
The Organizer and Its Signaling in Embryonic Development



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
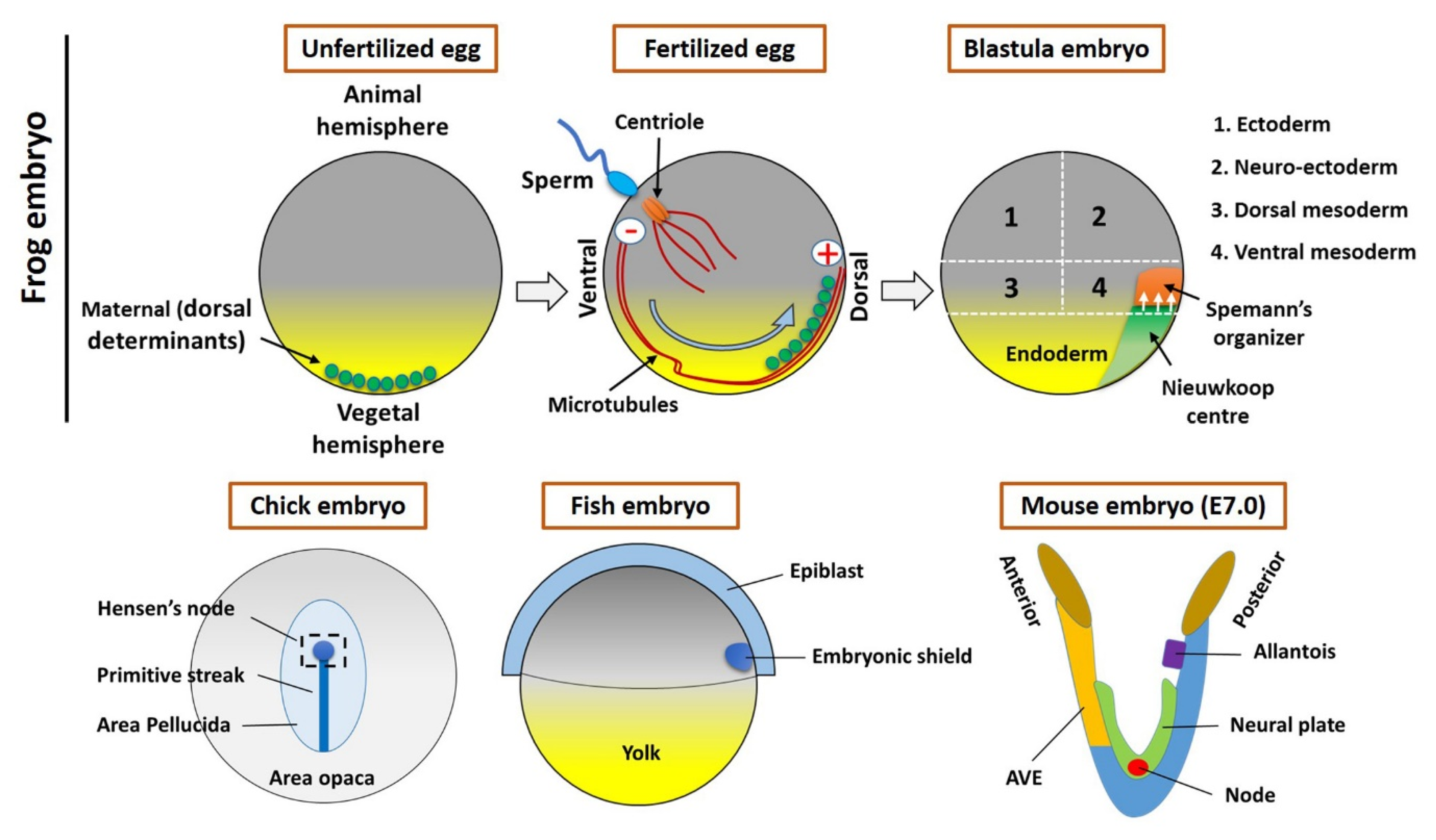
2. The Spemann’s Organizer and Homologous Tissue in Vertebrates
2.1. The Maternal Determinants Establish the Organizer
2.2. The Homologous Structure of Organizer in Other Vertebrates
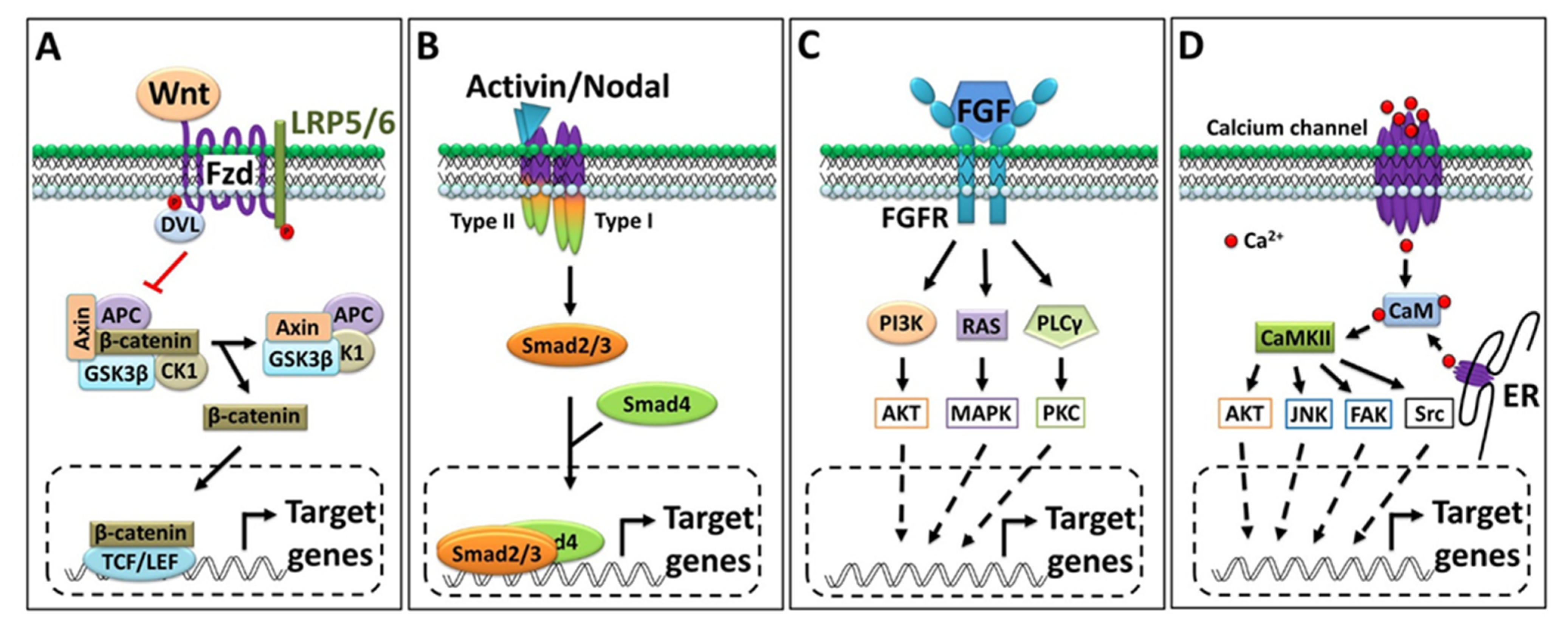
3. Overview of Major Signaling Pathways and Targets Involved in Organizer-Induced Embryonic Development
3.1. Wnt/β-Catenin Signaling
3.2. Activin/Inhibin and Nodal Signaling
3.3. FGF Signaling
3.4. Calcium Signaling
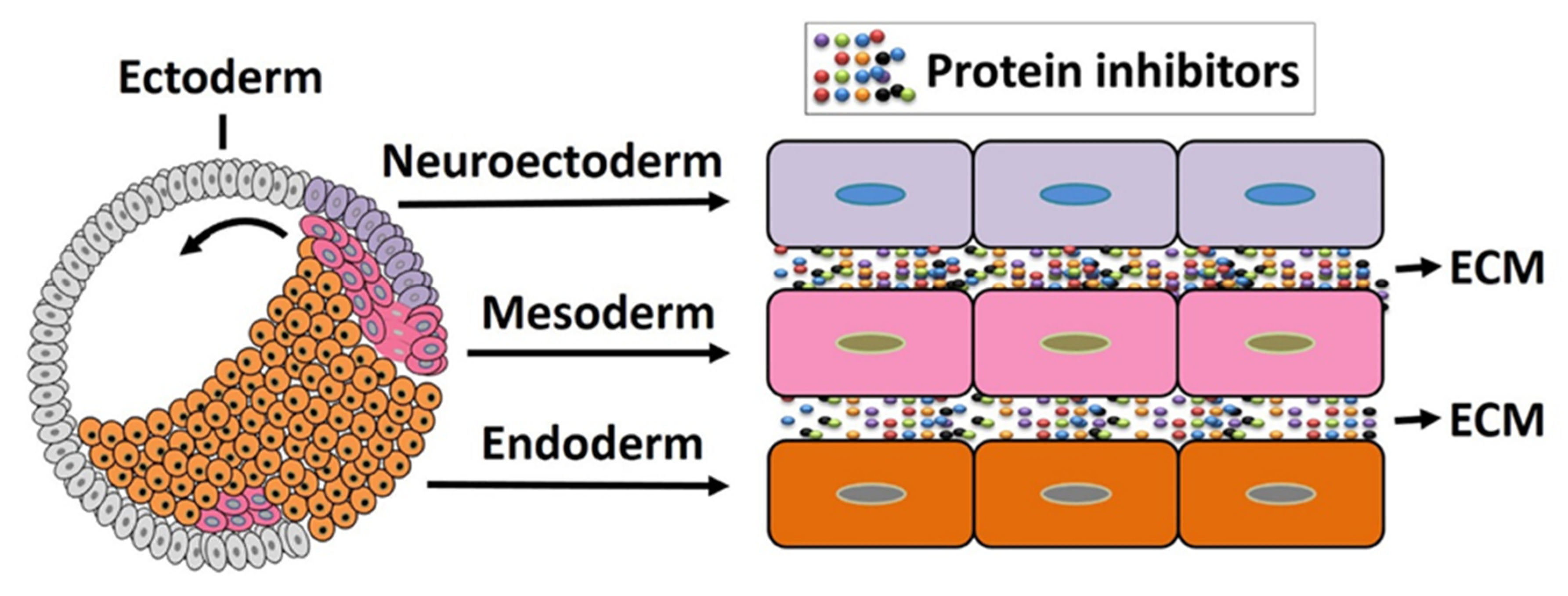
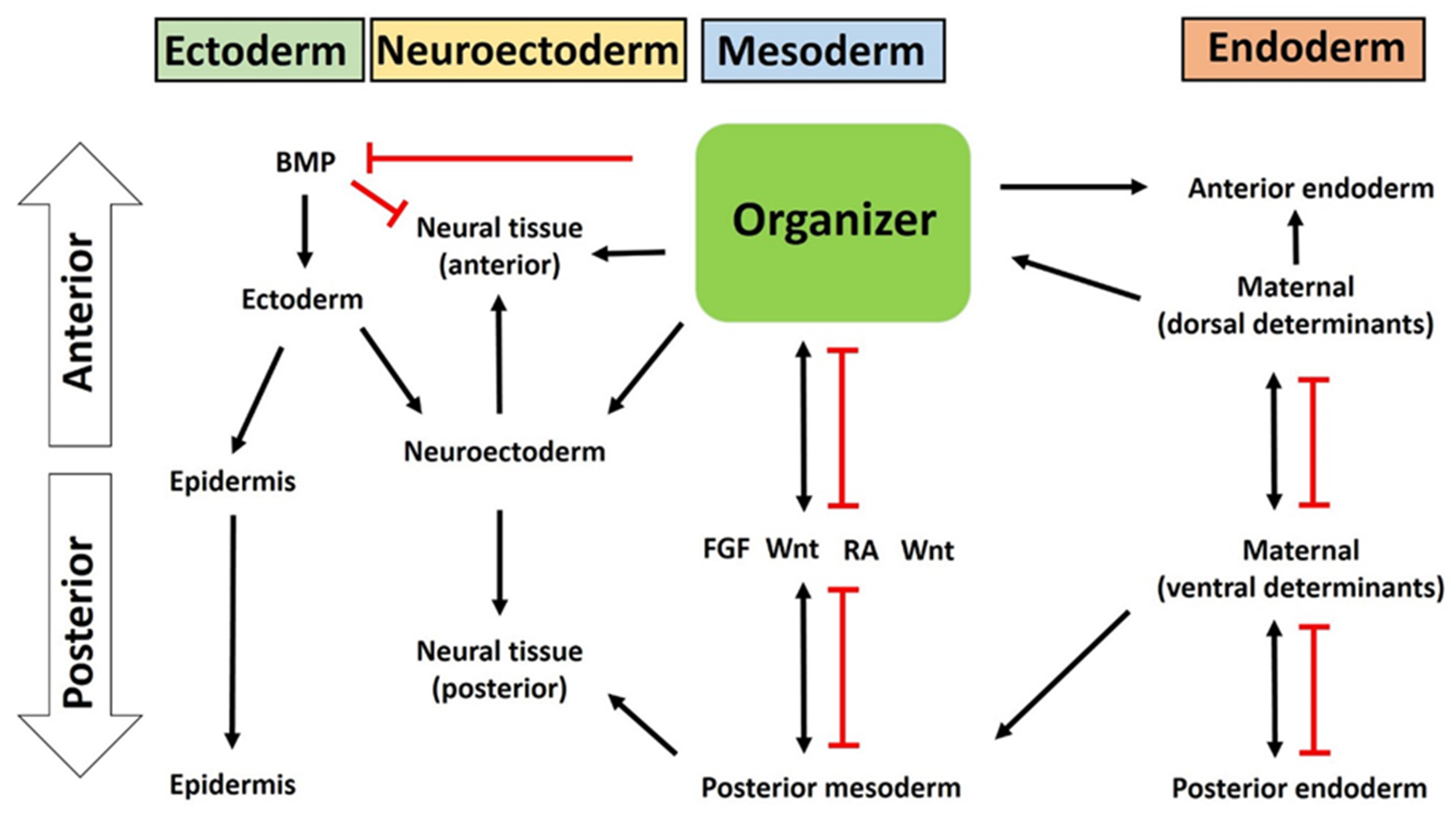
4. The Organizer as an Organizing Center of Vertebrate Embryonic Patterning
4.1. Organizer in Mesoderm Formation and Patterning
4.2. The Organizer in Ectoderm Patterning
4.3. The Organizer Induces Neural Induction and Neuroectoderm Formation
4.4. The Organizer Is Involved in Endoderm Patterning
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spemann, H.; Mangold, H. über Induktion von Embryonalanlagen durch Implantation artfremder Organisatoren. Arch. Für Mikrosk. Anat. Und Entwickl. 1924, 100, 599–638. [Google Scholar]
- De Robertis, E.M. Spemann’s organizer and self-regulation in amphibian embryos. Nat. Rev. Mol. Cell Biol. 2006, 7, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, P.; Kodjabachian, L. The vertebrate organizer: Structure and molecules. Trends Genet. 1996, 12, 525–531. [Google Scholar] [CrossRef]
- Anderson, C.; Khan, M.A.F.; Wong, F.; Solovieva, T.; Oliveira, N.M.M.; Baldock, R.A.; Tickle, C.; Burt, D.W.; Stern, C.D. A strategy to discover new organizers identifies a putative heart organizer. Nat. Commun. 2016, 7, 12656. [Google Scholar] [CrossRef]
- Thisse, B.; Thisse, C. Formation of the vertebrate embryo: Moving beyond the Spemann organizer. Semin. Cell Dev. Biol. 2015, 42, 94–102. [Google Scholar] [CrossRef]
- Harland, R.; Gerhart, J. Formation and function of Spemann’s organizer. Annu. Rev. Cell Dev. Biol. 1997, 13, 611–667. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, Y.; De Robertis, E.M. Embryonic regeneration by relocalization of the Spemann organizer during twinning in Xenopus. Proc. Natl. Acad. Sci. USA 2018, 115, E4815–E4822. [Google Scholar] [CrossRef]
- Garcia-Fernandez, J.; D’ Aniello, S.; Escriva, H. Organizing chordates with an organizer. Bioessays 2007, 29, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Xanthos, J.B.; Kofron, M.; Tao, Q.; Schaible, K.; Wylie, C.; Heasman, J. The roles of three signaling pathways in the formation and function of the Spemann Organizer. Development 2002, 129, 4027–4043. [Google Scholar] [CrossRef]
- De Robertis, E.M.; Larrain, J.; Oelgeschlager, M.; Wessely, O. The establishment of Spemann’s organizer and patterning of the vertebrate embryo. Nat. Rev. Genet. 2000, 1, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Carron, C.; Shi, D.L. Specification of anteroposterior axis by combinatorial signaling during Xenopus development. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 150–168. [Google Scholar] [CrossRef]
- Martinez Arias, A.; Steventon, B. On the nature and function of organizers. Development 2018, 145, dev159525. [Google Scholar] [CrossRef]
- Rodriguez, T.A.; Srinivas, S.; Clements, M.P.; Smith, J.C.; Beddington, R.S. Induction and migration of the anterior visceral endoderm is regulated by the extra-embryonic ectoderm. Development 2005, 132, 2513–2520. [Google Scholar] [CrossRef] [PubMed]
- Leikola, A. Hensen’s node—the ‘organizer’ of the amniote embryo. Experientia 1976, 32, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Niehrs, C. Regionally specific induction by the Spemann-Mangold organizer. Nat. Rev. Genet. 2004, 5, 425–434. [Google Scholar] [CrossRef]
- Ding, Y.; Ploper, D.; Sosa, E.A.; Colozza, G.; Moriyama, Y.; Benitez, M.D.; Zhang, K.; Merkurjev, D.; De Robertis, E.M. Spemann organizer transcriptome induction by early beta-catenin, Wnt, Nodal, and Siamois signals in Xenopus laevis. Proc. Natl. Acad. Sci. USA 2017, 114, E3081–E3090. [Google Scholar] [CrossRef] [PubMed]
