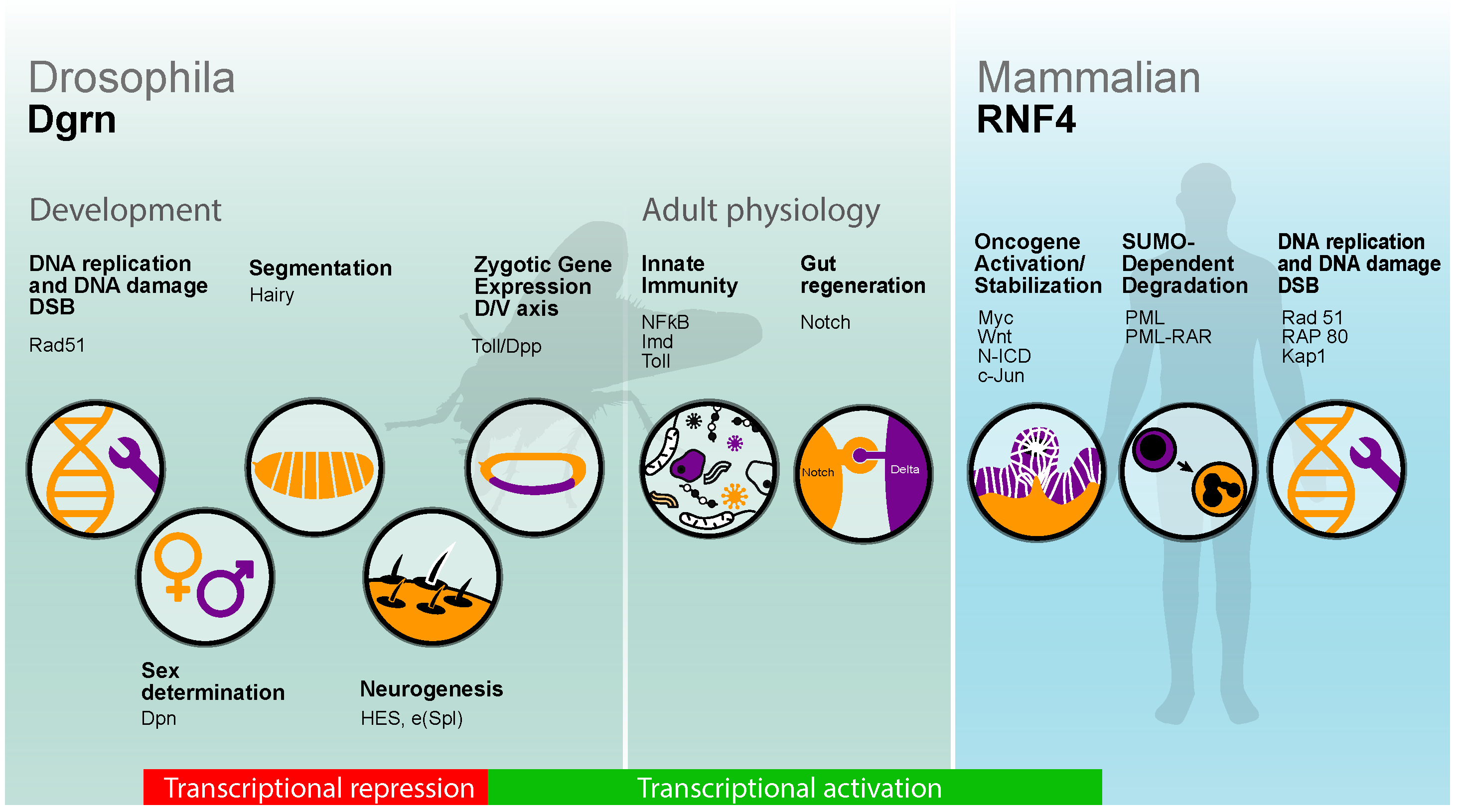
The Biology of SUMO-Targeted Ubiquitin Ligases in Drosophila Development, Immunity, and Cancer

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Ubiquitination, SUMOylation, and Their Enzymatic Connectors
2. Dgrn, Early Drosophila Embryogenesis, and DNA Repair
3. The Role of Dgrn in Transcriptional Repression
4. Dgrn Regulates Transcriptional Activation during Early Development and Adult Immune Response
5. Dgrn, the Local Regenerative Response, and Notch-Dependent Gene Expression
6. RNF4 and Transcriptional Activation in Cancer
7. Concluding Remarks and Future Challenges
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ciehanover, A.; Hod, Y.; Hershko, A. A heat-stable polypeptide component of an ATP-dependent proteolytic system from reticulocytes. Biochem. Biophys. Res. Commun. 1978, 81, 1100–1105. [Google Scholar] [CrossRef]
- Hershko, A.; Heller, H.; Elias, S.; Ciechanover, A. Components of ubiquitin-protein ligase system. Resolution, affinity purification, and role in protein breakdown. J. Biol. Chem. 1983, 258, 8206–8214. [Google Scholar] [PubMed]
- Bhogaraju, S.; Kalayil, S.; Liu, Y.; Bonn, F.; Colby, T.; Matic, I.; Dikic, I. Phosphoribosylation of Ubiquitin Promotes Serine Ubiquitination and Impairs Conventional Ubiquitination. Cell 2016, 167, 1636–1649.e13. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Clague, M.J.; Urbé, S. Breaking the chains: Structure and function of the deubiquitinases. Nat. Rev. Mol. Cell Biol. 2009, 10, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Chau, V.; Tobias, J.W.; Bachmair, A.; Marriott, D.; Ecker, D.J.; Gonda, D.K.; Varshavsky, A. A multiubiquitin chain is confined to specific lysine in a targeted short-lived protein. Science 1989, 243, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Rape, M. Ubiquitylation at the crossroads of development and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Cappadocia, L.; Lima, C.D. Ubiquitin-like Protein Conjugation: Structures, Chemistry, and Mechanism. Chem. Rev. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kerscher, O.; Felberbaum, R.; Hochstrasser, M. Modification of Proteins by Ubiquitin and Ubiquitin-Like Proteins. Annu. Rev. Cell Dev. Biol. 2006. [Google Scholar] [CrossRef] [PubMed]
- Seeler, J.S.; Dejean, A. SUMO and the robustness of cancer. Nat. Rev. Cancer 2017, 17, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Flotho, A.; Melchior, F. Sumoylation: A Regulatory Protein Modification in Health and Disease. Annu. Rev. Biochem. 2013, 82, 357–385. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Courey, A.J. SUMO in Drosophila development. Adv. Exp. Med. Biol. 2017. [Google Scholar] [CrossRef]
- Hendriks, I.A.; Schimmel, J.; Eifler, K.; Olsen, J.V.; Vertegaal, A.C.O. Ubiquitin-specific protease 11 (USP11) deubiquitinates hybrid small ubiquitin-like modifier (SUMO)-ubiquitin chains to counteract RING finger protein 4 (RNF4). J. Biol. Chem. 2015, 290, 15526–15537. [Google Scholar] [CrossRef] [PubMed]
