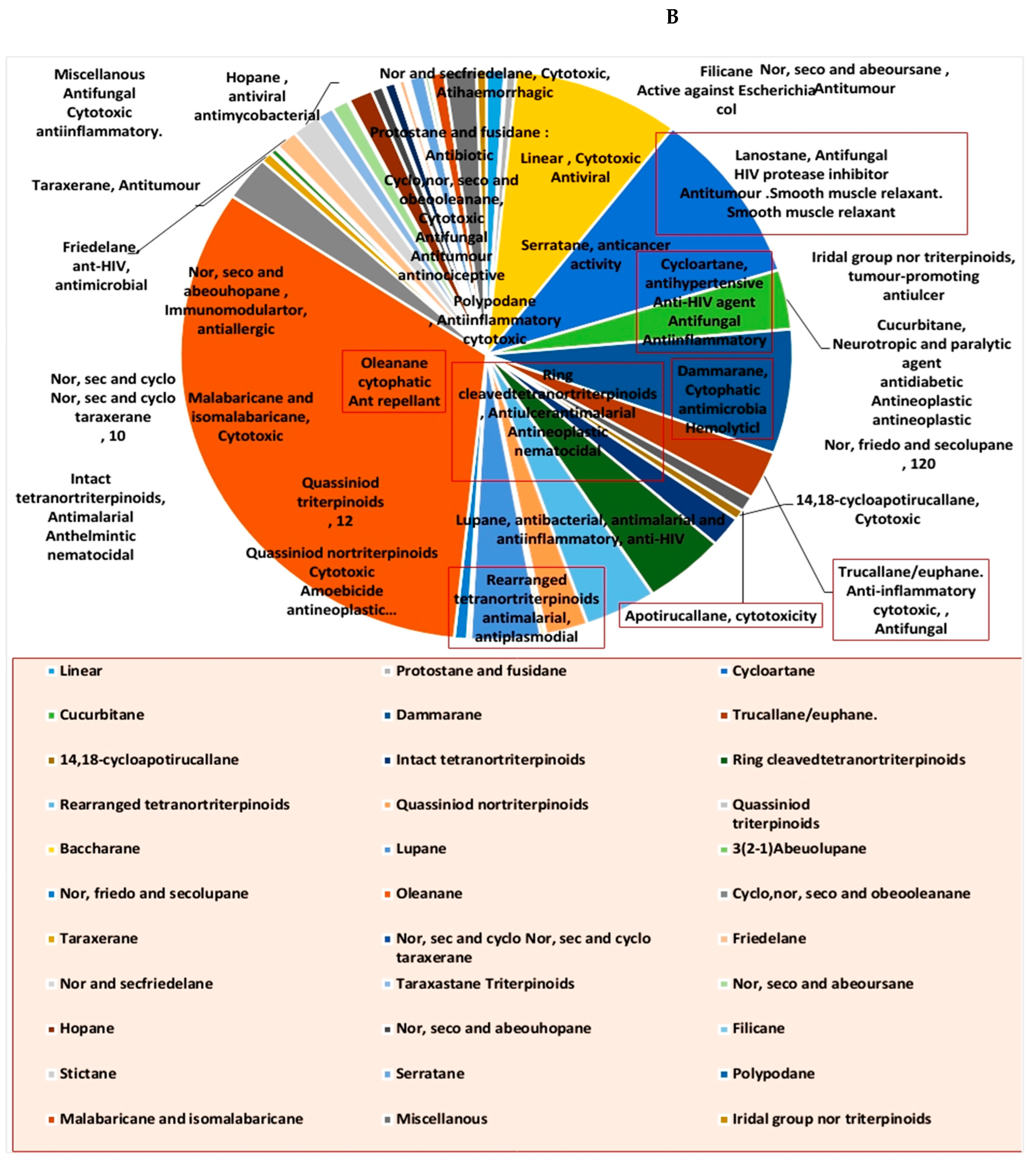
1. Introduction
2. Methodology
2.1. Databases
2.2. Molecular Docking
3. Findings and Discussion
3.1. Anti-HIV Agents
3.1.1. Reverse Transcriptase Inhibitors
3.1.2. Protease Inhibitors
3.1.3. DNA Polymerase Inhibitors
3.2. Replication Inhibitors:
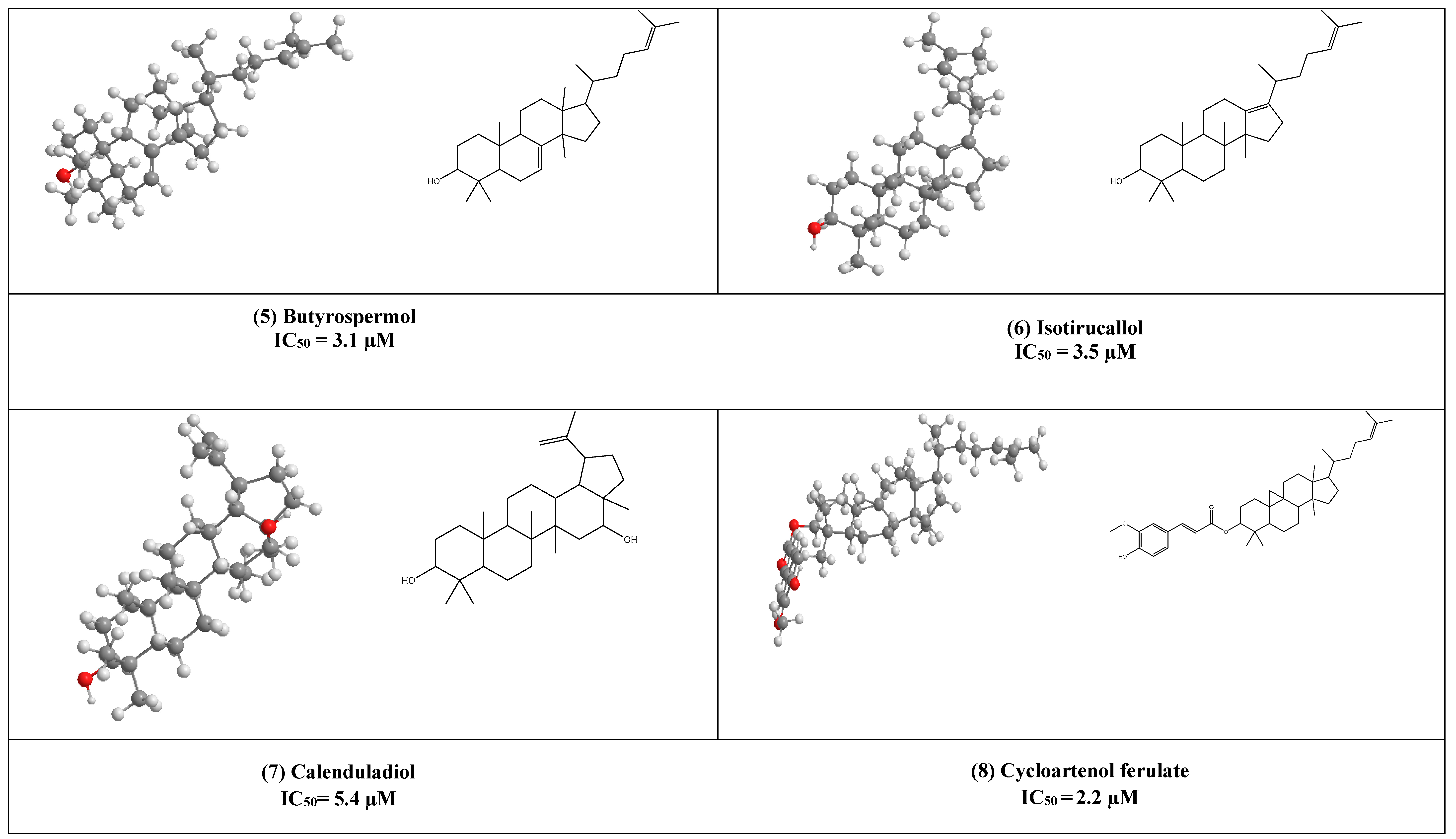
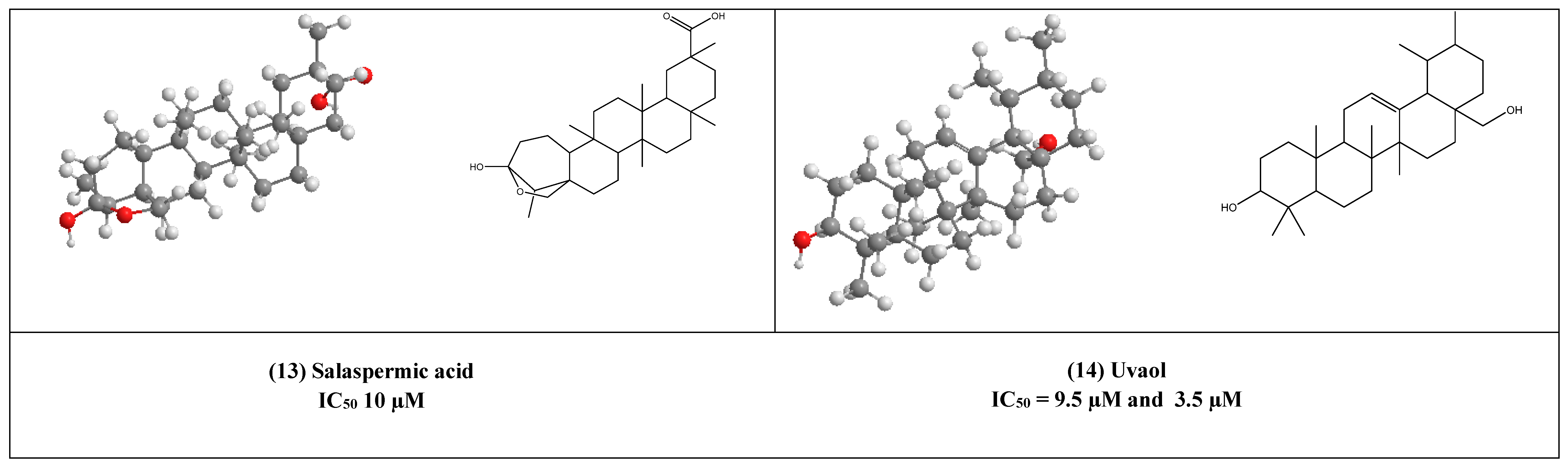
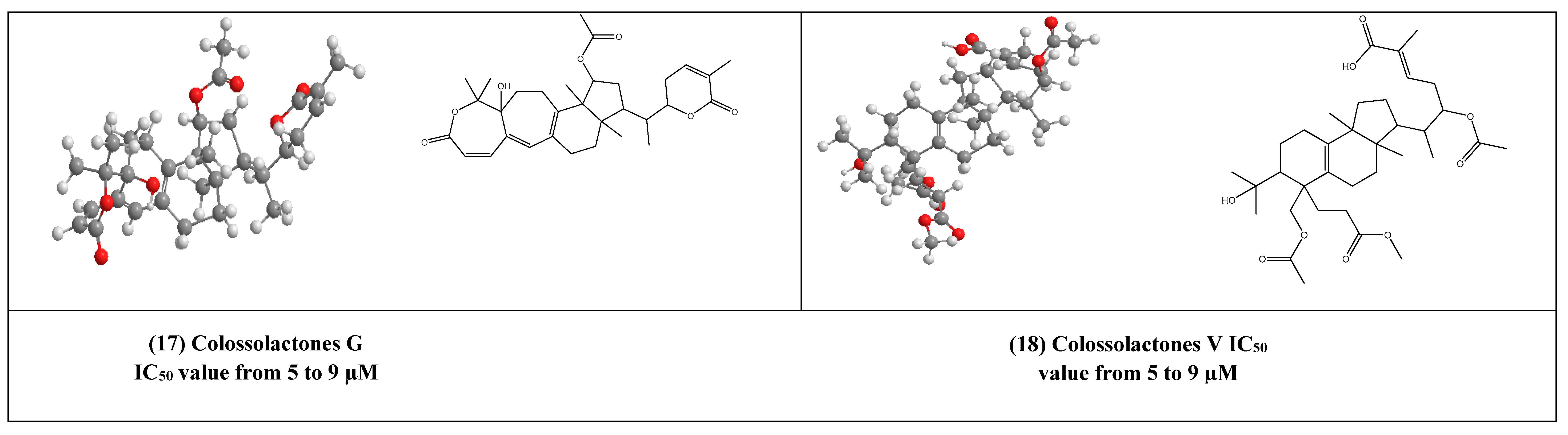
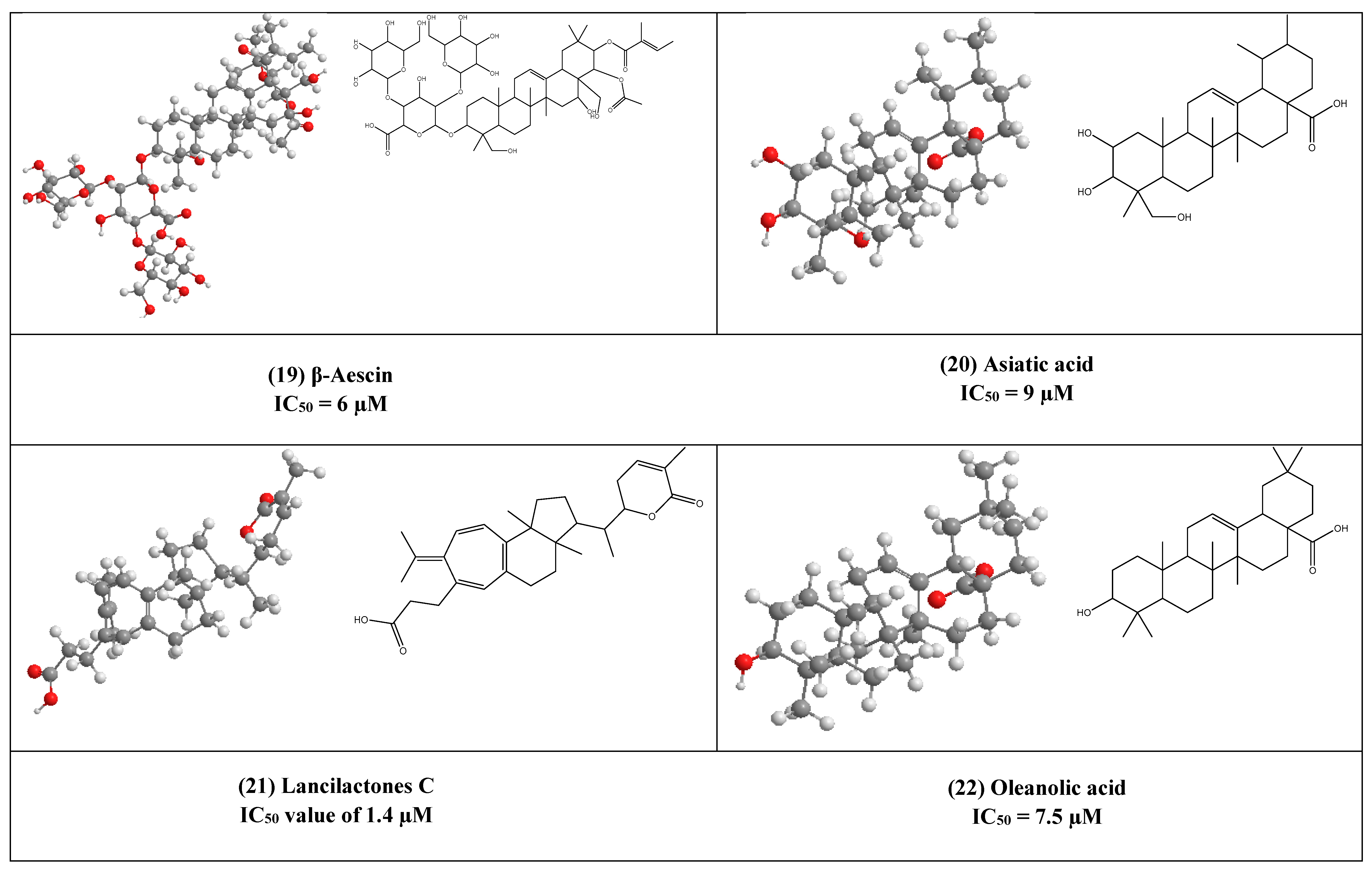
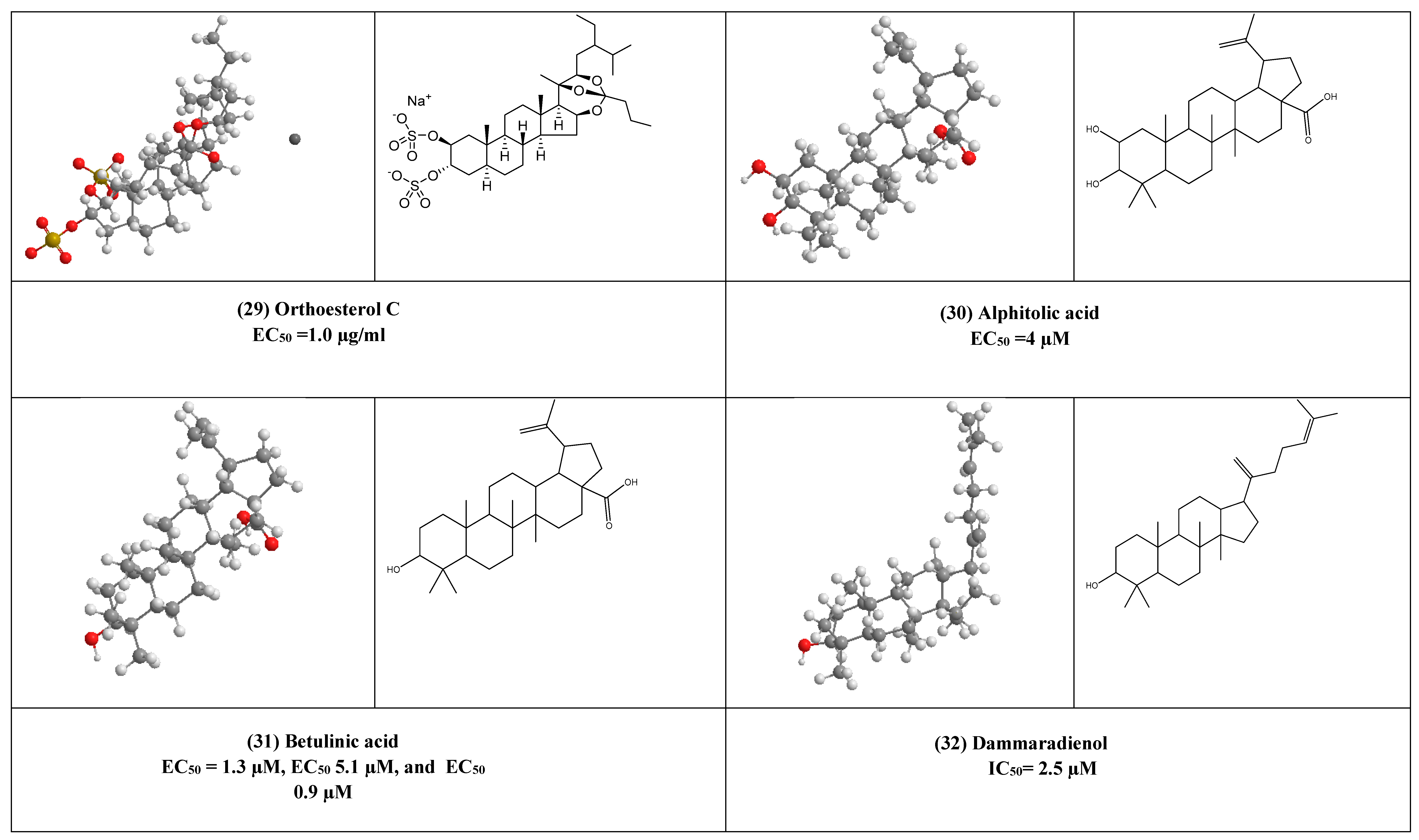
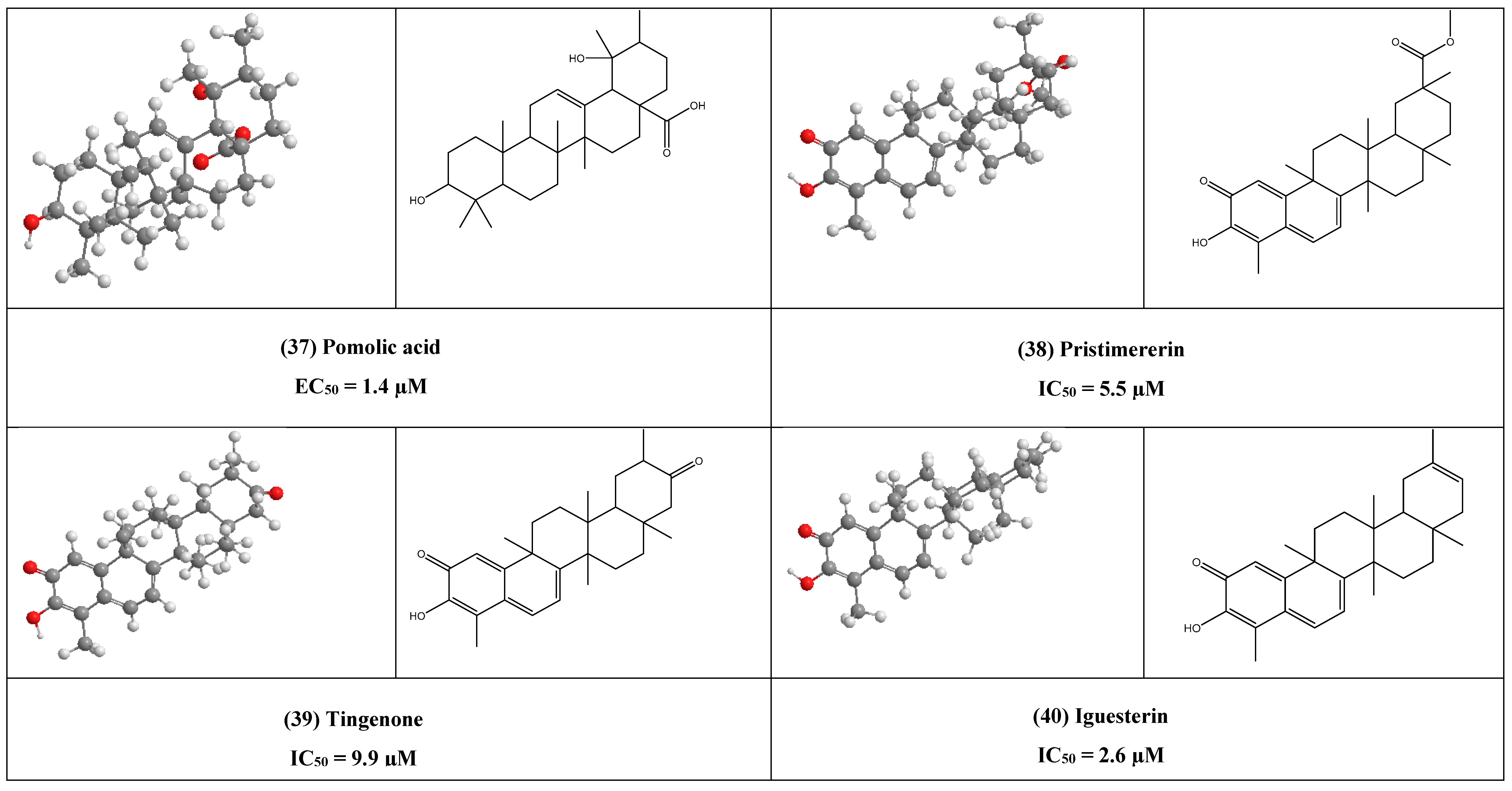
4. Other Compounds with Antiviral Activity
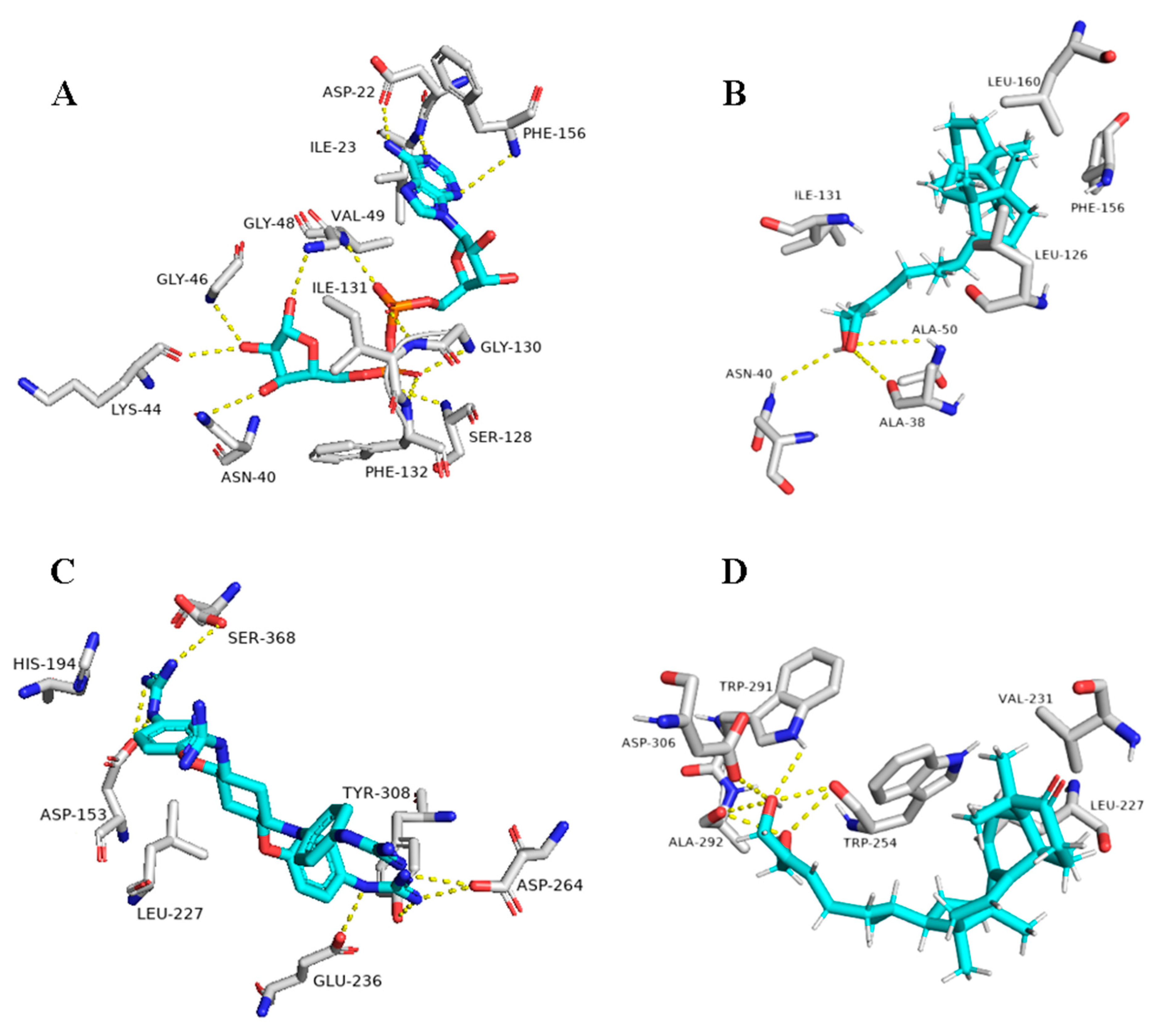
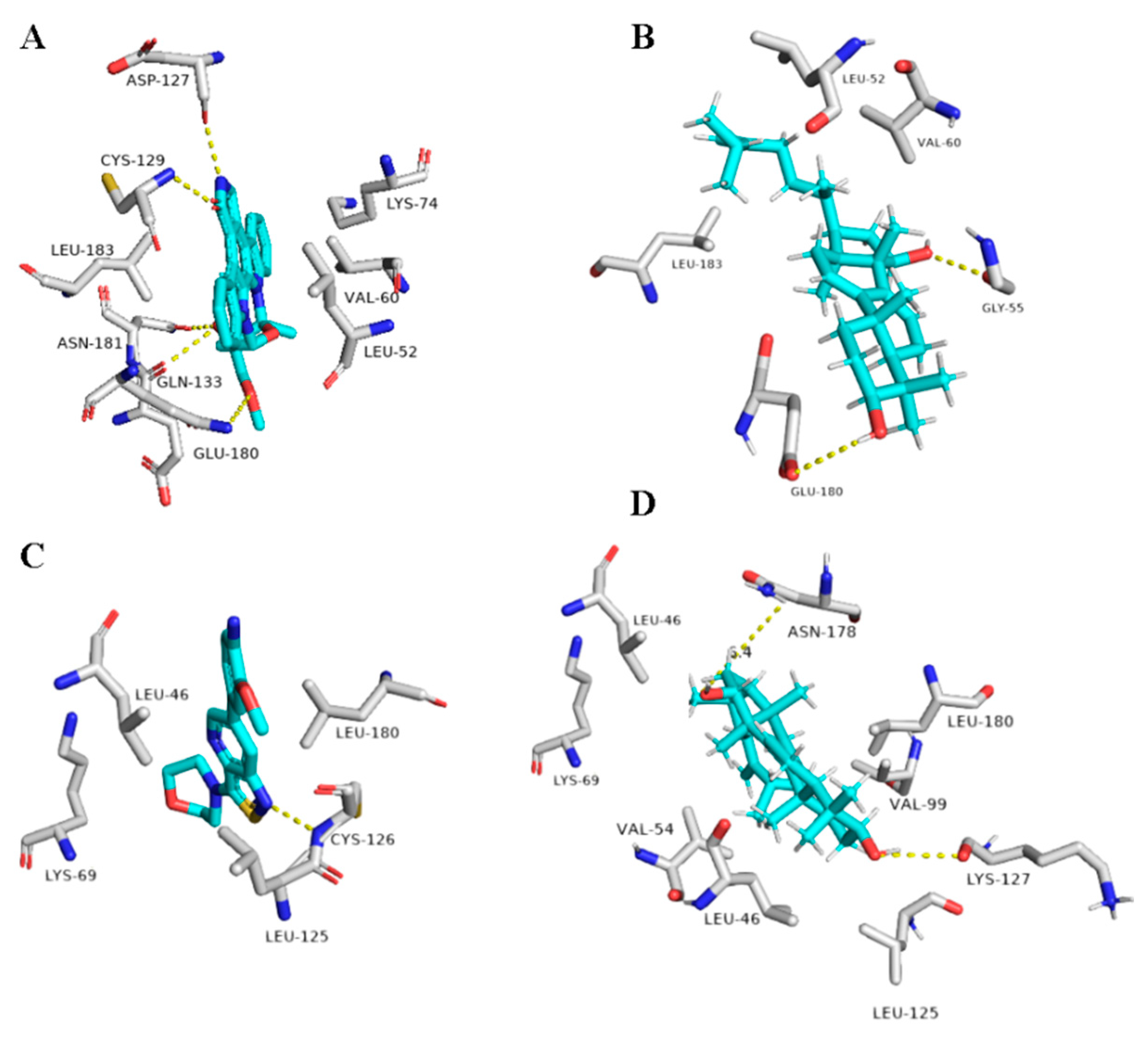
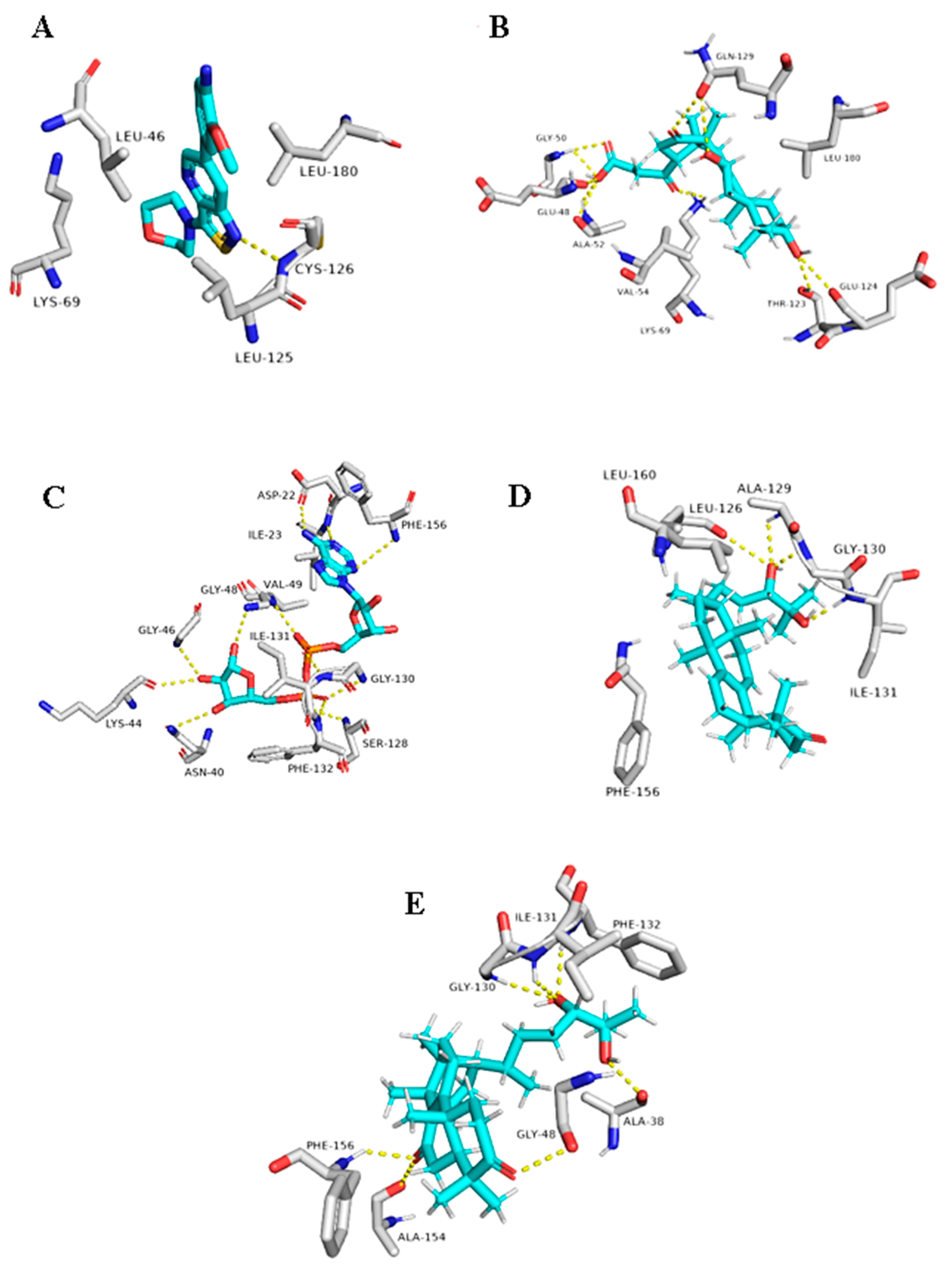
5. In Silico Investigation
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2013, 30, 237–323. [Google Scholar] [CrossRef] [PubMed]
- Salendra, L.; Lin, X.; Chen, W.; Pang, X.; Luo, X.; Long, J.; Liao, S.; Wang, J.; Zhou, X.; Liu, Y. Cytotoxicity of polyketides and steroids isolated from the sponge-associated fungus Penicillium citrinum SCSIO 41017. Nat. Prod. Res. 2019, 1478–6419, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vil, V.A.; Terent’ev, A.O.; Savidov, N.; Gloriozova, T.A.; Poroikov, V.V.; Pounina, T.A.; Dembitsky, V.M. Hydroperoxy steroids and triterpenoids derived from plant and fungi: Origin, structures, and biological activities. J. Steroid Biochem. Mol. Biol. 2019, 190, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Withering, W. An Account of the Foxglove, and Some of Its Medical Uses; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Krim, S.R.; Vivo, R.P.; Perez, J.; Inklab, M.; Tenner Jr, T.; Hodgson, J. Digoxin: Current use and approach to toxicity. Am. J. Med Sci. 2008, 336, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Russell, A. Mupirocin, fusidic acid and bacitracin: Activity, action and clinical uses of three topical antibiotics. Vet. Dermatol. 1999, 10, 225–240. [Google Scholar] [CrossRef]
- Mori, R.C.; da Silva, T.P.; Campello, R.S.; Machado, U.F. Carbenoxolone enhances peripheral insulin sensitivity and GLUT4 expression in skeletal muscle of obese rats: Potential participation of UBC9 protein. Life Sci. 2019, 229, 157–165. [Google Scholar] [CrossRef]
- Doll, R.; Hill, I.; Hutton, C. Treatment of gastric ulcer with carbenoxolone sodium and oestrogens. Gut 1965, 6, 19. [Google Scholar] [CrossRef]
- Ming, Z.; Hu, Y.; Qiu, Y.; Cao, L.; Zhang, X. Synergistic effects of β-aescin and 5-fluorouracil in human hepatocellular carcinoma SMMC-7721 cells. Phytomedicine 2010, 17, 575–580. [Google Scholar] [CrossRef]
- Shamsabadipour, S.; Ghanadian, M.; Saeedi, H.; Rahimnejad, M.R.; Mohammadi-Kamalabadi, M.; Ayatollahi, S.M.; Salimzadeh, L. Triterpenes and steroids from Euphorbia denticulata Lam. with anti-Herpes symplex virus activity. Iran. J. Pharm. Res. 2013, 12, 759. [Google Scholar]
- Zhou, W.B.; Tao, J.Y.; Xu, H.M.; Chen, K.L.; Zeng, G.Z.; Ji, C.J.; Zhang, Y.M.; Tan, N.H. Three new antiviral triterpenes from Aster tataricus. Z. Nat. B 2010, 65, 1393–1396. [Google Scholar] [CrossRef]