- Pierce, S.B.; Kimelman, D. Regulation of Spemann organizer formation by the intracellular kinase Xgsk-3. Development 1995, 121, 755–765. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.S.; Kim, M.; Glinka, A.; Reinhard, C.; Niehrs, C. The tumor suppressor PTPRK promotes ZNRF3 internalization and is required for Wnt inhibition in the Spemann organizer. Elife 2020, 9, e51248. [Google Scholar] [CrossRef] [PubMed]
- Schohl, A.; Fagotto, F. A role for maternal beta-catenin in early mesoderm induction in Xenopus. EMBO J. 2003, 22, 3303–3313. [Google Scholar] [CrossRef] [PubMed]
- Wessely, O.; Kim, J.I.; Geissert, D.; Tran, U.; De Robertis, E.M. Analysis of Spemann organizer formation in Xenopus embryos by cDNA macroarrays. Dev. Biol. 2004, 269, 552–566. [Google Scholar] [CrossRef] [PubMed]
- Gritsman, K.; Talbot, W.S.; Schier, A.F. Nodal signaling patterns the organizer. Development 2000, 127, 921–932. [Google Scholar] [CrossRef]
- Nieto, M.A. Reorganizing the organizer 75 years on. Cell 1999, 98, 417–425. [Google Scholar] [CrossRef]
- Sudou, N.; Yamamoto, S.; Ogino, H.; Taira, M. Dynamic in vivo binding of transcription factors to cis-regulatory modules of cer and gsc in the stepwise formation of the Spemann-Mangold organizer. Development 2012, 139, 1651–1661. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Yokota, C.; Takano, K.; Tanegashima, K.; Onuma, Y.; Goto, J.; Asashima, M. Two novel nodal-related genes initiate early inductive events in Xenopus Nieuwkoop center. Development 2000, 127, 5319–5329. [Google Scholar] [CrossRef] [PubMed]
- Agius, E.; Oelgeschlager, M.; Wessely, O.; Kemp, C.; De Robertis, E.M. Endodermal Nodal-related signals and mesoderm induction in Xenopus. Development 2000, 127, 1173–1183. [Google Scholar] [CrossRef]
- Wittler, L.; Kessel, M. The acquisition of neural fate in the chick. Mech. Dev. 2004, 121, 1031–1042. [Google Scholar] [CrossRef]
- Shih, J.; Fraser, S.E. Characterizing the zebrafish organizer: Microsurgical analysis at the early-shield stage. Development 1996, 122, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Nitsche, F.; Bazzi, H. Visualizing the Node and Notochordal Plate In Gastrulating Mouse Embryos Using Scanning Electron Microscopy and Whole Mount Immunofluorescence. J. Vis. Exp. 2018, 9, e58321. [Google Scholar] [CrossRef]
- Tam, P.P.; Behringer, R.R. Mouse gastrulation: The formation of a mammalian body plan. Mech. Dev. 1997, 68, 3–25. [Google Scholar] [CrossRef]
- Satoh, N.; Tagawa, K.; Takahashi, H. How was the notochord born? Evol. Dev. 2012, 14, 56–75. [Google Scholar] [CrossRef]
- Agathon, A.; Thisse, C.; Thisse, B. The molecular nature of the zebrafish tail organizer. Nature 2003, 424, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Stuhlmiller, T.J.; Garcia-Castro, M.I. Current perspectives of the signaling pathways directing neural crest induction. Cell. Mol. Life Sci. 2012, 69, 3715–3737. [Google Scholar] [CrossRef]
- Stemple, D.L. Structure and function of the notochord: An essential organ for chordate development. Development 2005, 132, 2503–2512. [Google Scholar] [CrossRef] [PubMed]
- Darras, S.; Nishida, H. The BMP signaling pathway is required together with the FGF pathway for notochord induction in the ascidian embryo. Development 2001, 128, 2629–2638. [Google Scholar] [CrossRef]
- Kirsch, N.; Chang, L.S.; Koch, S.; Glinka, A.; Dolde, C.; Colozza, G.; Benitez, M.D.J.; De Robertis, E.M.; Niehrs, C. Angiopoietin-like 4 Is a Wnt Signaling Antagonist that Promotes LRP6 Turnover. Dev. Cell 2017, 43, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Langdon, Y.G.; Mullins, M.C. Maternal and zygotic control of zebrafish dorsoventral axial patterning. Annu. Rev. Genet. 2011, 45, 357–377. [Google Scholar] [CrossRef]
- De Robertis, E.M. Spemann’s organizer and the self-regulation of embryonic fields. Mech. Dev. 2009, 126, 925–941. [Google Scholar] [CrossRef]
- Joubin, K.; Stern, C.D. Molecular interactions continuously define the organizer during the cell movements of gastrulation. Cell 1999, 98, 559–571. [Google Scholar] [CrossRef]