- Abed, M.; Bitman-Lotan, E.; Orian, A. A fly view of a SUMO-targeted ubiquitin ligase. Fly (Austin) 2011, 5, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Sriramachandran, A.M.; Dohmen, R.J. SUMO-targeted ubiquitin ligases. Biochim. Biophys. Acta. 2014, 1843, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.J.P.; Tainer, J.A.; Boddy, M.N. A SIM-ultaneous role for SUMO and ubiquitin. Trends Biochem. Sci. 2008, 33, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Barry, K.C.; Abed, M.; Kenyagin, D.; Werwie, T.R.; Boico, O.; Orian, A.; Parkhurst, S.M. The Drosophila STUbL protein Degringolade limits HES functions during embryogenesis. Development 2011. [Google Scholar] [CrossRef] [PubMed]
- Abed, M.; Barry, K.C.; Kenyagin, D.; Koltun, B.; Phippen, T.M.; Delrow, J.J.; Parkhurst, S.M.; Orian, A. Degringolade, a SUMO-targeted ubiquitin ligase, inhibits Hairy/Groucho-mediated repression. EMBO J. 2011. [Google Scholar] [CrossRef] [PubMed]
- Ryu, T.; Spatola, B.; Delabaere, L.; Bowlin, K.; Hopp, H.; Kunitake, R.; Karpen, G.H.; Chiolo, I. Heterochromatic breaks move to the nuclear periphery to continue recombinational repair. Nat. Cell Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Koltun, B.; Shackelford, E.; Bonnay, F.; Matt, N.; Reichhart, J.M.; Orian, A. The SUMO-targeted ubiquitin ligase, Dgrn, is essential for Drosophila innate immunity. Int. J. Dev. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, W.; Fogarty, P.; Theurkauf, W. Mutations affecting the cytoskeletal organization of syncytial Drosophila embryos. Development 1993, 118, 1245–1254. [Google Scholar] [PubMed]
- Nie, M.; Xie, Y.; Loo, J.A.; Courey, A.J. Genetic and proteomic evidence for roles of Drosophila SUMO in cell cycle control, Ras signaling, and early pattern formation. PLoS ONE 2009, 4, e5905. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, K.J.; Woo, L.L.; Zhu, J.; Huo, D.; Matunis, M.J.; Ellis, N.A. SUMO modification regulates BLM and RAD51 interaction at damaged replication forks. PLoS Biol. 2009, 7, e1000252. [Google Scholar] [CrossRef] [PubMed]
- Streich, F.C.; Lima, C.D. Capturing a substrate in an activated RING E3/E2-SUMO complex. Nature 2016, 536, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Cipolla, L.; Maffia, A.; Bertoletti, F.; Sabbioneda, S. The regulation of DNA damage tolerance by ubiquitin and ubiquitin-like modifiers. Front. Genet. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Guzzo, C.M.; Berndsen, C.E.; Zhu, J.; Gupta, V.; Datta, A.; Greenberg, R.A.; Wolberger, C.; Matunis, M.J. RNF4-Dependent Hybrid SUMO-Ubiquitin Chains Are Signals for RAP80 and Thereby Mediate the Recruitment of BRCA1 to Sites of DNA Damage. Sci. Signal. 2012, 5, ra88. [Google Scholar] [CrossRef] [PubMed]
