Abstract
Dynein is a multi-subunit motor protein that moves toward the minus-end of microtubules, and plays important roles in fly development. We identified Dhc64Cm115, a new mutant allele of the fly Dynein heavy chain 64C (Dhc64C) gene whose heterozygotes survive against lethality induced by overexpression of Sol narae (Sona). Sona is a secreted metalloprotease that positively regulates Wingless (Wg) signaling, and promotes cell survival and proliferation. Knockdown of Dhc64C in fly wings induced extensive cell death accompanied by widespread and disorganized expression of Wg. The disrupted pattern of the Wg protein was due to cell death of the Wg-producing cells at the DV midline and overproliferation of the Wg-producing cells at the hinge in disorganized ways. Coexpression of Dhc64C RNAi and p35 resulted in no cell death and normal pattern of Wg, demonstrating that cell death is responsible for all phenotypes induced by Dhc64C RNAi expression. The effect of Dhc64C on Wg-producing cells was unique among components of Dynein and other microtubule motors. We propose that Dhc64C differentially regulates survival of Wg-producing cells, which is essential for maintaining normal expression pattern of Wg for wing development.
1. Introduction
Apoptosis is a programmed cell death critical for the development and homeostasis of all organisms, which functions by eliminating unwanted cells generated under both normal and stress conditions [1]. Apoptosis often occurs concomitantly with compensatory proliferation to maintain tissue size and shape [2]. Drosophila melanogaster is a great model system for studying apoptosis and compensatory proliferation whose mechanism and components are highly conserved during evolution [3,4,5]. The wing imaginal disc, the primordium of the adult wing, is especially suitable for studying these processes because it shows little apoptosis during normal development but develops to a normal wing even with 40–60% of apoptotic cells via efficient compensatory proliferation [6].
The family of A Disintegrin and Metalloproteinase with ThromboSpondin motifs (ADAMTS) has important roles in cell proliferation, cell survival, cell migration, blood coagulation, angiogenesis in mammals, and their malfunctions result in numerous diseases including cancer, arthrosclerosis, and arthritis [7,8,9,10]. Sol narae (Sona) is a member of fly ADAMTS proteins, and positively regulates Wg signaling by promoting secretion of Wg [11]. Sona is also important for cell survival and compensatory proliferation by increasing the level of Cyclin D for cell division in order to maintain tissue size and shape [11,12].
To understand the function of Sona, our laboratory previously carried out a genetic screen, and identified suppressors that overcome lethality induced by overexpressed Sona. Several suppressors that have mutations in wntless, pou domain factor 3 (pdm3), archipelago (ago), arrow (arr) and anastral spindle 3 genes have been reported [11,13,14,15,16]. The focus of this study is m115, one of the suppressors that has a missense mutation in the Dhc64C gene. Fly Dhc64C is a component of cytoplasmic Dynein, a microtubule force-producing protein motor moving toward the minus end of microtubules essential for organelle transport and centrosome assembly [16]. Cytoplasmic Dynein is a macromolecular complex that contains two heavy, two intermediate, two light intermediate and several light chains. The two identical heavy chains in Dynein form dimers and have motor machinery that transduces chemical energy derived from ATP hydrolysis into mechanical force [17,18].
We report here that sona and Dhc64C show a positive genetic interaction, and the knockdown of Dhc64C induces apoptosis of Wg-producing cells in the DV midline, whereas uncontrolled compensatory proliferation of Wg-producing cells originated from the hinge region, which resulted in abnormally high numbers of Wg-producing cells in entire wing discs. Knockdown of genes encoding other subunits in Dynein or Kinesin did not show such phenotypes, suggesting that Dhc64C plays a unique role in cell survival and proliferation of Wg-producing cells in fly wing discs.
2. Materials and Methods
2.1. Fly Strains
Gal4 drivers such as patched (ptc)-Gal4, 30A-Gal4, nubbin (nub)-Gal4, engrailed (en)-Gal4, C96-gal4, UAS lines such as UAS-GFP, UAS-Dhc64C RNAi (BL28749), UAS-Khc RNAi (BL35770), and flies for mapping were obtained from Bloomington Stock Center. UAS-cut up RNAi (v43115), UAS-Klp64D RNAi (NIG10642R-1) and UAS-Dhc64C RNAi (NIG7507R-2) were obtained from Vienna Drosophila Resource Center and National Institute of Genetics. Deficiency lines (Df(3L)BSC436, Df(3L)ED210, Df(3L)Exel6102) that uncover the Dhc64C gene and, Dhc64C6.6−16 (BL32015) were used for complementation test of Dhc64C. We used UAS-Sona that had been generated in previous study [11]. Fly cultures were carried out at 25 °C unless indicated otherwise.
2.2. Immunohistochemistry
Wing discs of the third instar larvae were dissected, fixed, blocked, and incubated with the primary antibodies, as described [19]. After washing several times, samples were incubated with secondary antibodies in washing buffer for 2 h at room temperature and stained with 4′, 6-diamidino-2-phenylindole (DAPI). Then, samples were mounted using Vectashield (Vector Laboratories, Burlingame, CA, USA). All confocal images were acquired using Zeiss LSM710 confocal microscope and ZEN software.
Primary antibodies were used in the following dilutions: chicken anti-β-gal (ab 9361), 1:100; rabbit anti-cleaved Caspase-3 (Cell Signaling Technology, Danvers, MA, USA), 1:250; rabbit anti-Dlg, 1:500 [20], mouse anti-Wg (DSHB #4D4), 1:100; mouse anti-Cut (DSHB #2B10), 1:100; guinea pig anti-Senseless (gift from Hugo Bellen), 1:1000 sheep anti-GFP (Ab direct serotec), 1:100. Secondary antibodies conjugated with Cy3 (1:200), Cy5 (1:500) and FITC (1:200) were from Alexa Flour (Jackson Immunoresearches, West Grove, PA, USA).
2.3. Adult Wing Mounting
The left wings of 2–7 days old flies were dissected and mounted in Gary’s magic mounting solution (Canada Balsam and methyl salicylate, 4:1). The wing images were obtained using Zeiss Axio imager M2 and Axiocam software. All wing images were taken with 50× magnification.