- Rezanka, T.; Siristova, L.; Sigler, K. Sterols and triterpenoids with antiviral activity. Anti Infect. Agents Med. Chem. 2009, 8, 193–210. [Google Scholar] [CrossRef]
- Buckingham, J. (Ed.) Dictionary of Natural Products; Chapman and Hall: London, UK, 1994. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Alhadrami, H.A.; El-Gendy, A.O.; Shamikh, Y.I.; Belbahri, L.; Hassan, H.M.; Abdelmohsen, U.R.; Rateb, M.E. Microbial natural products as potential inhibitors of SARS-CoV-2 main protease (Mpro). Microorganisms 2020, 8, 970. [Google Scholar] [CrossRef] [PubMed]
- Lill, M.A.; Danielson, M.L. Computer-aided drug design platform using PyMOL. J. Comput. Aided Mol. Des. 2011, 25, 13–19. [Google Scholar] [CrossRef]
- Akihisa, T.; Ogihara, J.; Kato, J.; Yasukawa, K.; Ukiya, M.; Yamanouchi, S.; Oishi, K. Inhibitory effects of triterpenoids and sterols on human immunodeficiency virus-1 reverse transcriptase. Lipids 2001, 36, 507–512. [Google Scholar] [CrossRef]
- Kim, S.-K.; Van Ta, Q. Chapter 17—Bioactive Sterols from Marine Resources and Their Potential Benefits for Human Health. In Advances in Food and Nutrition Research; Kim, S.-K., Ed.; Academic Press: Cambridge, MA, USA, 2012; Volume 65, pp. 261–268. [Google Scholar]
- Cong, R.; Zhang, Y.; Tian, W. A concise synthesis of the steroidal core of clathsterol. Tetrahedron Lett. 2010, 51, 3890–3892. [Google Scholar] [CrossRef]
- Quintão, N.L.M.; Rocha, L.W.; Silva, G.F.; Reichert, S.; Claudino, V.D.; Lucinda-Silva, R.M.; Malheiros, A.; Souza, M.M.D.; Bellé Bresolin, T.M.; Machado, M.D.S. Contribution of α, β-Amyrenone to the Anti-Inflammatory and Antihypersensitivity Effects of Aleurites moluccana (L.) Willd. Biomed Res. Int. 2014, 2014, 636839. [Google Scholar]
- Tanaka, S.; Uno, C.; Akimoto, M.; Tabata, M.; Honda, C.; Kamisako, W. Anti-allergic effect of bryonolic acid from Luffa cylindrica cell suspension cultures. Planta Med. 1991, 57, 527–530. [Google Scholar] [CrossRef]
- Lertphadungkit, P.; Suksiriworapong, J.; Satitpatipan, V.; Sirikantaramas, S.; Wongrakpanich, A.; Bunsupa, S. Enhanced Production of Bryonolic Acid in Trichosanthes cucumerina L. (Thai Cultivar) Cell Cultures by Elicitors and Their Biological Activities. Plants 2020, 9, 709. [Google Scholar] [CrossRef]
- Akihisa, T.; Yamamoto, K.; Tamura, T.; Kimura, Y.; Iida, T.; Nambara, T.; Chang, F.C. Triterpenoid Ketones from Lingnania chungii MoClure: Arborinone, friedelin and glutinone. Chem. Pharm. Bull. 1992, 40, 789–791. [Google Scholar] [CrossRef]
- Pengsuparp, T.; Cai, L.; Constant, H.; Fong, H.H.; Lin, L.-Z.; Kinghorn, A.D.; Pezzuto, J.M.; Cordell, G.A.; Ingolfsdöttir, K.; Wagner, H. Mechanistic evaluation of new plant-derived compounds that inhibit HIV-1 reverse transcriptase. J. Nat. Prod. 1995, 58, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Al-Rehaily, A.J.; El-Tahir, K.E.; Mossa, J.S.; Rafatullah, S. Pharmacological studies of various extracts and the major constituent, lupeol, obtained from hexane extract of Teclea nobilis in rodents. Nat. Prod. Sci. 2001, 7, 76–82. [Google Scholar]