- Amaral, N.; Ryu, T.; Li, X.; Chiolo, I. Nuclear Dynamics of Heterochromatin Repair. Trends Genet. 2017, 33, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Vyas, R.; Kumar, R.; Clermont, F.; Helfricht, A.; Kalev, P.; Sotiropoulou, P.; Hendriks, I.A.; Radaelli, E.; Hochepied, T.; Blanpain, C.; et al. RNF4 is required for DNA double-strand break repair in vivo. Cell Death Differ. 2013, 20, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Galanty, Y.; Belotserkovskaya, R.; Coates, J.; Jackson, S.P. RNF4, a SUMO-targeted ubiquitin E3 ligase, promotes DNA double-strand break repair. Genes Dev. 2012, 26, 1179–1195. [Google Scholar] [CrossRef] [PubMed]
- Prudden, J.; Pebernard, S.; Raffa, G.; Slavin, D.A.; Perry, J.J.P.; Tainer, J.A.; McGowan, C.H.; Boddy, M.N. SUMO-targeted ubiquitin ligases in genome stability. EMBO J. 2007, 26, 4089–4101. [Google Scholar] [CrossRef] [PubMed]
- Gibbs-Seymour, I.; Oka, Y.; Rajendra, E.; Weinert, B.T.; Passmore, L.A.; Patel, K.J.; Olsen, J.V.; Choudhary, C.; Bekker-Jensen, S.; Mailand, N. Ubiquitin-SUMO circuitry controls activated fanconi anemia ID complex dosage in response to DNA damage. Mol. Cell 2015, 57, 150–164. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Kim, H.; Moreau, L.A.; Puhalla, S.; Garber, J.; Al Abo, M.; Takeda, S.; D’Andrea, A.D. RNF4-mediated polyubiquitination regulates the Fanconi anemia/BRCA pathway. J. Clin. Investig. 2015, 125, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Seifert, A.; Chua, J.S.; Maure, J.-F.; Golebiowski, F.; Hay, R.T. SUMO-targeted ubiquitin E3 ligase RNF4 is required for the response of human cells to DNA damage. Genes Dev. 2012, 26, 1196–1208. [Google Scholar] [CrossRef] [PubMed]
- Lallemand-Breitenbach, V.; Jeanne, M.; Benhenda, S.; Nasr, R.; Lei, M.; Peres, L.; Zhou, J.; Zhu, J.; Raught, B.; de Thé, H. Arsenic degrades PML or PML-RARalpha through a SUMO-triggered RNF4/ubiquitin-mediated pathway. Nat. Cell Biol. 2008, 10, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Bruderer, R.; Tatham, M.H.; Plechanovova, A.; Matic, I.; Garg, A.K.; Hay, R.T. Purification and identification of endogenous polySUMO conjugates. EMBO Rep. 2011, 12, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.J.; Abed, M.; Heuberger, J.; Novak, R.; Zohar, Y.; Beltran Lopez, A.P.; Trausch-Azar, J.S.; Ilagan, M.X.G.; Benhamou, D.; Dittmar, G.; et al. RNF4-Dependent Oncogene Activation by Protein Stabilization. Cell Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Poortinga, G.; Watanabe, M.; Susan, S.M. Drosophila CtBP: A Hairy-interacting protein required for embryonic segmentation and Hairy-mediated transcriptional repression. EMBO J. 1998, 17, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Rubenstein, E.M.; Matt, T.; Hochstrasser, M. SUMO-independent in vivo activity of a SUMO-targeted ubiquitin ligase toward a short-lived transcription factor. Genes Dev. 2010, 24, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Bianchi-Frias, D.; Orian, A.; Delrow, J.J.; Vazquez, J.; Rosales-Nieves, A.E.; Parkhurst, S.M. Hairy transcriptional repression targets and cofactor recruitment in Drosophila. PLoS Biol. 2004, 2, e178. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.I.; Parkhurst, S.M. Drosophila Sir2 is required for heterochromatic silencing and by euchromatic Hairy/E(spl) bHLH repressors in segmentation and sex determination. Cell 2002, 109, 447–458. [Google Scholar] [CrossRef]