3. Results
3.1. A Sona Suppressor m115 has a Mutation in the ATPases Associated with Various Cellular Activities 3 (AAA3) Domain of Dhc64C Protein
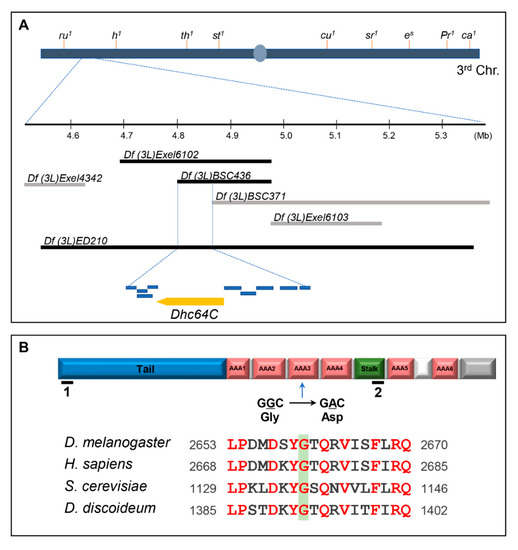
Heterozygous m115 was identified in a genetic screen as one of the 28 suppressors that overcome the lethality induced by Sona overexpression [11,14,16]. Homozygous m115 was embryonic lethal, so we mapped the lethal site by meiotic and deficiency mapping under the assumption that the lethal mutation of the m115 is responsible for suppression of Sona-induced lethality in m115 heterozygotes. Meiotic mapping localized the lethal site between the two markers in the third chromosome, roughoid (61F8) and hairy (66D10) (Supplementary Table S1). Subsequent deficiency mapping narrowed down the lethal site between 64B17 and 64C1, in which ten genes are present (Figure 1A). To identify the gene that harbors the lethal site, we then carried out complementation tests. One of the mutants that did not complement the lethality of m115 was Dhc64C6.6−16, a hypomorph that can develop to adulthood (Supplementary Table S2).
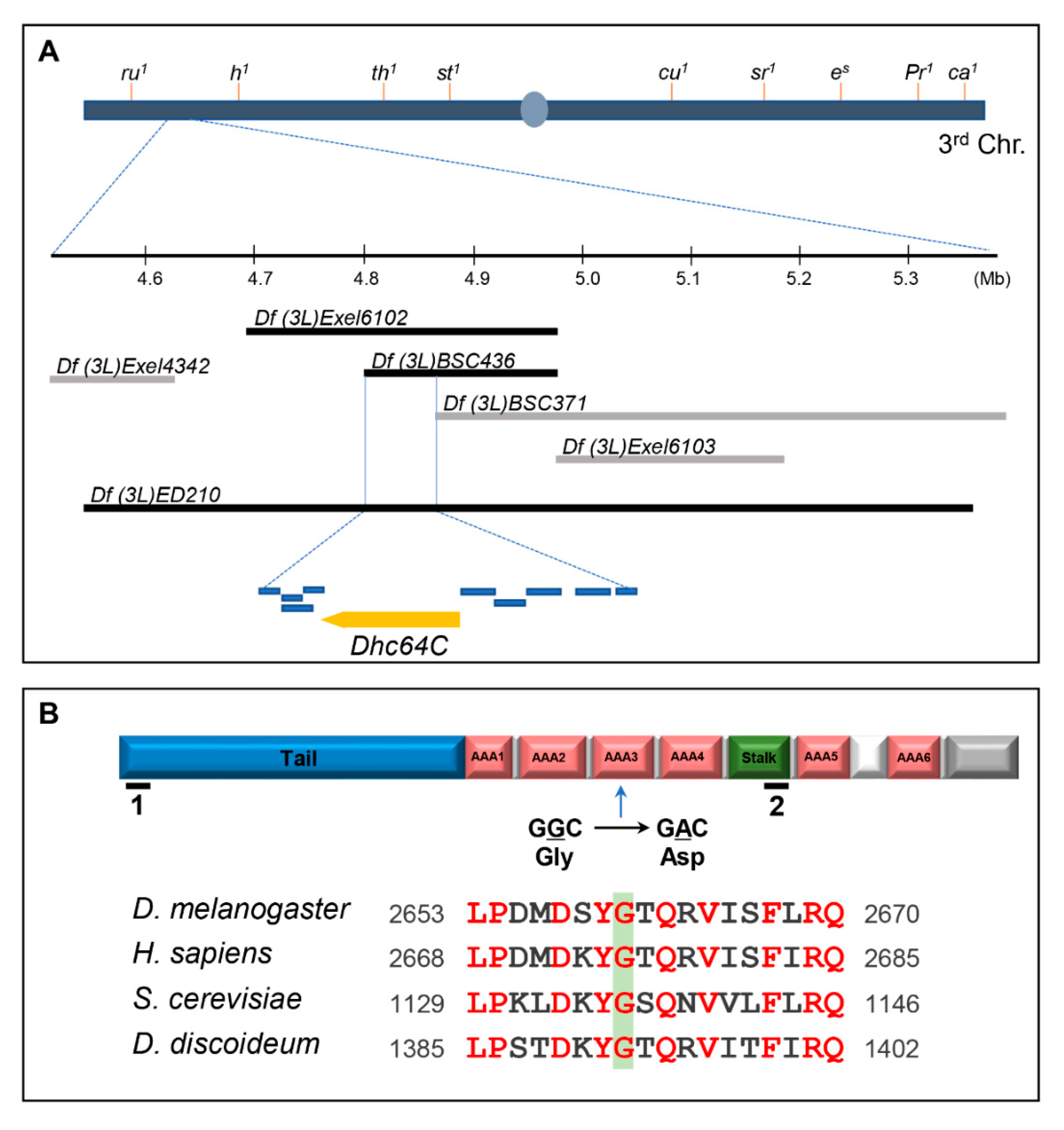
Figure 1.
The lethal site in the sona suppressor m115 is mapped to the Dhc64C gene. (A) Mapping of the lethal site in m115 by meiotic and deficiency mapping. Meiotic mapping localized the lethal site in m115 between ru and h, which is further explained in Table S1. Multiple deficiencies were used to narrow it down to 64B17-64C1. Deleted range in the two deficiencies that uncover the lethal site of m115 are marked with black bars. Other deficiencies are marked with gray bars. Dhc64C and nine genes in the region common in the two deficiencies are presented as yellow and blue bars, respectively. Their names are listed in Table S2. (B) The domain structure of Dynein heavy chain. A point mutation from G to A results in one amino acid change from glycine (G) to aspartic acid (D) in the AAA3 domain of Dhc64Cm115. The region including the mutated site of Dhc64Cm115 is conserved in Dhc proteins of model organisms. Red letters indicate amino acid residues conserved in these organisms. RNAi target site for BL28749 and NIG7507R-2 are marked by black lines with number 1 and 2, respectively.
