
Expanding the Junction: New Insights into Non-Occluding Roles for Septate Junction Proteins during Development


Abstract
1. Introduction
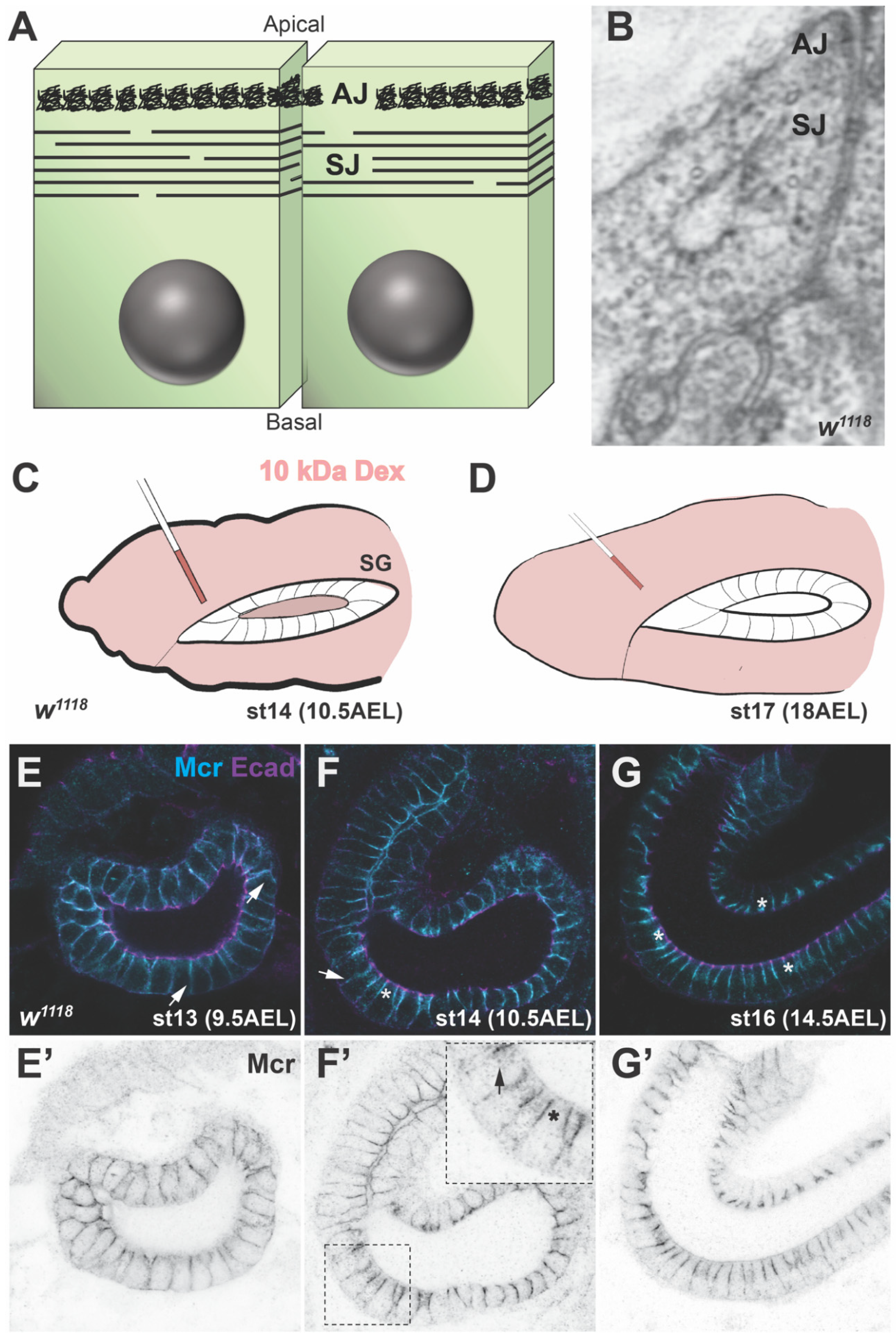
2. Structure and Composition of SJs
3. Biogenesis of the SJ
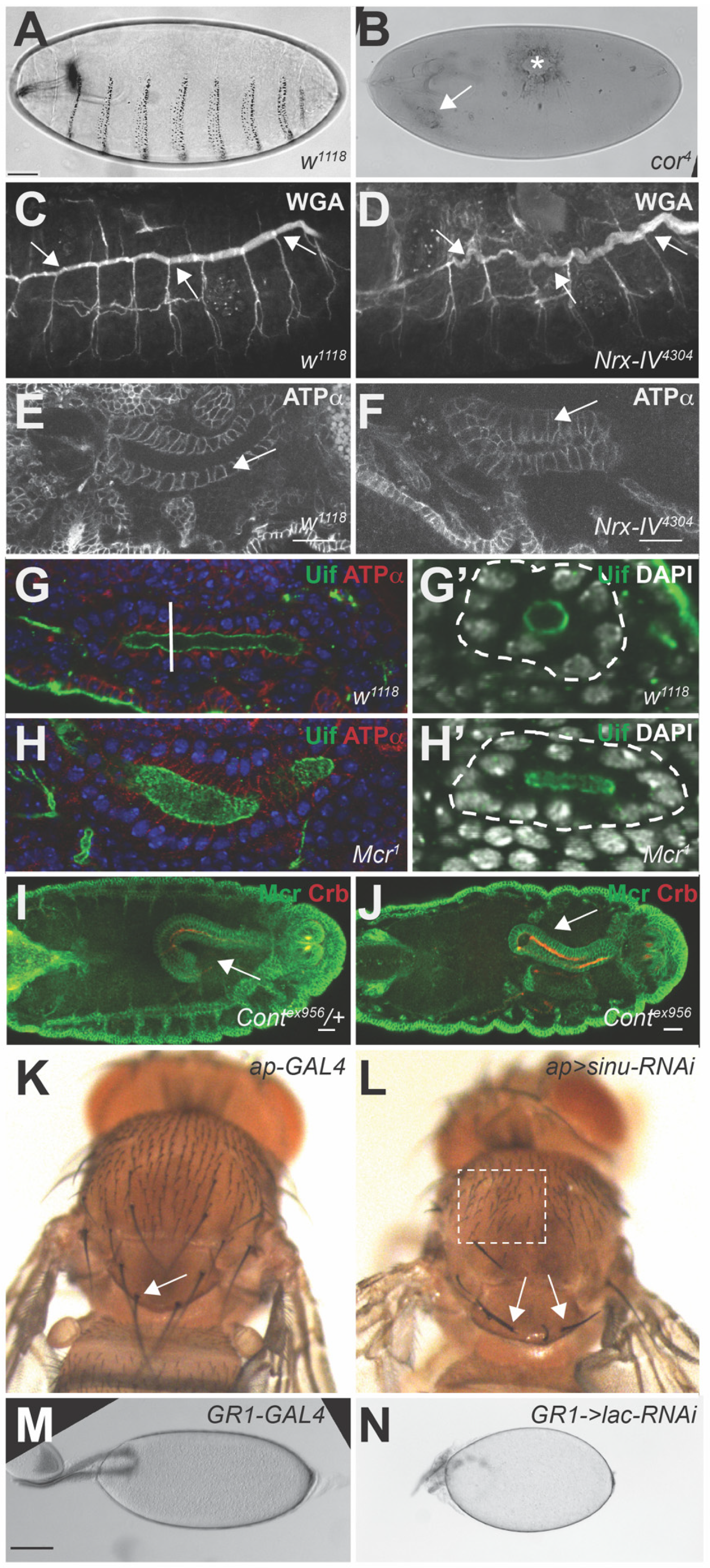
4. Developmental Functions of SJ Genes
4.1. Dorsal Closure (DC) and Head Involution (HI)
4.2. Organogenesis of the Trachea, Salivary Glands and Hindgut
4.3. Dorsal Vessel Development
4.4. Imaginal Disc Morphogenesis
4.5. Morphogenetic Events during Oogenesis
4.6. Hemocyte Encapsulation of Parasitic Wasp Eggs
5. Cellular Functions of SJ Genes
5.1. Biomechanical Tissue Regulation during Wound Healing
5.2. Planar Cell Polarity in Imaginal Epithelia
5.3. Apical-Basal Polarity in the Embryonic Epidermis
5.4. Lateral Membrane Adhesion
5.5. Polarized Secretion?
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lord, B.A.; DiBona, D.R. Role of the septate junction in the regulation of paracellular transepithelial flow. J. Cell Biol. 1976, 71, 967–972. [Google Scholar] [CrossRef]
- Furuse, M.; Izumi, Y. Molecular dissection of smooth septate junctions: Understanding their roles in arthropod physiology. Ann. N. Y. Acad. Sci. 2017, 1397, 17–24. [Google Scholar] [CrossRef]
- Noirot-Timothee, C.; Smith, D.S.; Cayer, M.L.; Noirot, C. Septate junctions in insects: Comparison between intercellular and intramembranous structures. Tissue Cell 1978, 10, 125–136. [Google Scholar] [CrossRef]
- Poodry, C.; Schneiderman, H. The ultrastructure of the developing leg of Drosophila melanogaster. Roux Archiv 1970, 166, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Tepass, U.; Hartenstein, V. The development of cellular junctions in the Drosophila embryo. Dev. Biol. 1994, 161, 563–596. [Google Scholar] [CrossRef]
- Mahowald, A.P. Ultrastructural observations on oogenesis in Drosophila. J. Morphol. 1972, 137, 29–48. [Google Scholar] [CrossRef]
- Fairchild, M.J.; Smendziuk, C.M.; Tanentzapf, G. A somatic permeability barrier around the germline is essential for Drosophila spermatogenesis. Development 2015, 142, 268–281. [Google Scholar] [CrossRef]
- Russo, J.; Dupas, S.; Frey, F.; Carton, Y.; Brehelin, M. Insect immunity: Early events in the encapsulation process of parasitoid (Leptopilina boulardi) eggs in resistant and susceptible strains of Drosophila. Parasitology 1996, 112 Pt 1, 135–142. [Google Scholar] [CrossRef]
- Flower, N.E.; Filshie, B.K. Junctional structures in the midgut cells of lepidopteran caterpillars. J. Cell Sci. 1975, 17, 221–239. [Google Scholar] [PubMed]
- Schulte, J.; Tepass, U.; Auld, V.J. Gliotactin, a novel marker of tricellular junctions, is necessary for septate junction development in Drosophila. J. Cell Biol. 2003, 161, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Byri, S.; Misra, T.; Syed, Z.A.; Batz, T.; Shah, J.; Boril, L.; Glashauser, J.; Aegerter-Wilmsen, T.; Matzat, T.; Moussian, B.; et al. The triple-repeat protein anakonda controls epithelial tricellular junction formation in Drosophila. Dev. Cell 2015, 33, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, A.; Pflanz, R.; Behr, M.; Tarp, T.; Riedel, D.; Schuh, R. Bark beetle controls epithelial morphogenesis by septate junction maturation in Drosophila. Dev. Biol. 2015, 400, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Zappia, M.P.; Brocco, M.A.; Billi, S.C.; Frasch, A.C.; Ceriani, M.F. M6 membrane protein plays an essential role in Drosophila oogenesis. PLoS ONE 2011, 6, e19715. [Google Scholar] [CrossRef] [PubMed]