- Fernández, A.; Alvarez, A.; García, M.D.; Sáenz, M.T. Anti-inflammatory effect of Pimenta racemosa var. ozua and isolation of the triterpene lupeol. Il Farm. 2001, 56, 335–338. [Google Scholar] [CrossRef]
- Fernández, M.A.; de las Heras, B.; Garcia, M.D.; Sáenz, M.T.; Villar, A. New insights into the mechanism of action of the anti-inflammatory triterpene lupeol. J. Pharm. Pharmacol. 2001, 53, 1533–1539. [Google Scholar] [CrossRef]
- Chaturvedi, P.K.; Bhui, K.; Shukla, Y. Lupeol: Connotations for chemoprevention. Cancer Lett. 2008, 263, 1–13. [Google Scholar] [CrossRef]
- Sudhahar, V.; Kumar, S.A.; Varalakshmi, P.; Sujatha, V. Protective effect of lupeol and lupeol linoleate in hypercholesterolemia associated renal damage. Mol. Cell. Biochem. 2008, 317, 11. [Google Scholar] [CrossRef]
- Sudhahar, V.; Veena, C.K.; Varalakshmi, P. Antiurolithic effect of lupeol and lupeol linoleate in experimental hyperoxaluria. J. Nat. Prod. 2008, 71, 1509–1512. [Google Scholar] [CrossRef]
- Siddique, H.R.; Saleem, M. Beneficial health effects of lupeol triterpene: A review of preclinical studies. Life Sci. 2011, 88, 285–293. [Google Scholar] [CrossRef]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J.; Collaboration, H.A.S. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033. [Google Scholar] [CrossRef]
- Perez, R. Antiviral activity of compounds isolated from plants. Pharm. Biol. 2003, 41, 107–157. [Google Scholar] [CrossRef]
- Khwaza, V.; Oyedeji, O.O.; Aderibigbe, B.A. Antiviral activities of oleanolic acid and its analogues. Molecules 2018, 23, 2300. [Google Scholar] [CrossRef] [PubMed]
- Dzubak, P.; Hajduch, M.; Vydra, D.; Hustova, A.; Kvasnica, M.; Biedermann, D.; Markova, L.; Urban, M.; Sarek, J. Pharmacological activities of natural triterpenoids and their therapeutic implications. Nat. Prod. Rep. 2006, 23, 394–411. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Khattab, A.R.; AboulMagd, A.M.; Hassan, H.M.; Rateb, M.E.; Zaid, H.; Abdelmohsen, U.R. Nature as a treasure trove of potential anti-SARS-CoV drug leads: A structural/mechanistic rationale. RSC Adv. 2020, 10, 19790–19802. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Sujarit, K.; Pattananandecha, T.; Saenjum, C.; Lumyong, S. Natural Bioactive Compounds from Fungi as Potential Candidates for Protease Inhibitors and Immunomodulators to Apply for Coronaviruses. Molecules 2020, 25, 1800. [Google Scholar] [CrossRef] [PubMed]
- El Dine, R.S.; El Halawany, A.M.; Ma, C.-M.; Hattori, M. Anti-HIV-1 protease activity of lanostane triterpenes from the vietnamese mushroom Ganoderma colossum. J. Nat. Prod. 2008, 71, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Sirtori, C.R. Aescin: Pharmacology, pharmacokinetics and therapeutic profile. Pharmacol. Res. 2001, 44, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Pittler, M.H.; Ernst, E. Horse chestnut seed extract for chronic venous insufficiency. Cochrane Database Syst. Rev. 2012, 1465–1858. [Google Scholar] [CrossRef]