- Ahn, J.W.; Lee, Y.A.; Ahn, J.H.; Choi, C.Y. Covalent conjugation of Groucho with SUMO-1 modulates its corepressor activity. Biochem. Biophys. Res. Commun. 2009, 379, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; González-Prieto, R.; Xiao, Z.; Verlaan-De Vries, M.; Vertegaal, A.C.O. The STUbL RNF4 regulates protein group SUMOylation by targeting the SUMO conjugation machinery. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.B.; Scott, M.P. Zygotically active genes that affect the spatial expression of the fushi tarazu segmentation gene during early Drosophila embryogenesis. Cell 1986, 45, 113–126. [Google Scholar] [CrossRef]
- Parkhurst, S.M.; Ish-Horowicz, D. Mis-regulating segmentation gene expression in Drosophila. Development 1991, 111, 1121–1135. [Google Scholar] [PubMed]
- Kellerman, K.A.; Mattson, D.M.; Duncan, I. Mutations affecting the stability of the fushi tarazu protein of Drosophila. Genes Dev. 1990, 4, 1936–1950. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.J.; Welcker, M.; Clurman, B.E. Tumor suppression by the Fbw7 ubiquitin ligase: Mechanisms and opportunities. Cancer Cell 2014, 26, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Ingham, P.W.; Pinchin, S.M.; Howard, K.R.; Ish-Horowicz, D. Genetic analysis of the hairy locus in Drosophila melanogaster. Genetics 1985, 111, 463–486. [Google Scholar] [PubMed]
- Davis, R.L.; Turner, D.L. Vertebrate hairy and Enhancer of split related proteins: Transcriptional repressors regulating cellular differentiation and embryonic patterning. Oncogene 2001, 20, 8342–8357. [Google Scholar] [CrossRef] [PubMed]
- Delidakis, C.; Monastirioti, M.; Magadi, S.S. E(spl): Genetic, developmental, and evolutionary aspects of a group of invertebrate hes proteins with close ties to notch signaling. Curr. Top. Dev. Biol. 2014, 110, 217–262. [Google Scholar] [CrossRef] [PubMed]
- Younger-Shepherd, S.; Vaessin, H.; Bier, E.; Jan, L.Y.; Jan, Y.N. Deadpan, an essential pan-neural gene encoding an HLH protein, acts as a denominator in Drosophila sex determination. Cell 1992, 70, 911–922. [Google Scholar] [CrossRef]
- Estes, P.A.; Keyes, L.N.; Schedl, P. Multiple response elements in the sex-lethal early promoter ensure its female-specific expression pattern. Mol. Cell. Biol. 1995, 15, 904–917. [Google Scholar] [CrossRef] [PubMed]
- Cline, T.W. Two closely linked mutations in Drosophila melanogaster that are lethal to opposite sexes and interact with daughterless. Genetics 1978, 90, 683–697. [Google Scholar] [PubMed]