- Higashi, T.; Chiba, H. Molecular organization, regulation and function of tricellular junctions. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183143. [Google Scholar] [CrossRef]
- Esmangart de Bournonville, T.; Le Borgne, R. Interplay between anakonda, gliotactin, and M6 for tricellular junction assembly and anchoring of septate junctions in Drosophila epithelium. Curr. Biol. 2020, 30, 4245–4253.e4244. [Google Scholar] [CrossRef] [PubMed]
- Wittek, A.; Hollmann, M.; Schleutker, R.; Luschnig, S. The transmembrane proteins M6 and anakonda cooperate to initiate tricellular junction assembly in epithelia of Drosophila. Curr. Biol. 2020, 30, 4254–4262.e4255. [Google Scholar] [CrossRef]
- Urakabe, S.; Handler, J.S.; Orloff, J. Effect of hypertonicity on permeability properties of the toad bladder. Am. J. Physiol. 1970, 218, 1179–1187. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lamb, R.S.; Ward, R.E.; Schweizer, L.; Fehon, R.G. Drosophila coracle, a member of the protein 4.1 superfamily, has essential structural functions in the septate junctions and developmental functions in embryonic and adult epithelial cells. Mol. Biol. Cell 1998, 9, 3505–3519. [Google Scholar] [CrossRef]
- Fairchild, M.J.; Yang, L.; Goodwin, K.; Tanentzapf, G. Occluding junctions maintain stem cell niche homeostasis in the fly testes. Curr. Biol. 2016, 26, 2492–2499. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, S.; Littleton, J.T.; Broadie, K.; Bhat, M.A.; Harbecke, R.; Lengyel, J.A.; Chiquet-Ehrismann, R.; Prokop, A.; Bellen, H.J. A Drosophila neurexin is required for septate junction and blood-nerve barrier formation and function. Cell 1996, 87, 1059–1068. [Google Scholar] [CrossRef]
- Genova, J.L.; Fehon, R.G. Neuroglian, gliotactin, and the Na+/K+ ATPase are essential for septate junction function in Drosophila. J. Cell Biol. 2003, 161, 979–989. [Google Scholar] [CrossRef]
- Batz, T.; Forster, D.; Luschnig, S. The transmembrane protein macroglobulin complement-related is essential for septate junction formation and epithelial barrier function in Drosophila. Development 2014, 141, 899–908. [Google Scholar] [CrossRef]
- Hall, S.; Bone, C.; Oshima, K.; Zhang, L.; McGraw, M.; Lucas, B.; Fehon, R.G.; Ward, R.E. Macroglobulin complement-related (MCR) encodes a protein required for septate junction organization and paracellular barrier function in Drosophila. Development 2014, 141, 889–898. [Google Scholar] [CrossRef]
- Wu, V.M.; Schulte, J.; Hirschi, A.; Tepass, U.; Beitel, G.J. Sinuous is a Drosophila claudin required for septate junction organization and epithelial tube size control. J. Cell Biol. 2004, 164, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.S.; Furuse, M.; Beitel, G.J. The Drosophila claudin kune-kune is required for septate junction organization and tracheal tube size control. Genetics 2010, 185, 831–839. [Google Scholar] [CrossRef]
- Faivre-Sarrailh, C.; Banerjee, S.; Li, J.; Hortsch, M.; Laval, M.; Bhat, M.A. Drosophila contactin, a homolog of vertebrate contactin, is required for septate junction organization and paracellular barrier function. Development 2004, 131, 4931–4942. [Google Scholar] [CrossRef]
- Fehon, R.G.; Dawson, I.A.; Artavanis-Tsakonas, S. A Drosophila homologue of membrane-skeleton protein 4.1 is associated with septate junctions and is encoded by the coracle gene. Development 1994, 120, 545–557. [Google Scholar] [PubMed]
- Wu, V.M.; Yu, M.H.; Paik, R.; Banerjee, S.; Liang, Z.; Paul, S.M.; Bhat, M.A.; Beitel, G.J. Drosophila varicose, a member of a new subgroup of basolateral MAGUKs, is required for septate junctions and tracheal morphogenesis. Development 2007, 134, 999–1009. [Google Scholar] [CrossRef]