Genomic sequencing of Dhc64Cm115 flies identified a guanine to adenosine transition that changes glycine to aspartic acid in the AAA3 domain of the Dhc64C protein, which is consistent with the mechanism of the mutagen ethyl methanesulfonate (EMS) used in our genetic screen, guanine alkylation, which changes GC pairs to AT pairs [11,14]. This glycine residue is conserved in all cytoplasmic Dynein heavy chain proteins examined (Figure 1B). The ATPase activity of AAA3 domain is especially important for release of Dynein from microtubules for the subsequent movement of Dynein along the microtubules [21]. Dhc64C is most homologous to the human cytoplasmic Dynein 1 heavy chain 1 (5NUG_A) with 72% identity and 85% similarity. It also shows 72% and 57% similarity to Dhc of D. discoideum and S. cerevisiae, respectively.
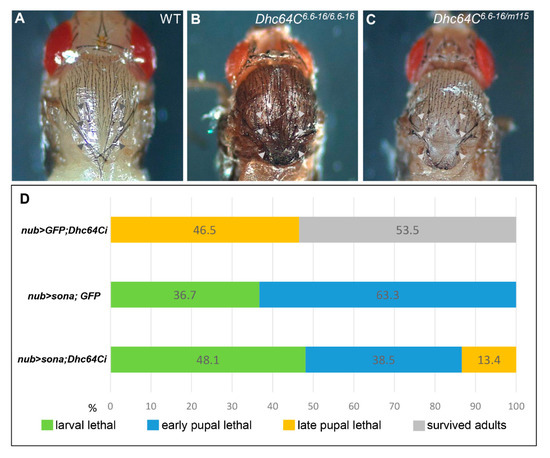
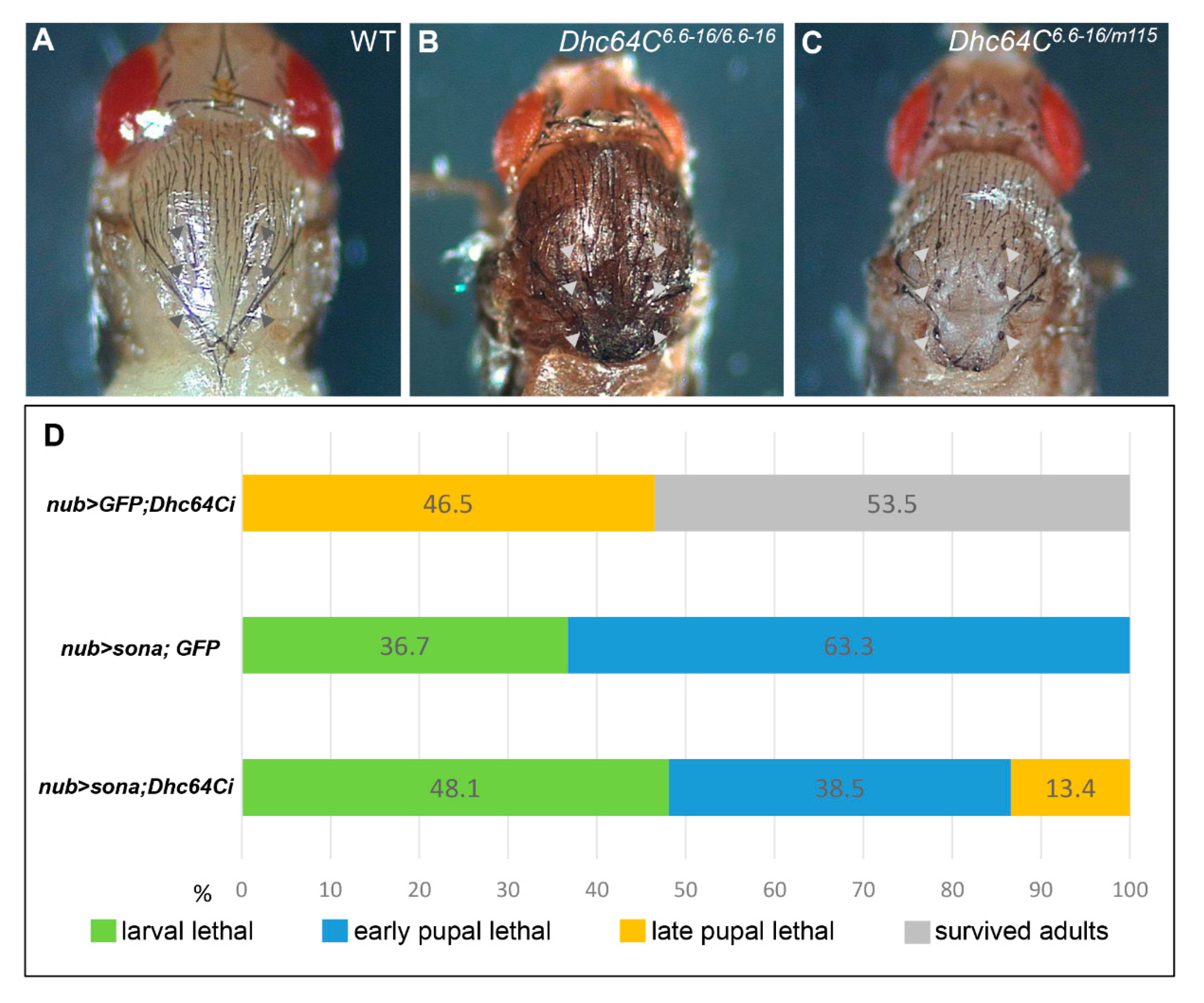
As mentioned above, Dhc64C6.6−16 can develop to adults, but m115/Dhc64C6.6−16 only survived up pharate adults, indicating that the mutation in m115 is responsible for the lethality of the heterozygotes. Pupae of both m115/Dhc64C6.6−16 transheterozygotes and Dhc64C6.6−16 homozygotes had short and deformed scutellar bristles, also indicating that bristle phenotype of Dhc64C6.6−16 is not complemented by m115 (Figure 2A–C). This bristle phenotype is consistent with the importance of Dhc64C in bristle formation [22]. In sum, both lethality and bristle defects are due to the mutation in the Dhc64C gene of m115, so m115 was named Dhc64Cm115 as a new Dhc64C allele.
Figure 2.