- Domanski, D.; Zegrocka-Stendel, O.; Perzanowska, A.; Dutkiewicz, M.; Kowalewska, M.; Grabowska, I.; Maciejko, D.; Fogtman, A.; Dadlez, M.; Koziak, K. Molecular mechanism for cellular response to β-escin and its therapeutic implications. PLoS ONE 2016, 11, e0164365. [Google Scholar] [CrossRef]
- Xin, W.; Zhang, L.; Sun, F.; Jiang, N.; Fan, H.; Wang, T.; Li, Z.; He, J.; Fu, F. Escin exerts synergistic anti-inflammatory effects with low doses of glucocorticoids in vivo and in vitro. Phytomedicine 2011, 18, 272–277. [Google Scholar] [CrossRef]
- Subramanya, S.B.; Venkataraman, B.; Meeran, M.F.N.; Goyal, S.N.; Patil, C.R.; Ojha, S. Therapeutic potential of plants and plant derived phytochemicals against acetaminophen-induced liver injury. Int. J. Mol. Sci. 2018, 19, 3776. [Google Scholar] [CrossRef]
- Kashiwada, Y.; Wang, H.-K.; Nagao, T.; Kitanaka, S.; Yasuda, I.; Fujioka, T.; Yamagishi, T.; Cosentino, L.M.; Kozuka, M.; Okabe, H. Anti-AIDS agents. 30. Anti-HIV activity of oleanolic acid, pomolic acid, and structurally related triterpenoids. J. Nat. Prod. 1998, 61, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-F.; Zhang, S.-X.; Wang, H.-K.; Zhang, S.-Y.; Sun, Q.-Z.; Cosentino, L.M.; Lee, K.-H. Novel anti-HIV lancilactone C and related triterpenes from Kadsura lancilimba. J. Nat. Prod. 1999, 62, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Oprean, C.; Mioc, M.; Csányi, E.; Ambrus, R.; Bojin, F.; Tatu, C.; Cristea, M.; Ivan, A.; Danciu, C.; Dehelean, C. Improvement of ursolic and oleanolic acids’ antitumor activity by complexation with hydrophilic cyclodextrins. Biomed. Pharmacother. 2016, 83, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zeng, H.; Wang, Y.; Fan, X.; Xu, C.; Deng, R.; Zhou, X.; Bi, H.; Huang, M. Low dose of oleanolic acid protects against lithocholic acid–induced cholestasis in mice: Potential involvement of nuclear factor-E2-related factor 2-mediated upregulation of multidrug resistance-associated proteins. Drug Metab. Dispos. 2014, 42, 844–852. [Google Scholar] [CrossRef]
- Jesus, J.A.; Lago, J.H.G.; Laurenti, M.D.; Yamamoto, E.S.; Passero, L.F.D. Antimicrobial activity of oleanolic and ursolic acids: An update. Evid. Based Complement. Altern. Med. 2015, 2015, 620472. [Google Scholar] [CrossRef]
- Zhu, Y.-M.; Shen, J.-K.; Wang, H.-K.; Cosentino, L.M.; Lee, K.-H. Synthesis and anti-HIV activity of oleanolic acid derivatives. Bioorg. Med. Chem. Lett. 2001, 11, 3115–3118. [Google Scholar] [CrossRef]
- Li, H.-Y.; Sun, N.-J.; Kashiwada, Y.; Sun, L.; Snider, J.V.; Cosentino, L.M.; Lee, K.-H. Anti-AIDS agents, 9. Suberosol, a new C31 lanostane-type triterpene and anti-HIV principle from Polyalthia suberosa. J. Nat. Prod. 1993, 56, 1130–1133. [Google Scholar] [CrossRef]
- Bifulco, G.; Bruno, I.; Minale, L.; Riccio, R. Novel HIV-inhibitory halistanol sulfates F-H from a marine sponge, Pseudoaxinissa digitata. J. Nat. Prod. 1994, 57, 164–167. [Google Scholar] [CrossRef]