- Woods, D.F.; Bryant, P.J. The discs-large tumor suppressor gene of Drosophila encodes a guanylate kinase homolog localized at septate junctions. Cell 1991, 66, 451–464. [Google Scholar] [CrossRef]
- Bilder, D.; Perrimon, N. Localization of apical epithelial determinants by the basolateral PDZ protein Scribble. Nature 2000, 403, 676–680. [Google Scholar] [CrossRef]
- Laprise, P.; Paul, S.M.; Boulanger, J.; Robbins, R.M.; Beitel, G.J.; Tepass, U. Epithelial polarity proteins regulate Drosophila tracheal tube size in parallel to the luminal matrix pathway. Curr. Biol. 2010, 20, 55–61. [Google Scholar] [CrossRef]
- Hijazi, A.; Haenlin, M.; Waltzer, L.; Roch, F. The Ly6 protein coiled is required for septate junction and blood brain barrier organisation in Drosophila. PLoS ONE 2011, 6, e17763. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, A.; Masson, W.; Auge, B.; Waltzer, L.; Haenlin, M.; Roch, F. boudin is required for septate junction organisation in Drosophila and codes for a diffusible protein of the Ly6 superfamily. Development 2009, 136, 2199–2209. [Google Scholar] [CrossRef]
- Nilton, A.; Oshima, K.; Zare, F.; Byri, S.; Nannmark, U.; Nyberg, K.G.; Fehon, R.G.; Uv, A.E. Crooked, coiled and crimpled are three Ly6-like proteins required for proper localization of septate junction components. Development 2010, 137, 2427–2437. [Google Scholar] [CrossRef]
- Syed, M.H.; Krudewig, A.; Engelen, D.; Stork, T.; Klambt, C. The CD59 family member leaky/coiled is required for the establishment of the blood-brain barrier in Drosophila. J. Neurol. Sci. 2011, 31, 7876–7885. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ile, K.E.; Tripathy, R.; Goldfinger, V.; Renault, A.D. Wunen, a Drosophila lipid phosphate phosphatase, is required for septate junction-mediated barrier function. Development 2012, 139, 2535–2546. [Google Scholar] [CrossRef] [PubMed]
- Tiklova, K.; Senti, K.A.; Wang, S.; Graslund, A.; Samakovlis, C. Epithelial septate junction assembly relies on melanotransferrin iron binding and endocytosis in Drosophila. Nat. Cell Biol. 2010, 12, 1071–1077. [Google Scholar] [CrossRef]
- Paul, S.M.; Ternet, M.; Salvaterra, P.M.; Beitel, G.J. The Na+/K+ ATPase is required for septate junction function and epithelial tube-size control in the Drosophila tracheal system. Development 2003, 130, 4963–4974. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Paik, R.; Mino, R.E.; Blauth, K.; Fisher, E.S.; Madden, V.J.; Fanning, A.S.; Bhat, M.A. A laminin G-EGF-laminin G module in neurexin IV is essential for the apico-lateral localization of contactin and organization of septate junctions. PLoS ONE 2011, 6, e25926. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, M.H.; Nolde, K.; Behr, M.; Joo, S.H.; Plessmann, U.; Nikolov, M.; Urlaub, H.; Schuh, R. The claudin Megatrachea protein complex. J. Biol. Chem. 2012, 287, 36756–36765. [Google Scholar] [CrossRef]
- Llimargas, M.; Strigini, M.; Katidou, M.; Karagogeos, D.; Casanova, J. Lachesin is a component of a septate junction-based mechanism that controls tube size and epithelial integrity in the Drosophila tracheal system. Development 2004, 131, 181–190. [Google Scholar] [CrossRef]
- Behr, M.; Riedel, D.; Schuh, R. The claudin-like megatrachea is essential in septate junctions for the epithelial barrier function in Drosophila. Dev. Cell 2003, 5, 611–620. [Google Scholar] [CrossRef]