- Edwards, N.A.; Zorn, A.M. Modeling endoderm development and disease in Xenopus. Curr. Top. Dev. Biol. 2021, 145, 61–90. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.H.; Brazil, D.P. Bone morphogenetic proteins and their antagonists: Current and emerging clinical uses. Br. J. Pharm. 2014, 171, 3620–3632. [Google Scholar] [CrossRef]
- Zhang, X.; Abreu, J.G.; Yokota, C.; MacDonald, B.T.; Singh, S.; Coburn, K.L.; Cheong, S.M.; Zhang, M.M.; Ye, Q.Z.; Hang, H.C.; et al. Tiki1 is required for head formation via Wnt cleavage-oxidation and inactivation. Cell 2012, 149, 1565–1577. [Google Scholar] [CrossRef] [PubMed]
- De Robertis, E.M.; Kuroda, H. Dorsal-ventral patterning and neural induction in Xenopus embryos. Annu. Rev. Cell. Dev. Biol. 2004, 20, 285–308. [Google Scholar] [CrossRef]
- Cruciat, C.M.; Niehrs, C. Secreted and transmembrane wnt inhibitors and activators. Cold Spring Harb. Perspect. Biol. 2013, 5, a015081. [Google Scholar] [CrossRef]
- Plouhinec, J.L.; Zakin, L.; Moriyama, Y.; De Robertis, E.M. Chordin forms a self-organizing morphogen gradient in the extracellular space between ectoderm and mesoderm in the Xenopus embryo. Proc. Natl. Acad. Sci. USA 2013, 110, 20372–20379. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; He, X. Wnt/beta-catenin signaling: New (and old) players and new insights. Curr. Opin. Cell Biol. 2008, 20, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Kozmikova, I.; Kozmik, Z. Wnt/beta-catenin signaling is an evolutionarily conserved determinant of chordate dorsal organizer. Elife 2020, 9, e56817. [Google Scholar] [CrossRef] [PubMed]
- Schohl, A.; Fagotto, F. Beta-catenin, MAPK and Smad signaling during early Xenopus development. Development 2002, 129, 37–52. [Google Scholar] [CrossRef]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; Hallagan, S.E.; McGrew, L.L.; Miller, J.R.; Moon, R.T. The maternal Xenopus beta-catenin signaling pathway, activated by frizzled homologs, induces goosecoid in a cell non-autonomous manner. Dev. Growth Differ. 2000, 42, 347–357. [Google Scholar] [CrossRef]
- Kjolby, R.A.S.; Harland, R.M. Genome-wide identification of Wnt/beta-catenin transcriptional targets during Xenopus gastrulation. Dev. Biol. 2017, 426, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Namwanje, M.; Brown, C.W. Activins and Inhibins: Roles in Development, Physiology, and Disease. Cold Spring Harb. Perspect. Biol. 2016, 8, a021881. [Google Scholar] [CrossRef] [PubMed]
- Schier, A.F. Nodal signaling in vertebrate development. Annu. Rev. Cell Dev. Biol. 2003, 19, 589–621. [Google Scholar] [CrossRef] [PubMed]
- Whitman, M. Nodal signaling in early vertebrate embryos: Themes and variations. Dev. Cell 2001, 1, 605–617. [Google Scholar] [CrossRef]
- Pauklin, S.; Vallier, L. Activin/Nodal signalling in stem cells. Development 2015, 142, 607–619. [Google Scholar] [CrossRef]
- Sebald, W.; Mueller, T.D. The interaction of BMP-7 and ActRII implicates a new mode of receptor assembly. Trends Biochem. Sci. 2003, 28, 518–521. [Google Scholar] [CrossRef]
- Kumar, V.; Umair, Z.; Kumar, S.; Lee, U.; Kim, J. Smad2 and Smad3 differentially modulate chordin transcription via direct binding on the distal elements in gastrula Xenopus embryos. Biochem. Biophys. Res. Commun. 2021, 559, 168–175. [Google Scholar] [CrossRef]
- Umair, Z.; Kumar, S.; Rafiq, K.; Kumar, V.; Reman, Z.U.; Lee, S.H.; Kim, S.; Lee, J.Y.; Lee, U.; Kim, J. Dusp1 modulates activin/smad2 mediated germ layer specification via FGF signal inhibition in Xenopus embryos. Anim. Cells Syst. 2020, 24, 359–370. [Google Scholar] [CrossRef]
- Papanayotou, C.; Collignon, J. Activin/Nodal signalling before implantation: Setting the stage for embryo patterning. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130539. [Google Scholar] [CrossRef] [PubMed]
- Timmer, J.; Chesnutt, C.; Niswander, L. The activin signaling pathway promotes differentiation of dI3 interneurons in the spinal neural tube. Dev. Biol. 2005, 285, 1–10. [Google Scholar] [CrossRef] [PubMed]
- McKendry, R.; Harland, R.M.; Stachel, S.E. Activin-induced factors maintain goosecoid transcription through a paired homeodomain binding site. Dev. Biol. 1998, 204, 172–186. [Google Scholar] [CrossRef]