Dhc64C has a positive genetic interaction with sona. (A–C) Scutellar bristles are defective in Dhc64C mutants. The normal scutellum of wild type Canton-S (CS) pupae ((A), arrowheads). Scutellar bristles are short and disrupted in hypomorphic Dhc64C6.6−16/6.6−16 adults ((B), arrowheads) and transheterozygous Dhc64C6.6−16/Dhc64Cm115 pharate adults ((C), arrowheads). (D) Knockdown of Dhc64C partially rescues Sona-induced lethality. Fly genotypes and developmental states in percentages are shown.
3.2. Knockdown of Dhc64C Reduces the Severity of Sona-Induced Lethality
We then examined whether the reduction in the level of wild-type Dhc64C by Dhc64C RNAi expression is as efficient as Dhc64Cm115 protein in suppressing Sona-induced lethality. For this work, we tested two Dhc64C RNAi (Dhc64Ci) lines, BL28749 and VDRC 7507R-2, covering different parts of the Dhc64C transcript (Figure 1B-1,2). Their expression by crossing with various Gal4 lines at 25 °C resulted in embryonic to late pupal lethality with small-sized larvae (Supplementary Table S3). Since both RNAi lines showed similar phenotypes, BL28749 was chosen for the knockdown of Dhc64C study in this report.
We generated UAS-GFP; UAS-Dhc64Ci, UAS-sona; UAS-GFP, and UAS-sona; UAS-Dhc64Ci flies, crossed them with nub-Gal4, and compared the lethal rate of their progenies at 18 °C (Figure 2D). All nub > sona; GFP flies (n = 109) were larval or early pupal lethal, while 46.5% of nub-Gal4 > UAS-GFP; UAS-Dhc64Ci (nub > GFP; Dhc64Ci) flies (n = 43) were late pupal lethal. In contrast, 13.4% of nub > sona; Dhc64Ci flies (n = 52) could develop up to late pupal stage. Therefore, the knockdown of Dhc64C reduced the severity of Sona-induced lethality, though less efficiently than the Dhc64Cm115 allele. These results are consistent with the identification of Dhc64Cm115 as a sona suppressor in the aforementioned genetic screen.
3.3. Knockdown of Dhc64C Induces Apoptosis
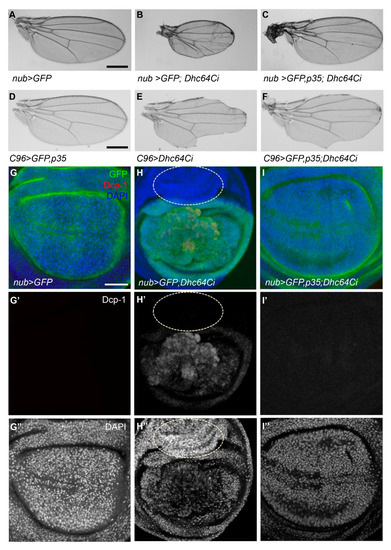
Since the knockdown of Sona induces cell death and generates small wing phenotype [11,12], we checked whether the knockdown of Dhc64C also generates small wings using multiple Gal4 lines. Some nub > Dhc64Ci flies at 18 °C developed into adults as described (Figure 2D), and all their wings were either absent or smaller than control nub > GFP wings (n > 20, each) (Figure 3A,B). With a similar tendency, all C96 > Dhc64Ci flies (n = 12) had notched wings (Figure 3D,E). All ptc > GFP; Dhc64Ci wings (n > 20) were not flat with some abnormally grown structures, their wing size was also smaller than wild-type wings, and the region between L3 and L4 veins was narrower than wild-type wings at 22 °C (Supplementary Figure S1A,B). Therefore, knockdown of Dhc64C by multiple Gal4 lines results in small or notched wings.
Figure 3.
The knockdown of Dhc64C induces apoptosis. (A–C) Knockdown of Dhc64C induces small wing phenotype, which can be rescued by coexpression of p35. Compared to control nub-Gal4/+ wings (A), nub > GFP; Dhc64Ci wings from rare survivors cultured at 18 °C are small and wrinkled (B), and nub > GFP, p35; Dhc64Ci wings are normal (C). Scale bar 500 µm. n = 10 each. (D–F) Compared to the normal C96 > GFP, p35 wings (D), C96 > Dhc64Ci wings are severely notched (E). This notching phenotype of C96 > GFP; Dhc64Ci wings is noticeably rescued by p35 (F). Scale bar, 500 µm. n = 10 each (G–I) Knockdown of Dhc64C in the wing pouch region induces both apoptosis and cell proliferation, which are rescued by p35. Activated Dcp1 is detected in wing discs of the nub > GFP; Dhc64Ci flies (H’), which are not detected from nub > GFP and nub > GFP, p35; Dhc64Ci (G’,I’, respectively) DAPI staining shows overall organization of wing pouch cells (G”–I”). All apoptosis-related phenomena are rescued by coexpression of p35 (I). Abnormal growth of hinge region outside of nub-Gal4-expressing region is marked with dotted circles (H–H’’). Anterior to the left. Scale bar 100 µm.
We then tested whether apoptosis induced by the knockdown of Dhc64C is responsible for these small or notched wing phenotypes. Indeed, co-expression of Dhc64C RNAi and p35 restored wing size of nub > Dhc64Ci and reduced notching phenotype of C96 > Dhc64Ci flies (Figure 3C,F). Consistent with these adult wing phenotypes, nub > GFP; Dhc64Ci wing discs had activated Drosophila caspase 1 (Dcp-1) signal in the GFP-expressing region, while control nub > GFP and nub > GFP, p35; Dhc64Ci wing discs had no Dcp-1 signal in all wing discs examined (Figure 3G’–I’). Furthermore, p35 completely normalized overall organization of nub > GFP, p35; Dhc64Ci wing discs compared to nub > GFP; Dhc64Ci wing discs (Figure 3G’’–I’’). These demonstrate that the knockdown of Dhc64C induces apoptosis.
Notched wing phenotype of C96 > Dhc64Ci flies may also be due to cell death, specifically at the DV midline region (Figure 3D–F). We therefore checked whether Senseless (Sens), a transcription factor involved in the differentiation of wing margin cells into sensory organs, is affected in C96 > Dhc64Ci flies [23,24]. Sens was not properly expressed at the DV midline in C96 > Dhc64Ci wings compared to control wings (Supplementary Figure S2A,B). Co-expression of Dhc64C RNAi with p35 partially restored Sens expression (Supplementary Figure S2C). In sum, apoptosis by knockdown of Dhc64C is responsible for small or notched wing phenotypes.