- Deligiannaki, M.; Casper, A.L.; Jung, C.; Gaul, U. Pasiflora proteins are novel core components of the septate junction. Development 2015, 142, 3046–3057. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, A.; Draga, M.; Grawe, F.; Knust, E. On the role of the MAGUK proteins encoded by Drosophila varicose during embryonic and postembryonic development. BMC Dev. Biol. 2008, 8, 55. [Google Scholar] [CrossRef]
- Moyer, K.E.; Jacobs, J.R. Varicose: A MAGUK required for the maturation and function of Drosophila septate junctions. BMC Dev. Biol. 2008, 8, 99. [Google Scholar] [CrossRef]
- Konigsmann, T.; Parfentev, I.; Urlaub, H.; Riedel, D.; Schuh, R. The bicistronic gene wurmchen encodes two essential components for epithelial development in Drosophila. Dev. Biol. 2020, 463, 53–62. [Google Scholar] [CrossRef]
- Woods, D.F.; Bryant, P.J. ZO-1, DlgA and PSD-95/SAP90: Homologous proteins in tight, septate and synaptic cell junctions. Mech. Dev. 1993, 44, 85–89. [Google Scholar] [CrossRef]
- Bilder, D. Epithelial polarity and proliferation control: Links from the Drosophila neoplastic tumor suppressors. Genes Dev. 2004, 18, 1909–1925. [Google Scholar] [CrossRef]
- Oshima, K.; Fehon, R.G. Analysis of protein dynamics within the septate junction reveals a highly stable core protein complex that does not include the basolateral polarity protein Discs large. J. Cell Sci. 2011, 124 Pt 16, 2861–2871. [Google Scholar] [CrossRef]
- Paul, S.M.; Palladino, M.J.; Beitel, G.J. A pump-independent function of the Na,K-ATPase is required for epithelial junction function and tracheal tube-size control. Development 2007, 134, 147–155. [Google Scholar] [CrossRef]
- Gunzel, D.; Yu, A.S. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef]
- Ward, R.E.; Lamb, R.S.; Fehon, R.G. A conserved functional domain of Drosophila coracle is required for localization at the septate junction and has membrane-organizing activity. J. Cell Biol. 1998, 140, 1463–1473. [Google Scholar] [CrossRef]
- Hall, S.; Ward, R.E. Septate junction proteins play essential roles in morphogenesis throughout embryonic development in Drosophila. G3 Bethesda 2016, 6, 2375–2384. [Google Scholar] [CrossRef]
- Laval, M.; Bel, C.; Faivre-Sarrailh, C. The lateral mobility of cell adhesion molecules is highly restricted at septate junctions in Drosophila. BMC Cell Biol. 2008, 9, 38. [Google Scholar] [CrossRef]
- Alhadyian, H.; Shoaib, D.; Ward, R.E. Septate junction proteins are required for egg elongation and border cell migration during oogenesis in Drosophila. G3 2021, in press. [Google Scholar]
- Muller, H.A. Genetic control of epithelial cell polarity: Lessons from Drosophila. Dev. Dyn. 2000, 218, 52–67. [Google Scholar] [CrossRef]
- Swanson, L.E.; Yu, M.; Nelson, K.S.; Laprise, P.; Tepass, U.; Beitel, G.J. Drosophila convoluted/dALS is an essential gene required for tracheal tube morphogenesis and apical matrix organization. Genetics 2009, 181, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- VanHook, A.; Letsou, A. Head involution in Drosophila: Genetic and morphogenetic connections to dorsal closure. Dev. Dyn. 2008, 237, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, B.G.; Arias, A.M.; Jacinto, A. Dpp signalling orchestrates dorsal closure by regulating cell shape changes both in the amnioserosa and in the epidermis. Mech. Dev. 2007, 124, 884–897. [Google Scholar] [CrossRef]
- Franke, J.D.; Montague, R.A.; Kiehart, D.P. Nonmuscle myosin II generates forces that transmit tension and drive contraction in multiple tissues during dorsal closure. Curr. Biol. 2005, 15, 2208–2221. [Google Scholar] [CrossRef]