- Kumar, V.; Goutam, R.S.; Park, S.; Lee, U.; Kim, J. Functional Roles of FGF Signaling in Early Development of Vertebrate Embryos. Cells 2021, 10, 2148. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, R.B.; Harland, R.M. The role of FGF signaling in the establishment and maintenance of mesodermal gene expression in Xenopus. Dev. Dyn. 2008, 237, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, H.V. New perspectives on the role of the fibroblast growth factor family in amphibian development. Cell. Mol. Life Sci. 1997, 53, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Christen, B.; Slack, J.M. FGF-8 is associated with anteroposterior patterning and limb regeneration in Xenopus. Dev. Biol. 1997, 192, 455–466. [Google Scholar] [CrossRef]
- Junho, C.V.C.; Caio-Silva, W.; Trentin-Sonoda, M.; Carneiro-Ramos, M.S. An Overview of the Role of Calcium/Calmodulin-Dependent Protein Kinase in Cardiorenal Syndrome. Front. Physiol. 2020, 11, 735. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Yamamoto, T.S.; Ueno, N. Intracellular calcium signal at the leading edge regulates mesodermal sheet migration during Xenopus gastrulation. Sci. Rep. 2018, 8, 2433. [Google Scholar] [CrossRef] [PubMed]
- Wurzinger, B.; Mair, A.; Pfister, B.; Teige, M. Cross-talk of calcium-dependent protein kinase and MAP kinase signaling. Plant Signal. Behav. 2011, 6, 8–12. [Google Scholar] [CrossRef]
- Wernimont, A.K.; Artz, J.D.; Finerty, P., Jr.; Lin, Y.H.; Amani, M.; Allali-Hassani, A.; Senisterra, G.; Vedadi, M.; Tempel, W.; Mackenzie, F.; et al. Structures of apicomplexan calcium-dependent protein kinases reveal mechanism of activation by calcium. Nat. Struct. Mol. Biol. 2010, 17, 596–601. [Google Scholar] [CrossRef]
- Webb, S.E.; Miller, A.L. Ca2+ signaling and early embryonic patterning during the blastula and gastrula periods of zebrafish and Xenopus development. Biochim. Biophys. Acta 2006, 1763, 1192–1208. [Google Scholar] [CrossRef]
- Palma, V.; Kukuljan, M.; Mayor, R. Calcium mediates dorsoventral patterning of mesoderm in Xenopus. Curr. Biol. 2001, 11, 1606–1610. [Google Scholar] [CrossRef][Green Version]
- Neant, I.; Leung, H.C.; Webb, S.E.; Miller, A.L.; Moreau, M.; Leclerc, C. Trpc1 as the Missing Link Between the Bmp and Ca(2+) Signalling Pathways During Neural Specification in Amphibians. Sci. Rep. 2019, 9, 16049. [Google Scholar] [CrossRef] [PubMed]
- Neant, I.; Mellstrom, B.; Gonzalez, P.; Naranjo, J.R.; Moreau, M.; Leclerc, C. Kcnip1 a Ca(2)(+)-dependent transcriptional repressor regulates the size of the neural plate in Xenopus. Biochim. Biophys. Acta 2015, 1853, 2077–2085. [Google Scholar] [CrossRef]
- Leclerc, C.; Neant, I.; Moreau, M. The calcium: An early signal that initiates the formation of the nervous system during embryogenesis. Front. Mol. Neurosci. 2012, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Moreau, M.; Leclerc, C. The choice between epidermal and neural fate: A matter of calcium. Int. J. Dev. Biol. 2004, 48, 75–84. [Google Scholar] [CrossRef]
- Stern, C.D. Initial patterning of the central nervous system: How many organizers? Nat. Rev. Neurosci. 2001, 2, 92–98. [Google Scholar] [CrossRef]
- Kiecker, C.; Bates, T.; Bell, E. Molecular specification of germ layers in vertebrate embryos. Cell. Mol. Life Sci. 2016, 73, 923–947. [Google Scholar] [CrossRef]
- Dick, A.; Hild, M.; Bauer, H.; Imai, Y.; Maifeld, H.; Schier, A.F.; Talbot, W.S.; Bouwmeester, T.; Hammerschmidt, M. Essential role of Bmp7 (snailhouse) and its prodomain in dorsoventral patterning of the zebrafish embryo. Development 2000, 127, 343–354. [Google Scholar] [CrossRef]
- Nguyen, V.H.; Schmid, B.; Trout, J.; Connors, S.A.; Ekker, M.; Mullins, M.C. Ventral and lateral regions of the zebrafish gastrula, including the neural crest progenitors, are established by a bmp2b/swirl pathway of genes. Dev. Biol. 1998, 199, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Merker, S.; Lee, K.J.; McMahon, A.P.; Hammerschmidt, M. The zebrafish organizer requires chordino. Nature 1997, 387, 862–863. [Google Scholar] [CrossRef]
- Winnier, G.; Blessing, M.; Labosky, P.A.; Hogan, B.L. Bone morphogenetic protein-4 is required for mesoderm formation and patterning in the mouse. Genes Dev. 1995, 9, 2105–2116. [Google Scholar] [CrossRef]