- Poukka, H.; Aarnisalo, P.; Santti, H.; Jänne, O.A.; Palvimo, J.J. Coregulator small nuclear RING finger protein (SNURF) enhances Sp1- and steroid receptor-mediated transcription by different mechanisms. J. Biol. Chem. 2000, 275, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.V.; Jürgens, G.; Nüsslein-Volhard, C. Establishment of dorsal-ventral polarity in the Drosophila embryo: Genetic studies on the role of the Toll gene product. Cell 1985, 42, 779–789. [Google Scholar] [CrossRef]
- Smith, M.; Turki-Judeh, W.; Courey, A.J. SUMOylation in Drosophila development. Biomolecules 2012, 2, 331–349. [Google Scholar] [CrossRef] [PubMed]
- Steward, R. Dorsal, an embryonic polarity gene in Drosophila, is homologous to the vertebrate proto-oncogene, c-rel. Science 1987, 238, 692–694. [Google Scholar] [CrossRef] [PubMed]
- Ip, Y.T.; Reach, M.; Engstrom, Y.; Kadalayil, L.; Cai, H.; González-Crespo, S.; Tatei, K.; Levine, M. Dif, a dorsal-related gene that mediates an immune response in Drosophila. Cell 1993, 75, 753–763. [Google Scholar] [CrossRef]
- Steward, R.; Zusman, S.B.; Huang, L.H.; Schedl, P. The dorsal protein is distributed in a gradient in early Drosophila embryos. Cell 1988, 55, 487–495. [Google Scholar] [CrossRef]
- Minakhina, S.; Steward, R. Nuclear factor-kappa B pathways in Drosophila. Oncogene 2006, 25, 6749–6757. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Hoffmann, J. The Host Defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, V.; Smith, M.; Courey, A.J. Conjugation of Smt3 to dorsal may potentiate the Drosophila immune response. Mol. Cell. Biol. 2002, 22, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Paddibhatla, I.; Lee, M.J.; Kalamarz, M.E.; Ferrarese, R.; Govind, S. Role for Sumoylation in Systemic Inflammation and Immune Homeostasis in Drosophila Larvae. PLoS Pathog. 2010, 6, e1001234. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.G.; Xu, W.; Nikkholgh, N.; Basu, S.; Nie, Y.; Thomas, M.; Satyamurti, M.; Budnik, B.A.; Ip, Y.T.; Veraksa, A. Regulation of toll signaling and inflammation by β-Arrestin and the SUMO protease Ulp1. Genetics 2013, 195, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, V.; Valentine, S.A.; Courey, A.J. A functional interaction between dorsal and components of the Smt3 conjugation machinery. J. Biol. Chem. 2000, 275, 4033–4040. [Google Scholar] [CrossRef] [PubMed]
- Minakhina, S.; Steward, R. Melanotic mutants in Drosophila: Pathways and phenotypes. Genetics 2006, 174, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Broderick, N.A.; Lemaitre, B. Gut homeostasis in a microbial world: Insights from Drosophila melanogaster. Nat. Rev. Microbiol. 