- Kiehart, D.P.; Galbraith, C.G.; Edwards, K.A.; Rickoll, W.L.; Montague, R.A. Multiple forces contribute to cell sheet morphogenesis for dorsal closure in Drosophila. J. Cell Biol. 2000, 149, 471–490. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.E.; Schweizer, L.; Lamb, R.S.; Fehon, R.G. The protein 4.1, ezrin, radixin, moesin (FERM) domain of Drosophila coracle, a cytoplasmic component of the septate junction, provides functions essential for embryonic development and imaginal cell proliferation. Genetics 2001, 159, 219–228. [Google Scholar] [PubMed]
- Maruyama, R.; Andrew, D.J. Drosophila as a model for epithelial tube formation. Dev. Dyn. 2012, 241, 119–135. [Google Scholar] [CrossRef]
- Luschnig, S.; Batz, T.; Armbruster, K.; Krasnow, M.A. serpentine and vermiform encode matrix proteins with chitin binding and deacetylation domains that limit tracheal tube length in Drosophila. Curr. Biol. 2006, 16, 186–194. [Google Scholar] [CrossRef]
- Wang, S.; Jayaram, S.A.; Hemphala, J.; Senti, K.A.; Tsarouhas, V.; Jin, H.; Samakovlis, C. Septate-junction-dependent luminal deposition of chitin deacetylases restricts tube elongation in the Drosophila trachea. Curr. Biol. 2006, 16, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.S.; Khan, Z.; Molnar, I.; Mihaly, J.; Kaschube, M.; Beitel, G.J. Drosophila Src regulates anisotropic apical surface growth to control epithelial tube size. Nat. Cell Biol. 2012, 14, 518–525. [Google Scholar] [CrossRef]
- Robbins, R.M.; Gbur, S.C.; Beitel, G.J. Non-canonical roles for Yorkie and Drosophila inhibitor of apoptosis 1 in epithelial tube size control. PLoS ONE 2014, 9, e101609. [Google Scholar] [CrossRef]
- Laprise, P.; Lau, K.M.; Harris, K.P.; Silva-Gagliardi, N.F.; Paul, S.M.; Beronja, S.; Beitel, G.J.; McGlade, C.J.; Tepass, U. Yurt, Coracle, Neurexin IV and the Na+,K+-ATPase form a novel group of epithelial polarity proteins. Nature 2009, 459, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.L.; Myat, M.M.; Comeaux, C.A.; Andrew, D.J. Posterior migration of the salivary gland requires an intact visceral mesoderm and integrin function. Dev. Biol. 2003, 257, 249–262. [Google Scholar] [CrossRef]
- Vining, M.S.; Bradley, P.L.; Comeaux, C.A.; Andrew, D.J. Organ positioning in Drosophila requires complex tissue-tissue interactions. Dev. Biol. 2005, 287, 19–34. [Google Scholar] [CrossRef]
- Xu, N.; Bagumian, G.; Galiano, M.; Myat, M.M. Rho GTPase controls Drosophila salivary gland lumen size through regulation of the actin cytoskeleton and moesin. Development 2011, 138, 5415–5427. [Google Scholar] [CrossRef]
- Wells, R.E.; Barry, J.D.; Warrington, S.J.; Cuhlmann, S.; Evans, P.; Huber, W.; Strutt, D.; Zeidler, M.P. Control of tissue morphology by fasciclin III-mediated intercellular adhesion. Development 2013, 140, 3858–3868. [Google Scholar] [CrossRef] [PubMed]
- Snow, P.M.; Bieber, A.J.; Goodman, C.S. Fasciclin III: A novel homophilic adhesion molecule in Drosophila. Cell 1989, 59, 313–323. [Google Scholar] [CrossRef]
- Rugendorff, A.; Younossi-Hartenstein, A.; Hartenstein, V. Embryonic origin and differentiation of the Drosophila heart. Rouxs Arch. Dev. Biol. 1994, 203, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Cripps, R.M.; Olson, E.N. Control of cardiac development by an evolutionarily conserved transcriptional network. Dev. Biol. 2002, 246, 14–28. [Google Scholar] [CrossRef]
- Das, D.; Aradhya, R.; Ashoka, D.; Inamdar, M. Macromolecular uptake in Drosophila pericardial cells requires rudhira function. Exp. Cell Res. 2008, 314, 1804–1810. [Google Scholar] [CrossRef]