- Tzahor, E.; Lassar, A.B. Wnt signals from the neural tube block ectopic cardiogenesis. Genes Dev. 2001, 15, 255–260. [Google Scholar] [CrossRef]
- Bates, T.J.; Vonica, A.; Heasman, J.; Brivanlou, A.H.; Bell, E. Coco regulates dorsoventral specification of germ layers via inhibition of TGFbeta signalling. Development 2013, 140, 4177–4181. [Google Scholar] [CrossRef]
- Vincent, S.D.; Dunn, N.R.; Hayashi, S.; Norris, D.P.; Robertson, E.J. Cell fate decisions within the mouse organizer are governed by graded Nodal signals. Genes Dev. 2003, 17, 1646–1662. [Google Scholar] [CrossRef]
- Lowe, L.A.; Yamada, S.; Kuehn, M.R. Genetic dissection of nodal function in patterning the mouse embryo. Development 2001, 128, 1831–1843. [Google Scholar] [CrossRef] [PubMed]
- Agathon, A.; Thisse, B.; Thisse, C. Morpholino knock-down of antivin1 and antivin2 upregulates nodal signaling. Genesis 2001, 30, 178–182. [Google Scholar] [CrossRef]
- Dougan, S.T.; Warga, R.M.; Kane, D.A.; Schier, A.F.; Talbot, W.S. The role of the zebrafish nodal-related genes squint and cyclops in patterning of mesendoderm. Development 2003, 130, 1837–1851. [Google Scholar] [CrossRef] [PubMed]
- Wessely, O.; Agius, E.; Oelgeschlager, M.; Pera, E.M.; De Robertis, E.M. Neural induction in the absence of mesoderm: Beta-catenin-dependent expression of secreted BMP antagonists at the blastula stage in Xenopus. Dev. Biol. 2001, 234, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Alev, C.; Wu, Y.; Nakaya, Y.; Sheng, G. Decoupling of amniote gastrulation and streak formation reveals a morphogenetic unity in vertebrate mesoderm induction. Development 2013, 140, 2691–2696. [Google Scholar] [CrossRef]
- Weinstein, D.C.; Marden, J.; Carnevali, F.; Hemmati-Brivanlou, A. FGF-mediated mesoderm induction involves the Src-family kinase Laloo. Nature 1998, 394, 904–908. [Google Scholar] [CrossRef]
- Umbhauer, M.; Marshall, C.J.; Mason, C.S.; Old, R.W.; Smith, J.C. Mesoderm induction in Xenopus caused by activation of MAP kinase. Nature 1995, 376, 58–62. [Google Scholar] [CrossRef]
- Mitrani, E.; Gruenbaum, Y.; Shohat, H.; Ziv, T. Fibroblast growth factor during mesoderm induction in the early chick embryo. Development 1990, 109, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, J.; Griffin, K.; Herbomel, P.; Dickmeis, T.; Strahle, U.; Kimelman, D.; Rosa, F.M.; Peyrieras, N. Nodal and Fgf pathways interact through a positive regulatory loop and synergize to maintain mesodermal cell populations. Development 2004, 131, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Kimelman, D.; Kirschner, M. Synergistic induction of mesoderm by FGF and TGF-beta and the identification of an mRNA coding for FGF in the early Xenopus embryo. Cell 1987, 51, 869–877. [Google Scholar] [CrossRef]
- Keenan, I.D.; Sharrard, R.M.; Isaacs, H.V. FGF signal transduction and the regulation of Cdx gene expression. Dev. Biol. 2006, 299, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Shiotsugu, J.; Katsuyama, Y.; Arima, K.; Baxter, A.; Koide, T.; Song, J.; Chandraratna, R.A.; Blumberg, B. Multiple points of interaction between retinoic acid and FGF signaling during embryonic axis formation. Development 2004, 131, 2653–2667. [Google Scholar] [CrossRef] [PubMed]
- Levy, V.; Marom, K.; Zins, S.; Koutsia, N.; Yelin, R.; Fainsod, A. The competence of marginal zone cells to become Spemann’s organizer is controlled by Xcad2. Dev. Biol. 2002, 248, 40–51. [Google Scholar] [CrossRef][Green Version]
- Amaya, E.; Musci, T.J.; Kirschner, M.W. Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos. Cell 1991, 66, 257–270. [Google Scholar] [CrossRef]
- Yao, J.; Kessler, D.S. Goosecoid promotes head organizer activity by direct repression of Xwnt8 in Spemann’s organizer. Development 2001, 128, 2975–2987. [Google Scholar] [CrossRef]
- Latinkic, B.V.; Umbhauer, M.; Neal, K.A.; Lerchner, W.; Smith, J.C.; Cunliffe, V. The Xenopus Brachyury promoter is activated by FGF and low concentrations of activin and suppressed by high concentrations of activin and by paired-type homeodomain proteins. Genes Dev. 1997, 11, 3265–3276. [Google Scholar] [CrossRef] [PubMed]