- Koehn, F.E.; Gunasekera, M.; Cross, S.S. New antiviral sterol disulfate ortho esters from the marine sponge Petrosia weinbergi. J. Org. Chem. 1991, 56, 1322–1325. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.; Gao, W.; Wang, H. Study on chemical composition, anti-inflammatory and anti-microbial activities of extracts from Chinese pear fruit (Pyrus bretschneideri Rehd.). Food Chem. Toxicol. 2012, 50, 3673–3679. [Google Scholar] [CrossRef]
- Moghaddam, M.G.; Faujan Bin, H.; Ahmad, F.H.; Kermani, A.S. Biological Activity of Betulinic Acid: A Review. Pharmacol. Pharm. 2012, 3, 119–123. [Google Scholar] [CrossRef]
- Yu, D.; Wild, C.T.; Martin, D.E.; Morris-Natschke, S.L.; Chen, C.-H.; Allaway, G.P.; Lee, K.-H. The discovery of a class of novel HIV-1 maturation inhibitors and their potential in the therapy of HIV. Expert Opin. Investig. Drugs 2005, 14, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Paduch, R.; Kandefer-Szerszen, M. Antitumor and antiviral activity of pentacyclic triterpenes. Mini Rev. Org. Chem. 2014, 11, 262–268. [Google Scholar] [CrossRef]
- Soler, F.; Poujade, C.; Evers, M.; Carry, J.-C.; Hénin, Y.; Bousseau, A.; Huet, T.; Pauwels, R.; De Clercq, E.; Mayaux, J.-F. Betulinic acid derivatives: A new class of specific inhibitors of human immunodeficiency virus type 1 entry. J. Med. Chem. 1996, 39, 1069–1083. [Google Scholar] [CrossRef]
- Qian, K.; Morris-Natschke, S.L.; Lee, K.H. HIV entry inhibitors and their potential in HIV therapy. Med. Res. Rev. 2009, 29, 369–393. [Google Scholar] [CrossRef]
- Pohjala, L.; Alakurtti, S.; Ahola, T.; Yli-Kauhaluoma, J.; Tammela, P. Betulin-derived compounds as inhibitors of alphavirus replication. J. Nat. Prod. 2009, 72, 1917–1926. [Google Scholar] [CrossRef]
- Pavlova, N.; Savinova, O.; Nikolaeva, S.; Boreko, E.; Flekhter, O. Antiviral activity of betulin, betulinic and betulonic acids against some enveloped and non-enveloped viruses. Fitoterapia 2003, 74, 489–492. [Google Scholar] [CrossRef]
- Flekhter, O.B.; Karachurina, L.T.; Nigmatullina, L.R.; Sapozhnikova, T.A.; Baltina, L.A.; Zarudii, F.S.; Galin, F.Z.; Spirikhin, L.V.; Tolstikov, G.A.; Plyasunova, O.A. Synthesis and pharmacological activity of betulin dinicotinate. Russ. J. Bioorg. Chem. 2002, 28, 494–500. [Google Scholar] [CrossRef]
- Baltina, L.A.; Flekhter, O.B.; Nigmatullina, L.R.; Boreko, E.I.; Pavlova, N.I.; Nikolaeve, S.N.; Savinova, O.V.; Tolstikov, G.A. Lupane triterpenes and derivatives with antiviral activity. Bioorg. Med. Chem. Lett. 2003, 13, 3549–3552. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Nguyen, A.H.; Kumar, A.P.; Tan, B.K.; Sethi, G. Targeted inhibition of tumor proliferation, survival, and metastasis by pentacyclic triterpenoids: Potential role in prevention and therapy of cancer. Cancer Lett. 2012, 320, 158–170. [Google Scholar] [CrossRef]
- Pompei, R.; Flore, O.; Marccialis, M.A.; Pani, A.; Loddo, B. Glycyrrhizic acid inhibits virus growth and inactivates virus particles. Nature 1979, 281, 689–690. [Google Scholar] [CrossRef] [PubMed]