- Lim, H.Y.; Bao, H.; Liu, Y.; Wang, W. Select septate junction proteins direct ROS-mediated paracrine regulation of Drosophila cardiac function. Cell Rep. 2019, 28, 1455–1470.e1454. [Google Scholar] [CrossRef]
- Mills, R.P.; King, R.C. The pericardial cells of Drosophila melanogaster. Q. J. Microsc. Sci. 1965, 106, 261–268. [Google Scholar]
- Wigglesworth, V.B. The pericardial cells of insects: Analogue of the reticuloendothelial system. J. Reticuloendothel. Soc. 1970, 7, 208–216. [Google Scholar] [PubMed]
- Yi, P.; Johnson, A.N.; Han, Z.; Wu, J.; Olson, E.N. Heterotrimeric G proteins regulate a noncanonical function of septate junction proteins to maintain cardiac integrity in Drosophila. Dev. Cell 2008, 15, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Yi, P.; Han, Z.; Li, X.; Olson, E.N. The mevalonate pathway controls heart formation in Drosophila by isoprenylation of Ggamma1. Science 2006, 313, 1301–1303. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.Y.; Wang, W.; Chen, J.; Ocorr, K.; Bodmer, R. ROS regulate cardiac function via a distinct paracrine mechanism. Cell Rep. 2014, 7, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Lehmacher, C.; Togel, M.; Pass, G.; Paululat, A. The Drosophila wing hearts consist of syncytial muscle cells that resemble adult somatic muscles. Arthropod Struct. Dev. 2009, 38, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Fristrom, D.; Fristrom, J.W. The metamorphic development of the adult epidermis. In The Development of Drosophila melanogaster; Bate, M., Martinez, A.A., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1993; Volume II, pp. 843–897. [Google Scholar]
- Ward, R.E.; Evans, J.; Thummel, C.S. Genetic modifier screens in Drosophila demonstrate a role for Rho1 signaling in ecdysone-triggered imaginal disc morphogenesis. Genetics 2003, 165, 1397–1415. [Google Scholar]
- Condic, M.L.; Fristrom, D.; Fristrom, J.W. Apical cell shape changes during Drosophila imaginal leg disc elongation: A novel morphogenetic mechanism. Development 1991, 111, 23–33. [Google Scholar] [PubMed]
- Fristrom, D. The mechanism of evagination of imaginal discs of Drosophila melanogaster. III. Evidence for cell rearrangement. Dev. Biol. 1976, 54, 163–171. [Google Scholar] [CrossRef]
- Venema, D.R.; Zeev-Ben-Mordehai, T.; Auld, V.J. Transient apical polarization of Gliotactin and Coracle is required for parallel alignment of wing hairs in Drosophila. Dev. Biol. 2004, 275, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Horne-Badovinac, S.; Bilder, D. Mass transit: Epithelial morphogenesis in the Drosophila egg chamber. Dev. Dyn. 2005, 232, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Ben-Zvi, D.S.; Volk, T. Escort cell encapsulation of Drosophila germline cells is maintained by irre cell recognition module proteins. Biol. Open 2019, 8, bio039842. [Google Scholar] [CrossRef]
- Maimon, I.; Popliker, M.; Gilboa, L. Without children is required for Stat-mediated zfh1 transcription and for germline stem cell differentiation. Development 2014, 141, 2602–2610. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Khalil, A.A.; Poulton, J.; Castillejo-Lopez, C.; Egger-Adam, D.; Wodarz, A.; Deng, W.M.; Baumgartner, S. Perlecan and dystroglycan act at the basal side of the Drosophila follicular epithelium to maintain epithelial organization. Development 2006, 133, 3805–3815. [Google Scholar] [CrossRef]
- Wei, J.; Hortsch, M.; Goode, S. Neuroglian stabilizes epithelial structure during Drosophila oogenesis. Dev. Dyn. 2004, 230, 800–808. [Google Scholar] [CrossRef]