- Taira, M.; Saint-Jeannet, J.P.; Dawid, I.B. Role of the Xlim-1 and Xbra genes in anteroposterior patterning of neural tissue by the head and trunk organizer. Proc. Natl. Acad. Sci. USA 1997, 94, 895–900. [Google Scholar] [CrossRef] [PubMed]
- O’ Reilly, M.A.; Smith, J.C.; Cunliffe, V. Patterning of the mesoderm in Xenopus: Dose-dependent and synergistic effects of Brachyury and Pintallavis. Development 1995, 121, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Umair, Z.; Kumar, V.; Lee, U.; Choi, S.C.; Kim, J. Ventx1.1 competes with a transcriptional activator Xcad2 to regulate negatively its own expression. BMB Rep. 2019, 52, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Umair, Z.; Yoon, J.; Lee, U.; Kim, S.C.; Park, J.B.; Lee, J.Y.; Kim, J. Xbra and Smad-1 cooperate to activate the transcription of neural repressor ventx1.1 in Xenopus embryos. Sci. Rep. 2018, 8, 11391. [Google Scholar] [CrossRef]
- Sander, V.; Reversade, B.; De Robertis, E.M. The opposing homeobox genes Goosecoid and Vent1/2 self-regulate Xenopus patterning. EMBO J. 2007, 26, 2955–2965. [Google Scholar] [CrossRef]
- Ferreiro, B.; Artinger, M.; Cho, K.; Niehrs, C. Antimorphic goosecoids. Development 1998, 125, 1347–1359. [Google Scholar] [CrossRef]
- Talbot, W.S.; Trevarrow, B.; Halpern, M.E.; Melby, A.E.; Farr, G.; Postlethwait, J.H.; Jowett, T.; Kimmel, C.B.; Kimelman, D. A homeobox gene essential for zebrafish notochord development. Nature 1995, 378, 150–157. [Google Scholar] [CrossRef]
- Steinbeisser, H.; Fainsod, A.; Niehrs, C.; Sasai, Y.; De Robertis, E.M. The role of gsc and BMP-4 in dorsal-ventral patterning of the marginal zone in Xenopus: A loss-of-function study using antisense RNA. EMBO J. 1995, 14, 5230–5243. [Google Scholar] [CrossRef]
- Niehrs, C.; Pollet, N. Synexpression groups in eukaryotes. Nature 1999, 402, 483–487. [Google Scholar] [CrossRef]
- Umair, Z.; Kumar, S.; Kim, D.H.; Rafiq, K.; Kumar, V.; Kim, S.; Park, J.B.; Lee, J.Y.; Lee, U.; Kim, J. Ventx1.1 as a Direct Repressor of Early Neural Gene zic3 in Xenopus laevis. Mol. Cells 2018, 41, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Vonica, A.; Gumbiner, B.M. The Xenopus Nieuwkoop center and Spemann-Mangold organizer share molecular components and a requirement for maternal Wnt activity. Dev. Biol. 2007, 312, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Larrain, J.; Oelgeschlager, M.; Ketpura, N.I.; Reversade, B.; Zakin, L.; De Robertis, E.M. Proteolytic cleavage of Chordin as a switch for the dual activities of Twisted gastrulation in BMP signaling. Development 2001, 128, 4439–4447. [Google Scholar] [CrossRef] [PubMed]
- Malaguti, M.; Nistor, P.A.; Blin, G.; Pegg, A.; Zhou, X.; Lowell, S. Bone morphogenic protein signalling suppresses differentiation of pluripotent cells by maintaining expression of E-Cadherin. Elife 2013, 2, e01197. [Google Scholar] [CrossRef]
- Kuo, C.L.; Lam, C.M.; Hewitt, J.E.; Scotting, P.J. Formation of the embryonic organizer is restricted by the competitive influences of Fgf signaling and the SoxB1 transcription factors. PLoS ONE 2013, 8, e57698. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Goutam, R.S.; Umair, Z.; Park, S.; Lee, U.; Kim, J. Foxd4l1.1 Negatively Regulates Chordin Transcription in Neuroectoderm of Xenopus Gastrula. Cells 2021, 10, 2779. [Google Scholar] [CrossRef]
- Rogers, C.D.; Moody, S.A.; Casey, E.S. Neural induction and factors that stabilize a neural fate. Birth Defects Res. C Embryo Today 2009, 87, 249–262. [Google Scholar] [CrossRef]
- Munoz-Sanjuan, I.; Brivanlou, A.H. Neural induction, the default model and embryonic stem cells. Nat. Rev. Neurosci. 2002, 3, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.A.; Hemmati-Brivanlou, A. Induction of epidermis and inhibition of neural fate by Bmp-4. Nature 1995, 376, 331–333. [Google Scholar] [CrossRef]
- Suzuki, A.; Ueno, N.; Hemmati-Brivanlou, A. Xenopus msx1 mediates epidermal induction and neural inhibition by BMP4. Development 1997, 124, 3037–3044. [Google Scholar] [CrossRef]
- Hawley, S.H.; Wunnenberg-Stapleton, K.; Hashimoto, C.; Laurent, M.N.; Watabe, T.; Blumberg, B.W.; Cho, K.W. Disruption of BMP signals in embryonic Xenopus ectoderm leads to direct neural induction. Genes Dev. 1995, 9, 2923–2935. [Google Scholar] [CrossRef] [PubMed]