2013, 11, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Edgar, B.A. Intestinal stem cell function in Drosophila and mice. Curr. Opin. Genet. Dev. 2012, 22, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Lucchetta, E.; Rafel, N.; Ohlstein, B. Maintenance of the adult Drosophila intestine: All roads lead to homeostasis. Curr. Opin. Genet. Dev. 2016, 40, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Kux, K.; Pitsouli, C. Tissue communication in regenerative inflammatory signaling: Lessons from the fly gut. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Boquete, J.P.; Lemaitre, B. A genetic framework controlling the differentiation of intestinal stem cells during regeneration in Drosophila. PLoS Genet. 2017, 13, e1006854. [Google Scholar] [CrossRef] [PubMed]
- Vooijs, M.; Liu, Z.; Kopan, R. Notch: Architect, landscaper, and guardian of the intestine. Gastroenterology 2011, 141, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Rijsewijk, F.; Schuermann, M.; Wagenaar, E.; Parren, P.; Weigel, D.; Nusse, R. The Drosophila homology of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless. Cell 1987, 50, 649–657. [Google Scholar] [CrossRef]
- Klaus, A.; Birchmeier, W. Wnt signalling and its impact on development and cancer. Nat. Rev. Cancer 2008, 8, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Jenny, F.H.; Basler, K. Powerful Drosophila screens that paved the wingless pathway. Fly (Austin) 2014, 8, 218–225. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sansom, O.J.; Meniel, V.S.; Muncan, V.; Phesse, T.J.; Wilkins, J.A.; Reed, K.R.; Vass, J.K.; Athineos, D.; Clevers, H.; Clarke, A.R. Myc deletion rescues Apc deficiency in the small intestine. Nature 2007, 446, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Weng, A.P.; Millholland, J.M.; Yashiro-Ohtani, Y.; Arcangeli, M.L.; Lau, A.; Wai, C.; Del Bianco, C.; Rodriguez, C.G.; Sai, H.; Tobias, J.; et al. c-Myc is an important direct target of Notch1 in T-cell acute lymphoblastic leukemia/lymphoma. Genes Dev. 2006, 20, 2096–2109. [Google Scholar] [CrossRef] [PubMed]
- Herranz, D.; Ambesi-Impiombato, A.; Palomero, T.; Schnell, S.A.; Belver, L.; Wendorff, A.A.; Xu, L.; Castillo-Martin, M.; Llobet-Navás, D.; Cordon-Cardo, C.; et al. A NOTCH1-driven MYC enhancer promotes T cell development, transformation and acute lymphoblastic leukemia. Nat. Med. 2014, 20, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Reymann, S.; Borlak, J. Transcription profiling of lung adenocarcinomas of c-myc-transgenic mice: Identification of the c-myc regulatory gene network. BMC Syst. Biol. 2008, 2. [Google Scholar] [CrossRef] [PubMed]
- Zeller, K.I.; Jegga, A.G.; Aronow, B.J.; O’Donnell, K.A.; Dang, C.V. An integrated database of genes responsive to the Myc oncogenic transcription factor: Identification of direct genomic targets. Genome Biol. 2003, 4, R69. [Google Scholar] [CrossRef] [PubMed]
- Gallant, P.; Shiio, Y.; Cheng, P.F.; Parkhurst, S.M.; Eisenman, R.N. Myc and Max homologs in Drosophila. Science. 1996, 274, 1523–1527. [Google Scholar] [CrossRef] [PubMed]