3.4. Knockdown of Dhc64C Induces Overproliferation in a Non-Cell Autonomous Manner
We have shown above that GFP-positive (GFP+) cells in nub > GFP; Dhc64Ci discs exhibit cell death because of Dhc64 RNAi expression driven by the nub-Gal4 driver, whereas GFP-negative (GFP−) cells in these discs without any Dhc64 RNAi expression showed no sign of cell death (Figure 3H). Interestingly, some GFP− cells in the region of outer ring hinge showed overproliferation and appeared to move into the wing pouch region where GFP+ cells are present (Figure 3H–H”). Obviously, these overproliferated GFP− cells did not express apoptotic marker DCP-1 because Dhc64C RNAi is not expressed in these cells (dotted circle in Figure 3H–H’’). In another nub > GFP; Dhc64Ci disc, these GFP− cells were present in between GFP+ cells probably by moving into the pouch from the hinge region (red arrows in Supplementary Figure S1C). This phenomenon implied that cell death by the knockdown of Dhc64C induces overproliferation of neighboring cells in a non-cell autonomous manner.
When p35 was coexpressed with Dhc64C RNAi in nub > GFP, p35; Dhc64Ci wing discs, neither cell death of GFP+ cells nor overproliferation of GFP- cells were detected (Figure 3I–I’’). Therefore, we concluded that the knockdown of Dhc64C induces cell death in a cell-autonomous manner and then overproliferation of neighboring cells in a non-cell-autonomous manner.
3.5. The Knockdown of Dhc64C Increases the Level of Wg in the Entire Wing Pouch
Co-occurrence of apoptosis and cell proliferation by the knockdown of Dhc64C was an intriguing phenomenon, so we examined whether the knockdown of Dhc64C affects the expression level or pattern of Wg based on the role of Sona in Wg signaling [11,25]. Wg signaling is known to be involved in compensatory proliferation and hyperplastic overgrowth caused by apoptotic cells [26].
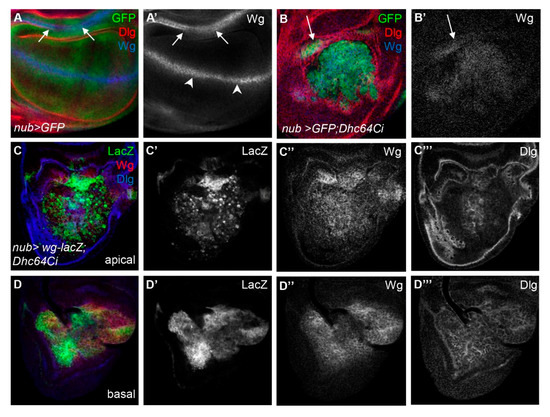
We therefore examined the pattern of Wg protein in nub > GFP; Dhc64Ci wing discs. We found that Wg protein was present in the entire pouch of nub > GFP; Dhc64Ci wing discs, unlike the control nub > GFP wing discs in which Wg was present at the DV midline and the inner ring hinge (Figure 4A,B). To check whether the change in Wg pattern by the knockdown of Dhc64C is a phenotype common to other components in microtubule motors, we knocked down the level of cut up (ctp), which encodes a light chain of cytoplasmic Dynein [27,28], Kinesin heavy chain (Khc), which encodes the heavy chain of Kinesin-1 [29,30], and Kinesin-like protein at 64D (Klp64D), which encodes a motor subunit of the Kinesin-2 [31,32] (Supplementary Figure S3). None of them showed a Wg pattern similar to that of Dhc64C knockdown, suggesting that Dhc64C is unique among microtubule motor subunits whose loss results in overall increase in Wg pattern in the entire pouch.
Figure 4.
Knockdown of Dhc64C disrupts expression pattern of Wg. (A–A’,B–B’) Knockdown of Dhc64C results in broad expression of Wg in the wing pouch. Pattern of Wg protein at the DV midline (arrowheads) and in the inner ring (arrows) in control wing disc (A,A’) and extensively disorganized and widely distributed pattern of Wg in severely affected nub > GFP; Dhc64Ci wing discs (B,B’). (C–C’’,D–D’’) The broad expression of Wg protein is due to widespread Wg-producing cells in both apical (C–C’’) and basal (D–D’’) regions of nub > GFP; Dhc64Ci wing discs.The expression pattern of the Dlg is also disrupted both in apical (C’’’) and basal (D’’’).
3.6. Knockdown of Dhc64C Induces Cell Death of Wg-Expressing Cells in the Apical Region
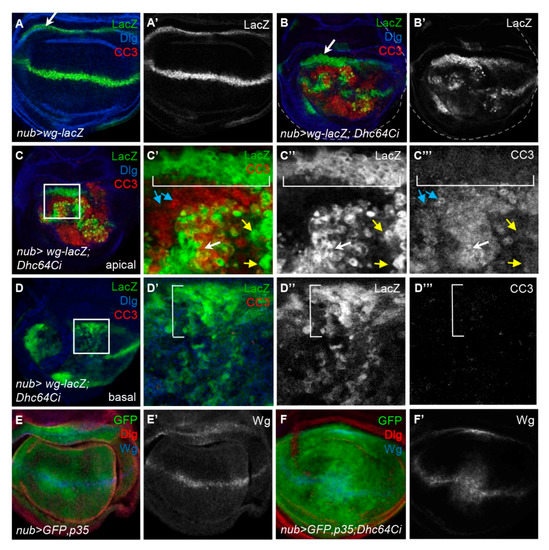
We then examined how the pattern of Wg is changed by the knockdown of Dhc64C. The pattern of Wg protein was similar to that of Wg-lacZ cells in both apical and basal regions of nub > wg-lacZ; Dhc64Ci wing discs, demonstrating that the widespread Wg-expressing cells are responsible for the broad distribution of Wg protein (Figure 4C,D). One noticeable difference between Wg-lacZ cells the apical and basal regions was the spottiness of the Wg-lacZ cells in the apical but not the basal region (Figure 4C’,D’).
The spotty Wg-lacZ cells in the apical region of nub > wg-lacZ; Dhc64Ci wing discs indeed had the cleaved Caspase 3 (CC3), a marker for apoptotic cells [30] (Figure 5A,B). Consistent with this result, apoptosis was most severe in the apical region, gradually decreased toward the basal region, and was almost absent in the basal region (Supplementary Figure S4). This suggests that the apically located cells are more prone to cell death by the knockdown of Dhc64C.
Figure 5.