- Sasal, Y.; Lu, B.; Steinbelsser, H.; De Robertis, E.M. Regulation of neural induction by the Chd and Bmp-4 antagonistic patterning signals in Xenopus. Nature 1995, 378, 419. [Google Scholar] [CrossRef] [PubMed]
- Iemura, S.; Yamamoto, T.S.; Takagi, C.; Kobayashi, H.; Ueno, N. Isolation and characterization of bone morphogenetic protein-binding proteins from the early Xenopus embryo. J. Biol. Chem. 1999, 274, 26843–26849. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yabe, T.; Shimizu, T.; Muraoka, O.; Bae, Y.K.; Hirata, T.; Nojima, H.; Kawakami, A.; Hirano, T.; Hibi, M. Ogon/Secreted Frizzled functions as a negative feedback regulator of Bmp signaling. Development 2003, 130, 2705–2716. [Google Scholar] [CrossRef]
- Miller-Bertoglio, V.; Carmany-Rampey, A.; Furthauer, M.; Gonzalez, E.M.; Thisse, C.; Thisse, B.; Halpern, M.E.; Solnica-Krezel, L. Maternal and zygotic activity of the zebrafish ogon locus antagonizes BMP signaling. Dev. Biol. 1999, 214, 72–86. [Google Scholar] [CrossRef]
- Khokha, M.K.; Yeh, J.; Grammer, T.C.; Harland, R.M. Depletion of three BMP antagonists from Spemann’s organizer leads to a catastrophic loss of dorsal structures. Dev. Cell 2005, 8, 401–411. [Google Scholar] [CrossRef]
- Bouwmeester, T.; Kim, S.; Sasai, Y.; Lu, B.; De Robertis, E.M. Cerberus is a head-inducing secreted factor expressed in the anterior endoderm of Spemann’s organizer. Nature 1996, 382, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.S.; Marion, C.D.; Steele, K.; George, S.; Smith, W.C. Direct neural induction and selective inhibition of mesoderm and epidermis inducers by Xnr3. Development 1997, 124, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Hemmati-Brivanlou, A. Cell fate determination in embryonic ectoderm. J. Neurobiol. 1998, 36, 128–151. [Google Scholar] [CrossRef]
- Ishimura, A.; Maeda, R.; Takeda, M.; Kikkawa, M.; Daar, I.O.; Maeno, M. Involvement of BMP-4/msx-1 and FGF pathways in neural induction in the Xenopus embryo. Dev. Growth Differ. 2000, 42, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Gawantka, V.; Delius, H.; Hirschfeld, K.; Blumenstock, C.; Niehrs, C. Antagonizing the Spemann organizer: Role of the homeobox gene Xvent-1. EMBO J. 1995, 14, 6268–6279. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.J.; Wang, T.; Kroll, K.L. Tcf- and Vent-binding sites regulate neural-specific geminin expression in the gastrula embryo. Dev. Biol. 2006, 289, 494–506. [Google Scholar] [CrossRef]
- Rogers, C.D.; Archer, T.C.; Cunningham, D.D.; Grammer, T.C.; Casey, E.M. Sox3 expression is maintained by FGF signaling and restricted to the neural plate by Vent proteins in the Xenopus embryo. Dev. Biol. 2008, 313, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Harland, R.M. Neural induction requires continued suppression of both Smad1 and Smad2 signals during gastrulation. Development 2007, 134, 3861–3872. [Google Scholar] [CrossRef] [PubMed]
- Delaune, E.; Lemaire, P.; Kodjabachian, L. Neural induction in Xenopus requires early FGF signalling in addition to BMP inhibition. Development 2005, 132, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Li, L.; Huang, C.; Shen, C.; Tan, F.; Xia, C.; Liu, P.; Rossant, J.; Jing, N. Distinct functions of BMP4 during different stages of mouse ES cell neural commitment. Development 2010, 137, 2095–2105. [Google Scholar] [CrossRef] [PubMed]
- Linker, C.; Stern, C.D. Neural induction requires BMP inhibition only as a late step, and involves signals other than FGF and Wnt antagonists. Development 2004, 131, 5671–5681. [Google Scholar] [CrossRef] [PubMed]
- Stern, C.D. Neural induction: 10 years on since the ’ default model’ . Curr. Opin. Cell Biol. 2006, 18, 692–697. [Google Scholar] [CrossRef]
- Kumar, S.; Umair, Z.; Kumar, V.; Kumar, S.; Lee, U.; Kim, J. Foxd4l1.1 negatively regulates transcription of neural repressor ventx1.1 during neuroectoderm formation in Xenopus embryos. Sci. Rep. 2020, 10, 16780. [Google Scholar] [CrossRef] [PubMed]