- Baba, M.; Shigeta, S. Antiviral activity of glycyrrhizin against varicella-zoster virus in vitro. Antivir. Res. 1987, 7, 99–107. [Google Scholar] [CrossRef]
- Crance, J.M.; Scaramozzino, N.; Jouan, A.; Garin, D. Interferon, ribavirin, 6-azauridine and glycyrrhizin: Antiviral compounds active against pathogenic flaviviruses. Antivir. Res. 2003, 58, 73–79. [Google Scholar] [CrossRef]
- Lin, J.-C.; Cherng, J.-M.; Hung, M.-S.; Baltina, L.A.; Baltina, L.; Kondratenko, R. Inhibitory effects of some derivatives of glycyrrhizic acid against Epstein-Barr virus infection: Structure–activity relationships. Antivir. Res. 2008, 79, 6–11. [Google Scholar] [CrossRef]
- Takahara, T.; Watanabe, A.; Shiraki, K. Effects of glycyrrhizin on hepatitis B surface antigen: A biochemical and morphological study. J. Hepatol. 1994, 21, 601–609. [Google Scholar] [CrossRef]
- Sasaki, H.; Takei, M.; Kobayashi, M.; Pollard, R.B.; Suzuki, F. Effect of glycyrrhizin, an active component of licorice roots, on HIV replication in cultures of peripheral blood mononuclear cells from HIV-seropositive patients. Pathobiology 2002, 70, 229–236. [Google Scholar] [CrossRef]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H. Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef]
- Wolkerstorfer, A.; Kurz, H.; Bachhofner, N.; Szolar, O.H. Glycyrrhizin inhibits influenza A virus uptake into the cell. Antivir. Res. 2009, 83, 171–178. [Google Scholar] [CrossRef]
- Sui, X.; Yin, J.; Ren, X. Antiviral effect of diammonium glycyrrhizinate and lithium chloride on cell infection by pseudorabies herpesvirus. Antivir. Res. 2010, 85, 346–353. [Google Scholar] [CrossRef]
- Hardy, M.E.; Hendricks, J.M.; Paulson, J.M.; Faunce, N.R. 18 β-glycyrrhetinic acid inhibits rotavirus replication in culture. Virol. J. 2012, 9, 1–7. [Google Scholar] [CrossRef]
- Kang, H.; Lieberman, P.M. Mechanism of glycyrrhizic acid inhibition of Kaposi’s sarcoma-associated herpesvirus: Disruption of CTCF-cohesin-mediated RNA polymerase II pausing and sister chromatid cohesion. J. Virol. 2011, 85, 11159–11169. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, U.A.; Masoud, M.S.; Nawaz, Z.; Riazuddin, S. Glycyrrhizin as antiviral agent against Hepatitis C Virus. J. Transl. Med. 2011, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chama, M.A.; Dziwornu, G.A.; Waibel, R.; Osei-Safo, D.; Addae-Mensah, I.; Otchere, J.; Wilson, M. Isolation, characterization, and anthelminthic activities of a novel dichapetalin and other constituents of Dichapetalum filicaule. Pharm. Biol. 2016, 54, 1179–1188. [Google Scholar] [PubMed]
- Rojano, B.; Saez, J.; Schinella, G.; Quijano, J.; Vélez, E.; Gil, A.; Notario, R. Experimental and theoretical determination of the antioxidant properties of isoespintanol (2-Isopropyl-3, 6-dimethoxy-5-methylphenol). J. Mol. Struct. 2008, 877, 1–6. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Park, S.-J.; Kim, Y.M.; Lee, J.-Y.; Seo, W.D.; Chang, J.S.; Park, K.H.; Rho, M.-C.; Lee, W.S. SARS-CoV 3CLpro inhibitory effects of quinone-methide triterpenes from Tripterygium regelii. Bioorg. Med. Chem. Lett. 2010, 20, 1873–1876. [Google Scholar] [CrossRef]