Knockdown of Dhc64C induces cell death and cell proliferation of Wg-producing cells in the apical and basal region of wing discs, respectively. (A–A’,B–B’) Comparison of Wg-lacZ pattern between a nub-Gal4 > wg-lacZ control disc (A,A’) and a nub-Gal4 > wg-lacZ; Dhc64Ci wing disc (B,B’) at the basolateral region. (B) is one of Z-stack images of a nub-Gal4 > wg-lacZ; Dhc64Ci wing disc in Figure S4. (C,D) Apoptotic cells are present in the apical region but not in the basal region. Images representing apical (C) and basal (D) region of a nub-Gal4 > wg-lacZ; Dhc64Ci wing disc are selected from the same Z-stack images in Figure S4. The boxed regions in C and D are magnified in C’–C’’’ and D’–D’’’. Apically located Wg-lacZ cells in the hinge region (brackets) show no apoptosis (C). Some Wg-lacZ cells (white arrows) are apoptotic while others (yellow arrows) are not in the pouch (C). Most pouch cells that do not express Wg-lacZ (blue arrows) are apoptotic (C). Basally located Wg-lacZ cells in the hinge region are increased in their number, and were dispersed into the pouch region (brackets in D’–D’’’). (E,E’,F,F’) Abnormal Wg pattern induced by the knockdown of Dhc64C is rescued by p35. Compared to Wg pattern of nub > GFP, p35 wing discs (E), that of nub > GFP; p35, Dhc64Ci wing (F) are almost normal unlike Figure 4B.
3.7. Knockdown of Dhc64C Does Not Induce Cell Death but Induces Overproliferation of Wg-Expressing Cells in the Inner Hinge
We further analyzed the regional difference in apoptosis of Wg-lacZ cells of nub > wg-lacZ; Dhc64Ci wing discs. In the apical region, most Wg-lacZ cells near the DV midline, as well as surrounding pouch cells, contained CC3 (white and blue arrows in Figure 5C’–C”’). However, the Wg-lacZ cells in the inner hinge did not have CC3 (brackets in Figure 5C’–C”’) and even became somewhat thicker (arrows in Figure 5A,B) although Dhc64C RNAi in that region is also driven by nub-Gal4. Similarly, the Wg-lacZ cells in the inner hinge of the basal region had no CC3 signal but instead were noticeably increased in number and dispersed into the pouch (bracket in Figure 5D; Supplementary Figure S4). Therefore, the Wg-lacZ cells in the inner hinge were especially resistant to apoptosis, unlike those in the pouch. The movement of these hinge cells toward the wing pouch has been also reported when the wing pouch is damaged by cell death [33,34].
Consistent with the phenotypic rescue of nub > Dhc64Ci flies by the coexpression of p35 (Figure 3C,I), p35 also rescued the distorted pattern of Wg induced by the knockdown of Dhc64C (Figure 5E,F). Therefore, cell death induced by the knockdown of Dhc64C is responsible for all phenotypes described so far, including overproliferation of Wg-lacZ cells in the inner hinge.
4. Discussion
We report here that Dhc64Cm115 as a heterozygote is isolated as a suppressor of Sona-induced lethality, and Dhc64C and sona have a positive genetic interaction (Figure 1 and Figure 2). The mutated site in Dhc64Cm115 protein is 5 amino acid residues away from the ATPase site in the AAA3 domain, and this mutation may compromise the MT release by reducing or abolishing the ATPase activity of the AAA3 domain [21]. Dhc64C was especially important for cell survival of the apically located Wg-producing cells in wing discs (Figure 4). Interestingly, cell death caused by knockdown of Dhc64C induced overproliferation of basally located Wg-producing cells and widespread expression of Wg in the wing pouch. These suggest that Dhc64C is required for cell survival and proper compensatory proliferation for maintenance of tissue size and shape. These properties of Dhc64C were not observed in other Dynein and Kinesin components, indicating that Dhc64C plays a unique role that is not shared by other motor components. Sona is also important for cell survival and cell proliferation, so Dhc64C and Sona may cooperate to carry out these functions.
We have shown that several sona suppressors function as dimers [13,14,15], and Arrm7 mutant protein acts as a dominant negative sona suppressor [13]. Dynein complex contains a homodimer of Dhc, so Dhc64Cm115-Dhc64C dimer in Dhc64Cm115/+ heterozygotes may act as a dominant negative and compromise the activity of Dynein but the amount of Dhc64C-Dhc64C dimer in Dhc64Cm115/+ heterozygotes may still be enough to support normal development. This explains how Dhc64Cm115/+ heterozygotes develop normally to adults, and yet suppress Sona-induced lethality. This may also explain why Sona is more efficiently suppressed in Dhc64Cm115/+ heterozygotes than by the expression of Dhc64C RNAi. Knockdown of Dhc64C by Dhc64C RNAi decreases the amount of wild-type Dhc64C, but not to the right amount for suppressing the excess amount of Sona. In addition, ectopic expression of Dhc64C RNAi by Gal4 drivers such as nub-Gal4 may not be adequate to ensure proper fly development. Further studies to compare the ability of Sona suppression of Dhc64Cm115/+ heterozygotes and Dhc64C RNAi are required. Whether Sona and Dhc64C directly interact is another interesting question to explore.
The knockdown of Dhc64C induces apoptosis, overproliferation, overall increase in the level of Wg in wing discs, and the formation of small adult wings (Figure 3 and Figure 4). These phenotypes are also induced by the knockdown of Rab5 [35]. Rab5 is involved in maturation of early endosomes to late endosomes such as multivesicular bodies (MVB) [36,37]. Sona colocalizes with Rab5 in early endosomes and is then released via exosomes to extracellular space by fusion of MVB to plasma membrane [11]. Since Rab5-containing early endosomes are transported by cytoplasmic Dynein, both Rab5 and Dynein are essential for the secretion of exosomal Sona [36,38]. Whether the knockdowns of other components in endosomal pathway show both cell death and overproliferation phenotypes is an interesting question that has yet to be explored.
Cytoplasmic Dynein helps to position Golgi complex and other organelles in the cell and transports membranous organelles such as Golgi-derived membranes, endosomes and lysosomes [39]. Interestingly, Sona and Wg are secreted both by Golgi transport and via exosomes that are derived from the endosomal pathway [11,40]. Given that the Dhc64Cm115 mutant is identified as a sona suppressor, Dhc64C may play a role in transporting Sona, and thereby affects Wg signaling. Further analysis is necessary to find out the biochemical relationship between Dhc64C, Sona and Wg.