- Pierce, S.B. dMyc is required for larval growth and endoreplication in Drosophila. Development 2004, 131, 2317–2327. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.L.; Nien, C.Y.; Liu, H.Y.; Metzstein, M.M.; Kirov, N.; Rushlow, C. The zinc-finger protein Zelda is a key activator of the early zygotic genome in Drosophila. Nature 2008, 456, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Hug, C.B.; Grimaldi, A.G.; Kruse, K.; Vaquerizas, J.M. Chromatin Architecture Emerges during Zygotic Genome Activation Independent of Transcription. Cell 2017, 169, 216–228.e19. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.A. Direct lineage reprogramming via pioneer factors; a detour through developmental gene regulatory networks. Development 2016, 143, 2696–2705. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.V.; Rodrigues, T.M.A.; Tao, H.; Baker, R.K.; Miraglia, L.; Orth, A.P.; Lyons, G.E.; Schultz, P.G.; Wu, X. Identification of RING finger protein 4 (RNF4) as a modulator of DNA demethylation through a functional genomics screen. Proc. Natl. Acad. Sci. USA 2010, 107, 15087–15092. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, T.; Fujimitsu, Y.; Yoshikai, Y.; Sasano, T.; Yamada, K.; Murakami, M.; Urano, T.; Sugasawa, K.; Saitoh, H. SUMO-modification and elimination of the active DNA demethylation enzyme TDG in cultured human cells. Biochem. Biophys. Res. Commun. 2014, 447, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. RING finger protein 4 (RNF4) derepresses gene expression from DNA methylation. J. Biol. Chem. 2014, 289, 33808–33813. [Google Scholar] [CrossRef] [PubMed]
- Ahner, A.; Gong, X.; Schmidt, B.Z.; Peters, K.W.; Rabeh, W.M.; Thibodeau, P.H.; Lukacs, G.L.; Frizzell, R.A. Small heat shock proteins target mutant cystic fibrosis transmembrane conductance regulator for degradation via a small ubiquitin-like modifier-dependent pathway. Mol. Biol. Cell 2013, 24, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Solimini, N.L.; Elledge, S.J. Principles of Cancer Therapy: Oncogene and Non-oncogene Addiction. Cell 2009, 136, 823–837. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abed, M.; Bitman-Lotan, E.; Orian, A. The Biology of SUMO-Targeted Ubiquitin Ligases in Drosophila Development, Immunity, and Cancer. J. Dev. Biol. 2018, 6, 2. https://doi.org/10.3390/jdb6010002
Abed M, Bitman-Lotan E, Orian A. The Biology of SUMO-Targeted Ubiquitin Ligases in Drosophila Development, Immunity, and Cancer. Journal of Developmental Biology. 2018; 6(1):2. https://doi.org/10.3390/jdb6010002
Chicago/Turabian StyleAbed, Mona, Eliya Bitman-Lotan, and Amir Orian. 2018. "The Biology of SUMO-Targeted Ubiquitin Ligases in Drosophila Development, Immunity, and Cancer" Journal of Developmental Biology 6, no. 1: 2. https://doi.org/10.3390/jdb6010002
APA StyleAbed, M., Bitman-Lotan, E., & Orian, A. (2018). The Biology of SUMO-Targeted Ubiquitin Ligases in Drosophila Development, Immunity, and Cancer. Journal of Developmental Biology, 6(1), 2. https://doi.org/10.3390/jdb6010002