- Nakanishi, K.; Takahashi, Y.; Budzikiewicz, H. Pristimerin. Spectroscopic Properties of the Dienone—Phenol-Type Rearrangement Products and Other Derivatives1. J. Org. Chem. 1965, 30, 1729–1734. [Google Scholar] [CrossRef]
- Monache, F.; Bettólo, G.; de Lima, O.; D’Albuquerque, I.; de Barros, C.J. The structure of tingenone, a quinonoid triterpene related to pristimerin. J. Chem. Soc. Perkin Trans. 1973, 22, 2725. [Google Scholar] [CrossRef]
- Itokawa, H.; Shirota, O.; Ikuta, H.; Morita, H.; Takeya, K.; Iitaka, Y. Triterpenes fromMaytenus ilicifolia. Phytochemistry 1991, 30, 3713–3716. [Google Scholar] [CrossRef]
- Pomponi, M.; Delle Monache, F. “Rearrangements of Tingenone.” IV. Researches on Quinonoid Triterpenes; Real Sociedad Española de Química: Madrid, Spain, 1974. [Google Scholar]
- Sotanaphun, U.; Lipipun, V.; Suttisri, R.; Bavovada, R. Antimicrobial activity and stability of tingenone derivatives. Planta Med. 1999, 65, 450–452. [Google Scholar] [CrossRef]
- Sotanaphun, U.; Lipipun, V.; Yaipakdee, P.; Bavovada, R. New acidic-rearranged compounds from tingenone derivatives and their biological activity. Pharm. Biol. 2005, 43, 39–46. [Google Scholar] [CrossRef][Green Version]
- Ito, M.; Nakashima, H.; Baba, M.; Pauwels, R.; De Clercq, E.; Shigeta, S.; Yamamoto, N. Inhibitory effect of glycyrrhizin on the in vitro infectivity and cytopathic activity of the human immunodeficiency virus [HIV (HTLV-III/LAV)]. Antivir. Res. 1987, 7, 127–137. [Google Scholar] [CrossRef]
- Ushio, Y.; Abe, H. Inactivation of measles virus and herpes simplex virus by saikosaponin d. Planta Med. 1992, 58, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Rao, G.S.; Sinsheimer, J.E.; Cochran, K.W. Antiviral activity of triterpenoid saponins containing acylated β-amyrin aglycones. J. Pharm. Sci. 1974, 63, 471–473. [Google Scholar] [PubMed]
- Amoros, M.; Fauconnier, B.; Girre, R. In vitro antiviral activity of a saponin from Anagallis arvensis, Primulaceae, against herpes simplex virus and poliovirus. Antivir. Res. 1987, 8, 13–25. [Google Scholar] [CrossRef]
- Chaudhuri, S.K.; Fullas, F.; Brown, D.M.; Wani, M.C.; Wall, M.E.; Cai, L.; Mar, W.; Lee, S.K.; Luo, Y.; Zaw, K. Isolation and structural elucidation of pentacyclic triterpenoids from Maprounea africana. J. Nat. Prod. 1995, 58, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rut, W.; Lv, Z.; Zmudzinski, M.; Patchett, S.; Nayak, D.; Snipas, S.J.; El Oualid, F.; Bekes, M.; Huang, T.T.; Drag, M. Activity profiling and structures of inhibitor-bound SARS-CoV-2-PLpro protease provides a framework for anti-COVID-19 drug design. bioRxiv 2020, 6, eabd4596. [Google Scholar]
- Bekerman, E.; Neveu, G.; Shulla, A.; Brannan, J.; Pu, S.-Y.; Wang, S.; Xiao, F.; Barouch-Bentov, R.; Bakken, R.R.; Mateo, R. Anticancer kinase inhibitors impair intracellular viral trafficking and exert broad-spectrum antiviral effects. J. Clin. Investig. 2017, 127, 1338–1352. [Google Scholar] [CrossRef]