One important finding is that the knockdown of Dhc64C causes cell death of the Wg-expressing cells in the apical region, while causing cell proliferation of the Wg-expressing cells in the basal region of wing discs (Figure 4 and Figure 5). Since hinge cells also proliferate and move to the pouch region when pouch cells become apoptotic under stress conditions [33,34], these Wg-lacZ cells in the basal region are most likely originated from the hinge, moving to the basal region of wing discs to repair the site emptied by cell death. Therefore, the apically located Wg-lacZ cells are the original residents of the disc proper of the wing pouch, while the basally located Wg-lacZ cells are the descendants of the hinge cells originated from peripodial epithelium. It is essential to understand how Dhc64C and Sona function in these hinge cells for cell survival and proliferation.
5. Conclusions
We identified Dhc64Cm115 as a suppressor of Sona-induced lethality. Dhc64C is important for cell survival, especially apical Wg-producing cells in wing discs. Therefore, Dhc64C is involved in Wg signaling by apical transport of wg transcript, and also in cell survival of Wg-producing cells. Since Sona is also important for cell survival, we propose that Dhc64C and Sona cooperate to promote cell survival by positively regulating Wg signaling.
Supplementary Materials
The following are available online at www.mdpi.com/article/10.3390/jdb9040043/s1, Table S1: Meiotic mapping of the m115 suppressor; Table S2: Ten potential genes for m115 identified by deficiency mapping; Table S3: The knockdown phenotypes of the two Dhc64C RNAi lines are identical; Figure S1: Dhc64C is important for cell survival; Figure S2: Knockdown of Dhc64C reduces the expression level of Sens, which is partially rescued by coexpression of p35; Figure S3: Subunits in Dynein and Kinesin microtubule motors do not induce overproliferation of Wg-producing cells; Figure S4: The Wg-lacZ cells in the apical region are apoptotic unlike the Wg-lacZ cells in the basal region.
Author Contributions
J.-Y.K., O.T. and K.-O.C. designed experiments, analyzed data, and wrote the paper. J.-Y.K. and O.T. conducted experiments. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research Foundation of Korea (NRF) funded by the Ministry of Education, 2017R1A2B4009254 and 2019R1H1A2039726.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The materials described in the manuscript, including all relevant raw data, will be freely available to any researcher wishing to use them for non-commercial purposes, without breaching participant confidentiality. The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We are grateful to colleagues in our lab and Kwang Choi’s lab for discussion and comments on this paper. We thank Bloomington Drosophila Stock Center, Vienna Drosophila Resource Center, National Institute of Genetics, and Developmental Studies Hybridoma Bank for antibodies and fly lines. This research was supported by the National Research Foundation of Korea (NRF) funded by the Ministry of Education, 2017R1A2B4009254 and 2019R1H1A2039726.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript, or in the decision to publish the results.
References
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, H.D.; Gorenc, T.; Steller, H. Apoptotic Cells Can Induce Compensatory Cell Proliferation through the JNK and the Wingless Signaling Pathways. Dev. Cell 2004, 7, 491–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Baehrecke, E.H.; Thummel, C.S. Steroid regulated programmed cell death during Drosophila metamorphosis. Development 1997, 124, 4673–4683. [Google Scholar] [CrossRef] [PubMed]
- Pazdera, T.M.; Janardhan, P.; Minden, J.S. Patterned epidermal cell death in wild-type and segment polarity mutant Drosophila embryos. Development 1998, 125, 3427–3436. [Google Scholar] [CrossRef] [PubMed]
- Wolff, T.; Ready, D.F. Cell death in normal and rough eye mutants of Drosophila. Development 1991, 113, 825–839. [Google Scholar] [CrossRef]
- Karpen, G.H.; Schubiger, G. Extensive regulatory capabilities of a Drosophila imaginal disk blastema. Nature 1981, 294, 744–747. [Google Scholar] [CrossRef]
- El Hour, M.; Moncada-Pazos, A.; Blacher, S.; Masset, A.; Cal, S.; Berndt, S.; Detilleux, J.; Host, L.; Obaya, A.J.; Maillard, C.; et al. Higher sensitivity of Adamts12-deficient mice to tumor growth and angiogenesis. Oncogene 2010, 29, 3025–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, P.H.Y.; Lung, H.L.; Cheung, A.K.L.; Apte, S.S.; Chan, K.W.; Kwong, F.M.; Ko, J.M.Y.; Cjeng, Y.; Law, S.; Srivastava, G.; et al. Extracellular protease ADAMTS9 suppresses esophageal and nasopharyngeal carcinoma tumor formation by inhibiting angiogenesis. Cancer Res. 2010, 70, 5567–5576. [Google Scholar] [CrossRef] [Green Version]
- Moncada-Pazos, A.; Obaya, A.J.; Fraga, M.F.; Viloria, C.G.; Capellá, G.; Gausachs, M.; Esteller, M.; López-Otín, C.; Cal, S. The ADAMTS12 metalloprotease gene is epigenetically silenced in tumor cells and transcriptionally activated in the stroma during progression of colon cancer. J. Cell Sci. 2009, 122, 2906–2913. [Google Scholar] [CrossRef] [Green Version]
- Rocks, N.; Paulissen, G.; El Hour, M.; Quesada, F.; Crahay, C.; Gueders, M.; Foidart, J.; Noel, A.; Cataldo, D. Emerging roles of ADAM and ADAMTS metalloproteinases in cancer. Biochimie 2008, 90, 369–379. [Google Scholar] [CrossRef]
- Kim, G.-W.; Won, J.-H.; Lee, O.-K.; Lee, S.-S.; Han, J.-H.; Tsogtbaatar, O.; Nam, S.; Kim, Y.; Cho, K.-O. Sol narae (Sona) is a Drosophila ADAMTS involved in Wg signaling. Sci. Rep. 2016, 6, 31863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsogtbaatar, O.; Won, J.H.; Kim, G.W.; Han, J.H.; Bae, Y.K.; Cho, K.O. An ADAMTS Sol narae is required for cell survival in Drosophila. Sci. Rep. 2019, 9, 1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.-H.; Kim, Y.; Cho, K.-O. Exosomal arrow (Arr)/lipoprotein receptor protein 6 (LRP6) in Drosophila melanogaster increases the extracellular level of Sol narae (Sona) in a Wnt-independent manner. Cell Death Dis. 2020, 11, 944. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Cho, K.-O. POU domain motif3 (Pdm3) induces wingless (wg) transcription and is essential for development of larval neuromuscular junctions in Drosophila. Sci. Rep. 2020, 10, 517. [Google Scholar] [CrossRef] [Green Version]