- Duhart, J.C.; Parsons, T.T.; Raftery, L.A. The repertoire of epithelial morphogenesis on display: Progressive elaboration of Drosophila egg structure. Mech Dev. 2017, 148, 18–39. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J. Drosophila hemopoiesis and cellular immunity. J. Immunol. 2007, 178, 4711–4716. [Google Scholar] [CrossRef]
- Williams, M.J. The Drosophila cell adhesion molecule Neuroglian regulates Lissencephaly-1 localisation in circulating immunosurveillance cells. BMC Immunol. 2009, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J.; Ando, I.; Hultmark, D. Drosophila melanogaster Rac2 is necessary for a proper cellular immune response. Genes Cells 2005, 10, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Wood, W.; Jacinto, A.; Grose, R.; Woolner, S.; Gale, J.; Wilson, C.; Martin, P. Wound healing recapitulates morphogenesis in Drosophila embryos. Nat. Cell Biol. 2002, 4, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Blanco, M.T.; Verboon, J.M.; Liu, R.; Watts, J.J.; Parkhurst, S.M. Drosophila embryos close epithelial wounds using a combination of cellular protrusions and an actomyosin purse string. J. Cell Sci. 2012, 125 Pt 24, 5984–5997. [Google Scholar] [CrossRef]
- Ducuing, A.; Vincent, S. The actin cable is dispensable in directing dorsal closure dynamics but neutralizes mechanical stress to prevent scarring in the Drosophila embryo. Nat. Cell Biol. 2016, 18, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Razzell, W.; Wood, W.; Martin, P. Recapitulation of morphogenetic cell shape changes enables wound re-epithelialisation. Development 2014, 141, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.; Patricio, P.; Ponte, S.; Heisenberg, C.P.; Almeida, L.; Nunes, A.S.; Araujo, N.A.M.; Jacinto, A. Occluding junctions as novel regulators of tissue mechanics during wound repair. J. Cell Biol. 2018, 217, 4267–4283. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.V.; Lee, D.M.; Harris, T.J.; Fernandez-Gonzalez, R. Polarized E-cadherin endocytosis directs actomyosin remodeling during embryonic wound repair. J. Cell Biol. 2015, 210, 801–816. [Google Scholar] [CrossRef]
- Maung, S.M.; Jenny, A. Planar cell polarity in Drosophila. Organogenesis 2011, 7, 165–179. [Google Scholar] [CrossRef]
- Fulford, A.D.; McNeill, H. Fat/dachsous family cadherins in cell and tissue organisation. Curr. Opin. Cell Biol. 2020, 62, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Vichas, A.; Zallen, J.A. Translating cell polarity into tissue elongation. Semin. Cell Dev. Biol. 2011, 22, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Tepass, U. The apical polarity protein network in Drosophila epithelial cells: Regulation of polarity, junctions, morphogenesis, cell growth, and survival. Annu Rev. Cell Dev. Biol. 2012, 28, 655–865. [Google Scholar] [CrossRef] [PubMed]
- Gamblin, C.L.; Hardy, E.J.; Chartier, F.J.; Bisson, N.; Laprise, P. A bidirectional antagonism between aPKC and Yurt regulates epithelial cell polarity. J. Cell Biol. 2014, 204, 487–495. [Google Scholar] [CrossRef]
- Laprise, P.; Beronja, S.; Silva-Gagliardi, N.F.; Pellikka, M.; Jensen, A.M.; McGlade, C.J.; Tepass, U. The FERM protein Yurt is a negative regulatory component of the Crumbs complex that controls epithelial polarity and apical membrane size. Dev. Cell 2006, 11, 363–374. [Google Scholar] [CrossRef]
- Bergstralh, D.T.; Lovegrove, H.E.; St Johnston, D. Lateral adhesion drives reintegration of misplaced cells into epithelial monolayers. Nat. Cell Biol. 2015, 17, 1497–1503. [Google Scholar] [CrossRef]