- Kazanskaya, O.; Glinka, A.; Niehrs, C. The role of Xenopus dickkopf1 in prechordal plate specification and neural patterning. Development 2000, 127, 4981–4992. [Google Scholar] [CrossRef] [PubMed]
- Glinka, A.; Wu, W.; Delius, H.; Monaghan, A.P.; Blumenstock, C.; Niehrs, C. Dickkopf-1 is a member of a new family of secreted proteins and functions in head induction. Nature 1998, 391, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.I.; Rydstrom, A.; Trimborn, T.; Willert, K.; Nusse, R.; Jessell, T.M.; Edlund, T. The status of Wnt signalling regulates neural and epidermal fates in the chick embryo. Nature 2001, 411, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Elkouby, Y.M.; Elias, S.; Casey, E.S.; Blythe, S.A.; Tsabar, N.; Klein, P.S.; Root, H.; Liu, K.J.; Frank, D. Mesodermal Wnt signaling organizes the neural plate via Meis3. Development 2010, 137, 1531–1541. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, H.V.; Pownall, M.E.; Slack, J.M. Regulation of Hox gene expression and posterior development by the Xenopus caudal homologue Xcad3. EMBO J. 1998, 17, 3413–3427. [Google Scholar] [CrossRef]
- Pera, E.M.; Ikeda, A.; Eivers, E.; De Robertis, E.M. Integration of IGF, FGF, and anti-BMP signals via Smad1 phosphorylation in neural induction. Genes Dev. 2003, 17, 3023–3028. [Google Scholar] [CrossRef] [PubMed]
- Furthauer, M.; Thisse, C.; Thisse, B. A role for FGF-8 in the dorsoventral patterning of the zebrafish gastrula. Development 1997, 124, 4253–4264. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.A.; Lagna, G.; Suzuki, A.; Hemmati-Brivanlou, A. Concentration-dependent patterning of the Xenopus ectoderm by BMP4 and its signal transducer Smad1. Development 1997, 124, 3177–3184. [Google Scholar] [CrossRef] [PubMed]
- Hongo, I.; Kengaku, M.; Okamoto, H. FGF signaling and the anterior neural induction in Xenopus. Dev. Biol. 1999, 216, 561–581. [Google Scholar] [CrossRef][Green Version]
- Lim, M.S.; Lee, S.Y.; Park, C.H. FGF8 is Essential for Functionality of Induced Neural Precursor Cell-derived Dopaminergic Neurons. Int. J. Stem Cells 2015, 8, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Pownall, M.E.; Isaacs, H.V.; Slack, J.M. Two phases of Hox gene regulation during early Xenopus development. Curr. Biol. 1998, 8, 673–676. [Google Scholar] [CrossRef]
- Cho, K.W.; De Robertis, E.M. Differential activation of Xenopus homeo box genes by mesoderm-inducing growth factors and retinoic acid. Genes Dev. 1990, 4, 1910–1916. [Google Scholar] [CrossRef] [PubMed]
- Haremaki, T.; Tanaka, Y.; Hongo, I.; Yuge, M.; Okamoto, H. Integration of multiple signal transducing pathways on Fgf response elements of the Xenopus caudal homologue Xcad3. Development 2003, 130, 4907–4917. [Google Scholar] [CrossRef]
- Zorn, A.M.; Wells, J.M. Vertebrate endoderm development and organ formation. Annu. Rev. Cell Dev. Biol. 2009, 25, 221–251. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Rankin, S.A.; Sinner, D.; Kenny, A.P.; Krieg, P.A.; Zorn, A.M. Sfrp5 coordinates foregut specification and morphogenesis by antagonizing both canonical and noncanonical Wnt11 signaling. Genes Dev. 2008, 22, 3050–3063. [Google Scholar] [CrossRef] [PubMed]
- Villegas, S.N.; Rothova, M.; Barrios-Llerena, M.E.; Pulina, M.; Hadjantonakis, A.K.; Le Bihan, T.; Astrof, S.; Brickman, J.M. PI3K/Akt1 signalling specifies foregut precursors by generating regionalized extra-cellular matrix. Elife 2013, 2, e00806. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, V.; Park, S.; Lee, U.; Kim, J. The Organizer and Its Signaling in Embryonic Development. J. Dev. Biol. 2021, 9, 47. https://doi.org/10.3390/jdb9040047
Kumar V, Park S, Lee U, Kim J. The Organizer and Its Signaling in Embryonic Development. Journal of Developmental Biology. 2021; 9(4):47. https://doi.org/10.3390/jdb9040047
Chicago/Turabian StyleKumar, Vijay, Soochul Park, Unjoo Lee, and Jaebong Kim. 2021. "The Organizer and Its Signaling in Embryonic Development" Journal of Developmental Biology 9, no. 4: 47. https://doi.org/10.3390/jdb9040047
APA StyleKumar, V., Park, S., Lee, U., & Kim, J. (2021). The Organizer and Its Signaling in Embryonic Development. Journal of Developmental Biology, 9(4), 47. https://doi.org/10.3390/jdb9040047