- Gil, C.; Ginex, T.; Maestro, I.; Nozal, V.; Barrado-Gil, L.; Cuesta-Geijo, M.A.; Urquiza, J.; Ramírez, D.; Alonso, C.; Campillo, N.E. COVID-19: Drug targets and potential treatments. J. Med. Chem. 2020, 63, 12359–12386. [Google Scholar] [CrossRef]
- Cantini, F.; Banci, L.; Altincekic, N.; Bains, J.; Dhamotharan, K.; Fuks, C.; Fürtig, B.; Gande, S.; Hargittay, B.; Hengesbach, M. 1 H, 13 C, and 15 N backbone chemical shift assignments of the apo and the ADP-ribose bound forms of the macrodomain of SARS-CoV-2 non-structural protein 3b. Biomol. NMR Assign. 2020, 14, 339–346. [Google Scholar] [CrossRef]
- Michalska, K.; Kim, Y.; Jedrzejczak, R.; Maltseva, N.I.; Stols, L.; Endres, M.; Joachimiak, A. Crystal structures of SARS-CoV-2 ADP-ribose phosphatase (ADRP): From the apo form to ligand complexes. bioRxiv 2020, 7, 814–824. [Google Scholar]
- Hattori, M.; Ma, C.-M.; Wei, Y.; Dine, R.; Sato, N. Survey of anti-HIV and anti-HCV compounds from Natural sources. Can Chem. Trans. 2013, 1, 116–140. [Google Scholar]
- Min, B.-S.; Nakamura, N.; Miyashiro, H.; BAE, K.-W.; Hattori, M. Triterpenes from the spores of Ganoderma lucidum and their inhibitory activity against HIV-1 protease. Chem. Pharm. Bull. 1998, 46, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ARP (6W02) | Mpro (6LU7) | PLpro (6WXR) | AAK1 (4WSQ) | GAK (4Y8D) | Cathepsin L(2YJC) | Furin (6EQX) |
|---|---|---|---|---|---|---|---|
| 1ß-Hydroxyaleuritolic acid 3-p-hydroxy-benzoate (11) | >−4 | −8.5 | −8 | −9.1 | >-4 | −8.1 | −8.7 |
| 20(21)-dehydrolucidenic acid (12) | >−4 | >−4 | >−4 | >−4 | −8.1 | >−4 | >−4 |
| Ganodermanondiol (13) | −8.3 | >−4 | >−4 | >−4 | >−4 | >−4 | >-4 |
| Ganoderiol F (14) | −8 | >−4 | >−4 | >−4 | >−4 | >−4 | −8.3 |
| Lucidumol A (15) | −9.6 | >−4 | >−4 | >−4 | >−4 | >−4 | |
| Suberosol (16) | >−4 | >−4 | >−4 | −9 | −8.2 | >−4 | >−4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hisham Shady, N.; Youssif, K.A.; Sayed, A.M.; Belbahri, L.; Oszako, T.; Hassan, H.M.; Abdelmohsen, U.R. Sterols and Triterpenes: Antiviral Potential Supported by In-Silico Analysis. Plants 2021, 10, 41. https://doi.org/10.3390/plants10010041
Hisham Shady N, Youssif KA, Sayed AM, Belbahri L, Oszako T, Hassan HM, Abdelmohsen UR. Sterols and Triterpenes: Antiviral Potential Supported by In-Silico Analysis. Plants. 2021; 10(1):41. https://doi.org/10.3390/plants10010041
Chicago/Turabian StyleHisham Shady, Nourhan, Khayrya A. Youssif, Ahmed M. Sayed, Lassaad Belbahri, Tomasz Oszako, Hossam M. Hassan, and Usama Ramadan Abdelmohsen. 2021. "Sterols and Triterpenes: Antiviral Potential Supported by In-Silico Analysis" Plants 10, no. 1: 41. https://doi.org/10.3390/plants10010041
APA StyleHisham Shady, N., Youssif, K. A., Sayed, A. M., Belbahri, L., Oszako, T., Hassan, H. M., & Abdelmohsen, U. R. (2021). Sterols and Triterpenes: Antiviral Potential Supported by In-Silico Analysis. Plants, 10(1), 41. https://doi.org/10.3390/plants10010041