- Cammarota, C.; Finegan, T.M.; Wilson, T.J.; Yang, S.; Bergstralh, D.T. An axon-pathfinding mechanism preserves epithelial tissue integrity. Curr. Biol. 2020, 30, 5049–5057.e5043. [Google Scholar] [CrossRef]
- Nakajima, Y.; Meyer, E.J.; Kroesen, A.; McKinney, S.A.; Gibson, M.C. Epithelial junctions maintain tissue architecture by directing planar spindle orientation. Nature 2013, 500, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Rosenbluth, J. Glial membranes and axo-glial junctions. In Neuoglia; Kettenmann, H., Ransom, B.R., Eds.; Oxford University Press: New York, NY, USA, 1995; pp. 613–633. [Google Scholar]



{kind=link}
{kind=link}
| Protein | Core/Resident/Accessory | Protein Domains | References |
|---|---|---|---|
| ATPα | Core | P-type cationic transporter | [21,38] |
| Contactin | Core | C-type lectin-like, Ig, FN3 | [26,39] |
| Coracle | Core | FERM | [18,27] |
| Crimpled | Core | Ly6 | [34,40] |
| Kune-kune | Core | PMP-22/claudin | [25] |
| Lachesin | Core | Ig | [41] |
| Macroglobulin complement-related | Core | LDL Rec A, α2 macroglobulin | [22,23] |
| Megatrachea (Pickel) | Core | PMP-22/claudin | [42] |
| Nervana 2 | Core | Na/K ATPase β subunit | [38] |
| Neurexin-IV | Core | FA58C, LamG, EGF-like | [20,39] |
| Neuroglian | Core | Ig, FN3 | [21] |
| Pasiflora 1 | Core | None (4 TM) | [43] |
| Pasiflora 2 | Core | None (4 TM) | [43] |
| Sinuous | Core | PMP-22/claudin | [24] |
| Transferrin 2 (Melanotransferrin) | Core | Transferrin-like | [37] |
| Varicose | Core | PDZ, SH3, GUK | [44,45] |
| Würmchen | Core | None | [46] |
| Bark Beetle/Anakonda | tSJ | SRCR, Parallel beta-helix | [11,12] |
| Gliotactin | tSJ | Carboxylesterase | [10] |
| M6 | tSJ | Myelin PLP | [13] |
| Discs Large | resident | PDZ, SH3, GUK | [29] |
| Fasciclin III | resident | Ig C2-set | [47] |
| Lethal (2) giant larvae | partial resident | WD40 repeats | [48] |
| Scribble | resident | Leucine-rich, PDZ | [30] |
| Boudin | accessory | Ly6 | [33] |
| Clathrin heavy chain | accessory | Clathrin heavy chain repeat | [37,49] |
| Coiled | accessory | Ly6 | [32,34,35] |
| Crooked | accessory | Ly6 | [34] |
| Rab 5 | accessory | Small GTPase | [37] |
| Rab 11 | accessory | Small GTPase | [37] |
| Shibire | accessory | Dynamin GTPase and effector domains, PH | [37,49] |
| Wunen 1 | accessory | PA phosphatase type 2 | [36] |
| Wunen 2 | accessory | PA phosphatase type 2 | [36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rice, C.; De, O.; Alhadyian, H.; Hall, S.; Ward, R.E. Expanding the Junction: New Insights into Non-Occluding Roles for Septate Junction Proteins during Development. J. Dev. Biol. 2021, 9, 11. https://doi.org/10.3390/jdb9010011
Rice C, De O, Alhadyian H, Hall S, Ward RE. Expanding the Junction: New Insights into Non-Occluding Roles for Septate Junction Proteins during Development. Journal of Developmental Biology. 2021; 9(1):11. https://doi.org/10.3390/jdb9010011
Chicago/Turabian StyleRice, Clinton, Oindrila De, Haifa Alhadyian, Sonia Hall, and Robert E. Ward. 2021. "Expanding the Junction: New Insights into Non-Occluding Roles for Septate Junction Proteins during Development" Journal of Developmental Biology 9, no. 1: 11. https://doi.org/10.3390/jdb9010011
APA StyleRice, C., De, O., Alhadyian, H., Hall, S., & Ward, R. E. (2021). Expanding the Junction: New Insights into Non-Occluding Roles for Septate Junction Proteins during Development. Journal of Developmental Biology, 9(1), 11. https://doi.org/10.3390/jdb9010011