- Nam, S.; Cho, K.-O. Wingless and Archipelago, a fly E3 ubiquitin ligase and a homolog of human tumor suppressor FBW7, show an antagonistic relationship in wing development. BMC Dev. Biol. 2020, 20, 14. [Google Scholar] [CrossRef]
- Cho, D.-G.; Lee, S.-S.; Cho, A.K.-O. Anastral Spindle 3/Rotatin Stabilizes Sol narae and Promotes Cell Survival in Drosophila melanogaster. Mol. Cells 2021, 44, 13–25. [Google Scholar] [CrossRef]
- Asai, D.J.; Koonce, M.P. The dynein heavy chain: Structure, mechanics and evolution. Trends Cell Biol. 2001, 11, 196–202. [Google Scholar] [CrossRef]
- Asai, D.J.; Wilkes, D.E. The dynein heavy chain family. J. Eukaryot. Microbiol. 2004, 51, 23–29. [Google Scholar] [CrossRef]
- McLean, I.W.; Nakane, P.K. Periodate-lysine-paraformaldehyde fixative. A new fixation for immunoelectron microscopy. J. Histochem. Cytochem. 1974, 22, 1077–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.-O.; Chern, J.; Izaddoost, S.; Choi, K.-W. Novel Signaling from the Peripodial Membrane Is Essential for Eye Disc Patterning in Drosophila. Cell 2000, 103, 331–342. [Google Scholar] [CrossRef] [Green Version]
- DeWitt, M.A.; Cypranowska, C.A.; Cleary, F.B.; Belyy, V.; Yildiz, A. The AAA3 domain of cytoplasmic dynein acts as a switch to facilitate microtubule release. Nat. Struct. Mol. Biol. 2015, 22, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Bitan, A.; Guild, G.M.; Bar-Dubin, D.; Abdu, U. Asymmetric Microtubule Function Is an Essential Requirement for Polarized Organization of the Drosophila Bristle. Mol. Cell. Biol. 2010, 30, 496–507. [Google Scholar] [CrossRef] [Green Version]
- Jack, J.; Dorsett, D.; DeLotto, Y.; Liu, S. Expression of the cut locus in the Drosophila wing margin is required for cell type specification and is regulated by a distant enhancer. Development 1991, 113, 735–747. [Google Scholar] [CrossRef]
- Seto, E.S.; Bellen, H.J. Internalization is required for proper Wingless signaling in Drosophila melanogaster. J. Cell Biol. 2006, 173, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Won, J.-H.; Kim, G.-W.; Kim, J.-Y.; Cho, D.-G.; Kwon, B.; Bae, Y.-K.; Cho, K.-O. ADAMTS Sol narae cleaves extracellular Wingless to generate a novel active form that regulates cell proliferation in Drosophila. Cell Death Dis. 2019, 10, 564. [Google Scholar] [CrossRef] [Green Version]
- Garijo, A.P.; Shlevkov, E.; Morata, G. The role of Dpp and Wg in compensatory proliferation and in the formation of hyperplastic overgrowths caused by apoptotic cells in the Drosophila wing disc. Development 2009, 136, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Batlevi, Y.; Martin, D.N.; Pandey, U.B.; Simon, C.R.; Powers, C.M.; Taylor, J.P.; Baehrecke, E.H. Dynein light chain 1 is required for autophagy, protein clearance, and cell death in Drosophila. Proc. Natl. Acad. Sci. USA 2010, 107, 742–747. [Google Scholar] [CrossRef] [Green Version]
- Dick, T.; Ray, K.; Salz, H.K.; Chia, W. Cytoplasmic dynein (ddlc1) mutations cause morphogenetic defects and apoptotic cell death in Drosophila melanogaster. Mol. Cell. Biol. 1996, 16, 1966–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolly, A.L.; Kim, H.; Srinivasan, D.; Lakonishok, M.; Larson, A.G.; Gelfand, V.I. Kinesin-1 heavy chain mediates microtubule sliding to drive changes in cell shape. Proc. Natl. Acad. Sci. USA 2010, 107, 12151–12156. [Google Scholar] [CrossRef] [Green Version]
- Saxton, W.M.; Hicks, J.; Goldstein, L.S.; Raff, E.C. Kinesin heavy chain is essential for viability and neuromuscular functions in Drosophila, but mutants show no defects in mitosis. Cell 1991, 64, 1093–1102. [Google Scholar] [CrossRef]
- Mukhopadhyay, B.; Nam, S.-C.; Choi, K.-W. Kinesin II is required for cell survival and adherens junction positioning in Drosophila photoreceptors. Genesis 2010, 48, 522–530. [Google Scholar] [CrossRef]
- Ray, K.; Perez, S.E.; Yang, Z.; Xu, J.; Ritchings, B.W.; Steller, H.; Goldstein, L.S. Kinesin-II Is Required for Axonal Transport of Choline Acetyltransferase in Drosophila. J. Cell Biol. 1999, 147, 507–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, S.C.; Martín, R.; Morata, G. Tissue Homeostasis in the Wing Disc of Drosophila melanogaster: Immediate Response to Massive Damage during Development. PLoS Genet. 2013, 9, e1003446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verghese, S.; Su, T.T. Drosophila Wnt and STAT Define Apoptosis-Resistant Epithelial Cells for Tissue Regeneration after Irradiation. PLoS Biol. 2016, 14, e1002536. [Google Scholar] [CrossRef]
- Ballesteros-Arias, L.; Saavedra, V.; Morata, G. Cell competition may function either as tumour-suppressing or as tumour-stimulating factor in Drosophila. Oncogene 2014, 33, 4377–4384. [Google Scholar] [CrossRef] [Green Version]
- Marois, E.; Mahmoud, A.; Eaton, S. The endocytic pathway and formation of the Wingless morphogen gradient. Development 2006, 133, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Satoh, D.; Sato, D.; Tsuyama, T.; Saito, M.; Ohkura, H.; Rolls, M.; Ishikawa, F.; Uemura, T. Spatial control of branching within dendritic arbors by dynein-dependent transport of Rab5-endosomes. Nature 2008, 10, 1164–1171. [Google Scholar] [CrossRef]
- Lin, S.X.; A Collins, C. Immunolocalization of cytoplasmic dynein to lysosomes in cultured cells. J. Cell Sci. 1992, 101, 125–137. [Google Scholar] [CrossRef]
- Gross, J.C.; Chaudhary, V.; Bartscherer, K.; Boutros, M. Active Wnt proteins are secreted on exosomes. Nature 2012, 14, 1036–1045. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).