
Hedgehog Signaling in Prostate Development, Regeneration and Cancer

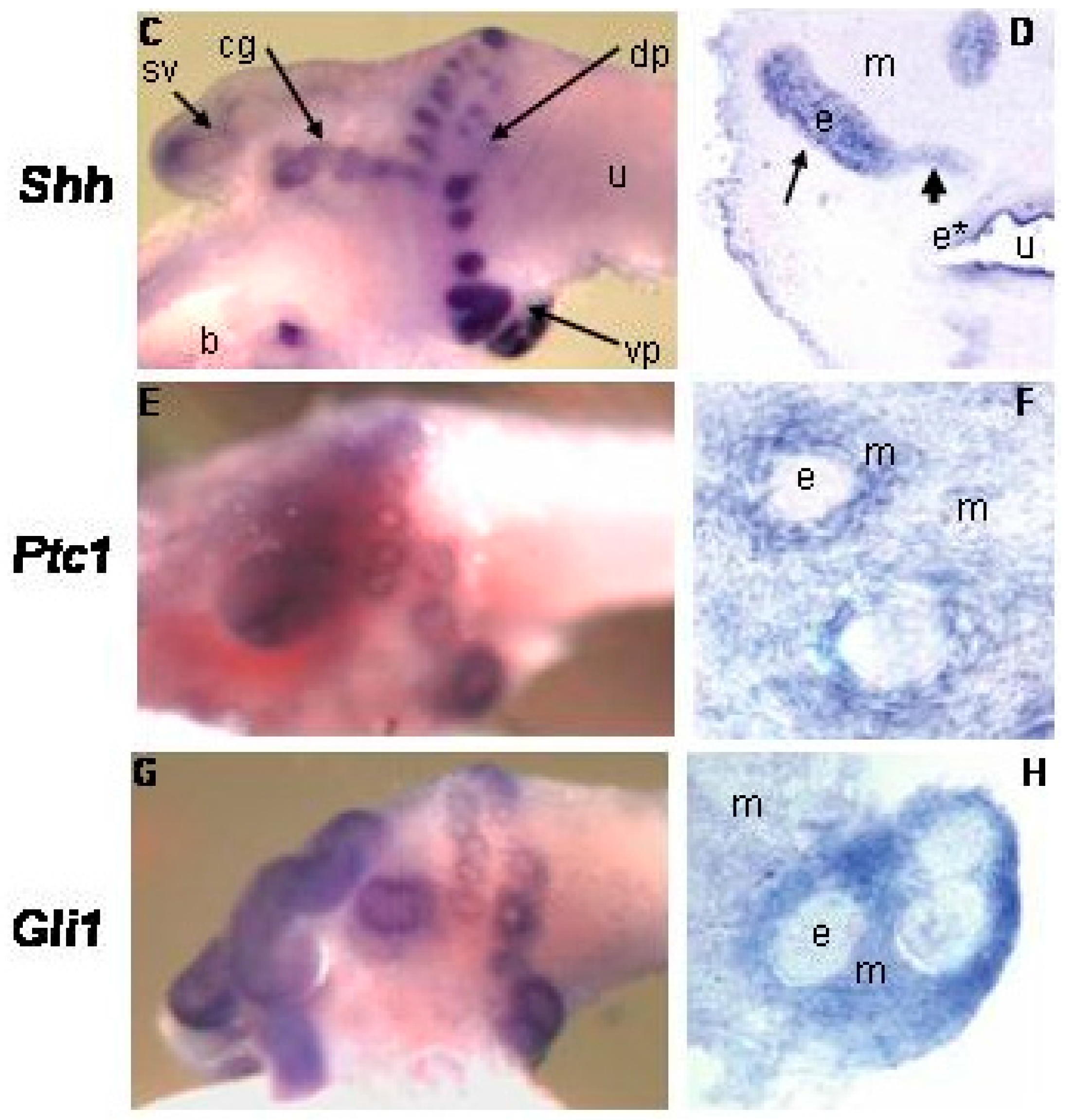
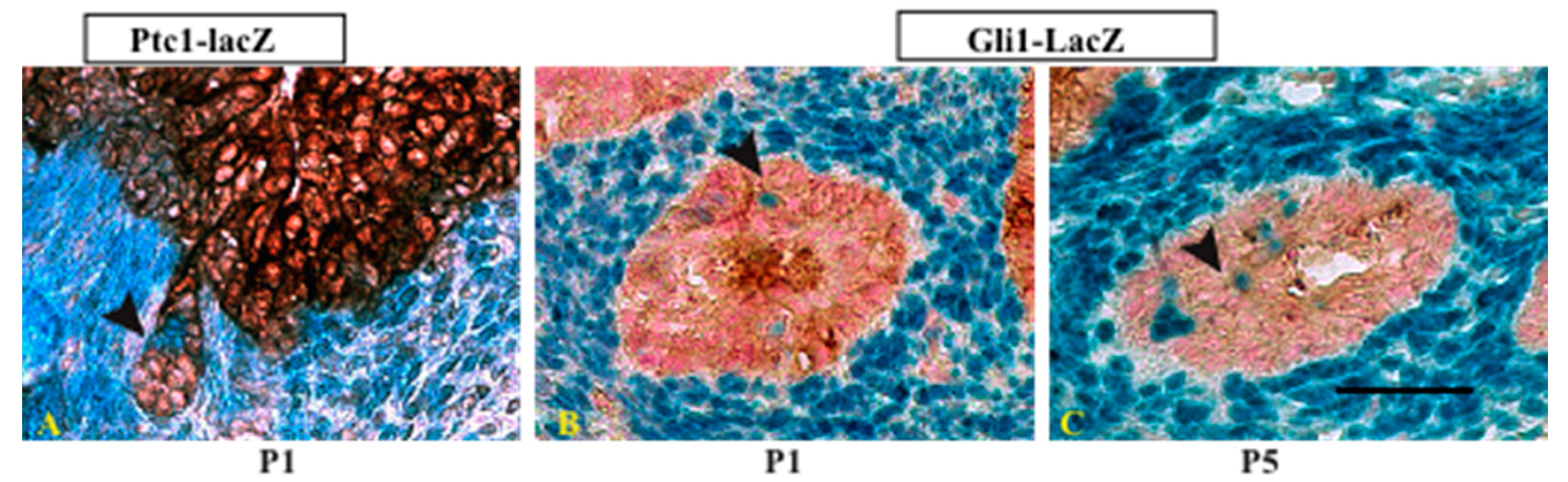
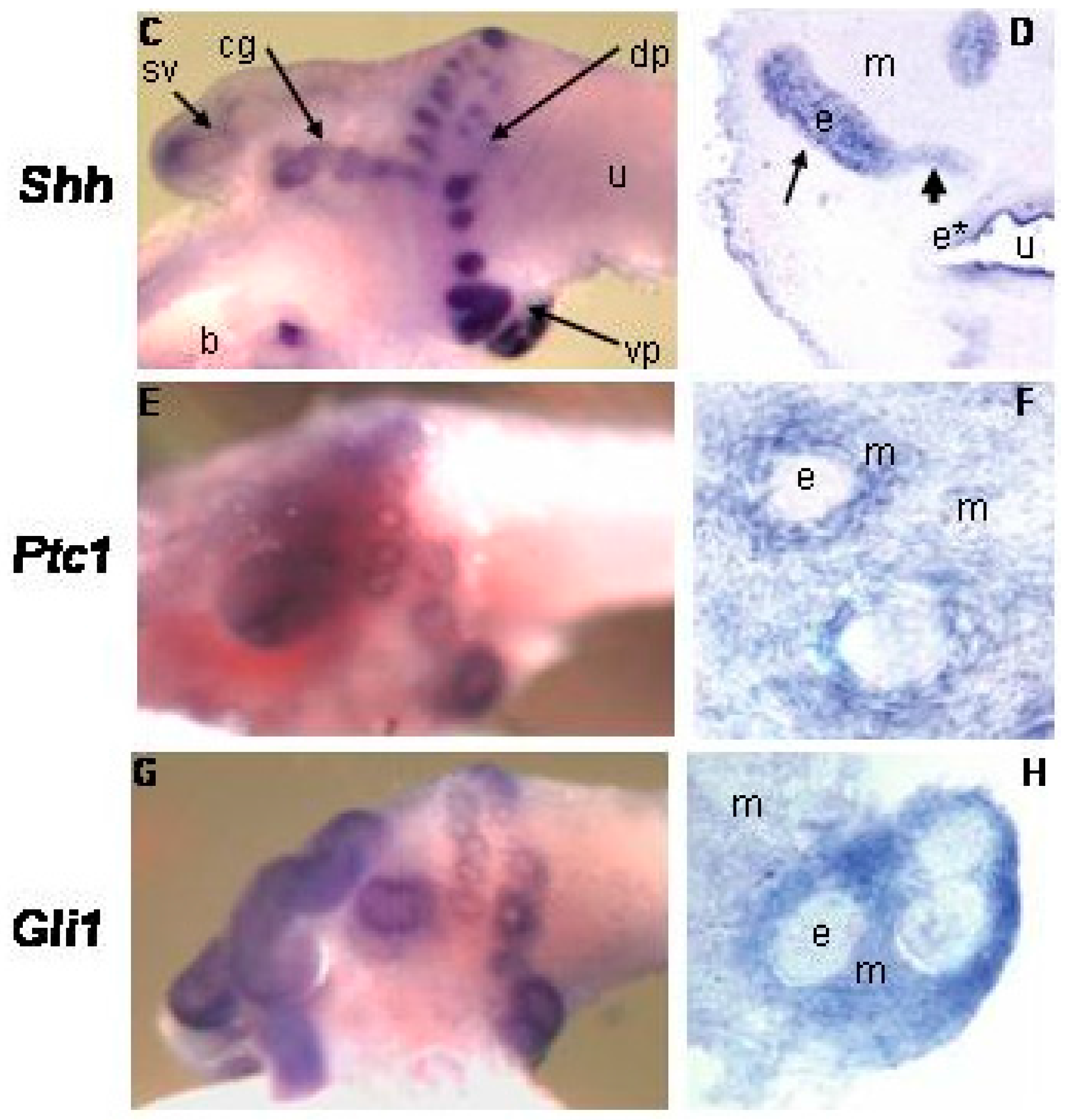{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
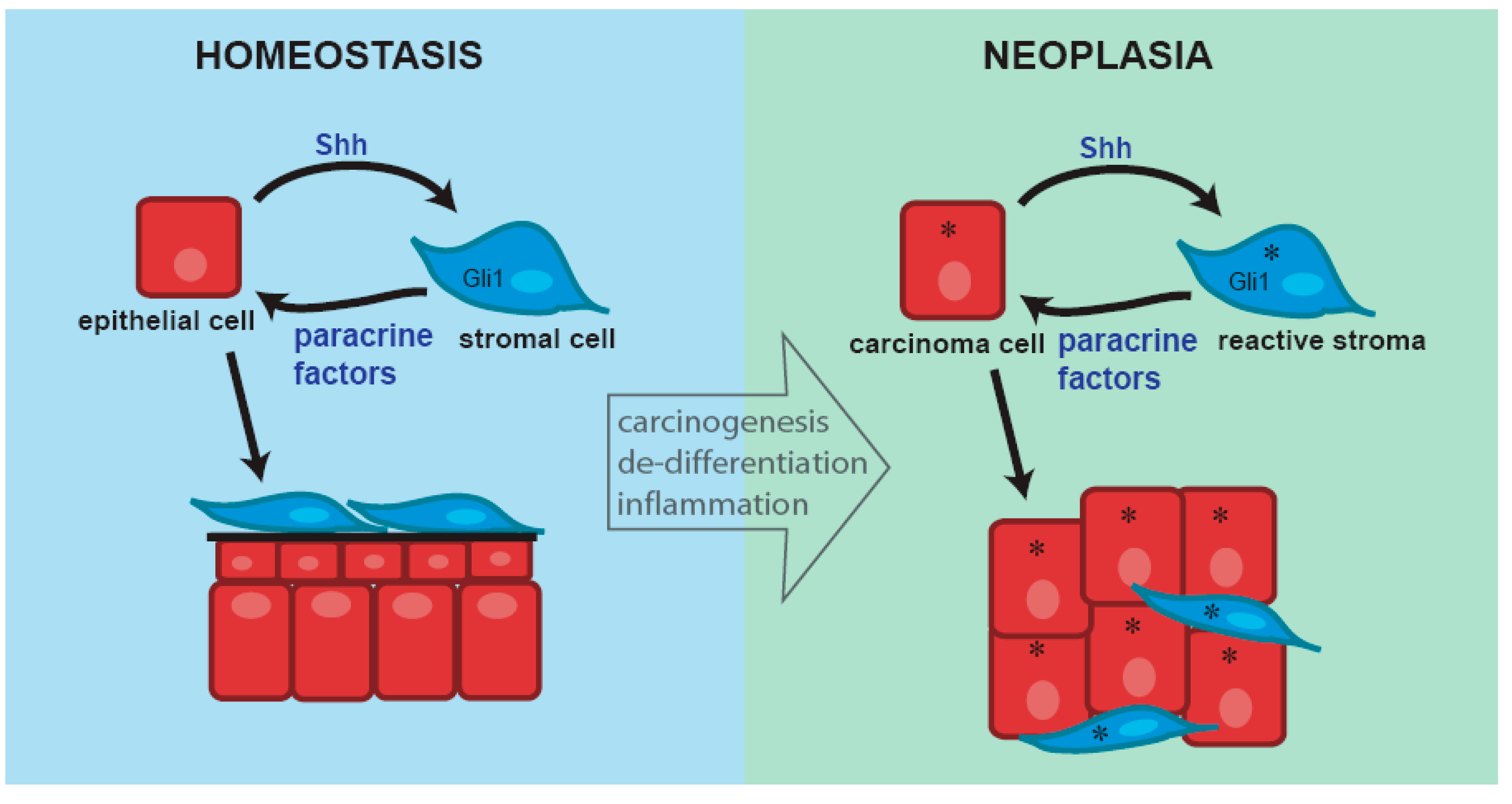{kind=link}
Abstract
:1. Prostate Development
2. Hedgehog Signaling in the Developing Prostate
3. Requirement for Prostate Development
4. Effects on Growth and Ductal Morphogenesis
5. Autocrine Signaling
6. Prostate Regeneration
7. Prostate Cancer
8. Summary and Conclusions
Acknowledgments
Conflicts of Interest
References
- Cunha, G.R.; Sekkingstad, M.; Meloy, B.A. Heterospecific induction of prostatic development in tissue recombinants prepared with mouse, rat, rabbit and human tissues. Differentiation 1983, 24, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, Y.; Cunha, G.R.; Donjacour, A.A. Morphogenesis of ductal networks in the mouse prostate. Biol. Reprod. 1986, 34, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Podlasek, C.A.; Barnett, D.H.; Clemens, J.Q.; Bak, P.M.; Bushman, W. Prostate development requires Sonic hedgehog expressed by the urogenital sinus epithelium. Dev. Biol. 1999, 209, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Lamm, M.L.; Catbagan, W.S.; Laciak, R.J.; Barnett, D.H.; Hebner, C.M.; Gaffield, W.; Walterhouse, D.; Iannaccone, P.; Bushman, W. Sonic hedgehog activates mesenchymal Gli1 expression during prostate ductal bud formation. Dev. Biol. 2002, 249, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.E.; Shou, J.; Ross, S.; Koeppen, H.; de Sauvage, F.J.; Gao, W.Q. Inhibition of epithelial ductal branching in the prostate by sonic hedgehog is indirectly mediated by stromal cells. J. Biol. Chem. 2003, 278, 18506–18513. [Google Scholar] [CrossRef] [PubMed]
- Freestone, S.H.; Marker, P.; Grace, O.C.; Tomlinson, D.C.; Cunha, G.R.; Harnden, P.; Thomson, A.A. Sonic hedgehog regulates prostatic growth and epithelial differentiation. Dev. Biol. 2003, 264, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Berman, D.M.; Desai, N.; Wang, X.; Karhadkar, S.S.; Reynon, M.; Abate-Shen, C.; Beachy, P.A.; Shen, M.M. Roles for Hedgehog signaling in androgen production and prostate ductal morphogenesis. Dev. Biol. 2004, 267, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Bushman, W. Differential stage-dependent regulation of prostatic epithelial morphogenesis by Hedgehog signaling. Dev. Biol. 2013, 380, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Gipp, J.; Yoon, J.W.; Iannaccone, P.; Walterhouse, D.; Bushman, W. Sonic hedgehog-responsive genes in the fetal prostate. J. Biol. Chem. 2009, 284, 5620–5629. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Ishii, K.; Mirosevich, J.; Kuwajima, S.; Oppenheimer, S.R.; Roberts, R.L.; Jiang, M.; Yu, X.; Shappell, S.B.; Caprioli, R.M.; et al. Forkhead box A1 regulates prostate ductal morphogenesis and promotes epithelial cell maturation. Development 2005, 132, 3431–3443. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Huang, L.; Prins, G.S. Sonic hedgehog-patched Gli signaling in the developing rat prostate gland: Lobe-specific suppression by neonatal estrogens reduces ductal growth and branching. Dev. Biol. 2004, 273, 257–275. [Google Scholar] [CrossRef] [PubMed]
- Doles, J.; Cook, C.; Shi, X.; Valosky, J.; Lipinski, R.; Bushman, W. Functional compensation in Hedgehog signaling during mouse prostate development. Dev. Biol. 2006, 295, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.; Shin, K.; Zhao, C.; Kawano, S.; Beachy, P.A. Spatially restricted Hedgehog signalling regulates HGF-induced branching of the adult prostate. Nat. Cell Biol. 2014, 16, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.K.; Francis, J.C.; Swain, A. The role of Sox9 in prostate development. Differentiation 2008, 76, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Mehta, V.; Schmitz, C.T.; Keil, K.P.; Joshi, P.S.; Abler, L.L.; Lin, T.M.; Taketo, M.M.; Sun, X.; Vezina, C.M. Beta-catenin (CTNNB1) induces Bmp expression in urogenital sinus epithelium and participates in prostatic bud initiation and patterning. Dev. Biol. 2013, 376, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Doles, J.D.; Vezina, C.M.; Lipinski, R.J.; Peterson, R.E.; Bushman, W. Growth, morphogenesis, and differentiation during mouse prostate development in situ, in renal grafts, and in vitro. Prostate 2005, 65, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Bushman, W.; University of Wisconsin, Madison, WI, USA. Unpublished work. 2013.
- Shi, X.; Gipp, J.; Bushman, W. Anchorage-independent culture maintains prostate stem cells. Dev. Biol. 2007, 312, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Karhadkar, S.S.; Bova, G.S.; Abdallah, N.; Dhara, S.; Gardner, D.; Maitra, A.; Isaacs, J.T.; Berman, D.M.; Beachy, P.A. Hedgehog signalling in prostate regeneration, neoplasia and metastasis. Nature 2004, 431, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Pepicelli, C.V.; Dibble, C.C.; Catbagan, W.; Zarycki, J.L.; Laciak, R.; Gipp, J.; Shaw, A.; Lamm, M.L.; Munoz, A.; et al. Hedgehog signaling promotes prostate xenograft tumor growth. Endocrinology 2004, 145, 3961–3970. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, P.; Hernández, A.M.; Stecca, B.; Kahler, A.J.; DeGueme, A.M.; Barrett, A.; Beyna, M.; Datta, M.W.; Datta, S.; Ruiz i Altaba, A. Inhibition of prostate cancer proliferation by interference with SONIC HEDGEHOG-GLI1 signaling. Proc. Natl. Acad. Sci. USA 2004, 101, 12561–12566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lipinski, R.; Shaw, A.; Gipp, J.; Bushman, W. Lack of demonstrable autocrine hedgehog signaling in human prostate cancer cell lines. J. Urol. 2007, 177, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, F.R.; Brown, A.J. Autonomous Hedgehog signalling is undetectable in PC-3 prostate cancer cells. Biochem. Biophys. Res. Commun. 2008, 373, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Sheng, T.; Li, C.; Zhang, X.; Chi, S.; He, N.; Chen, K.; McCormick, F.; Gatalica, Z.; Xie, J. Activation of the hedgehog pathway in advanced prostate cancer. Mol. Cancer 2004, 3, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bushman, W. Hedgehog Signaling in Prostate Development, Regeneration and Cancer. J. Dev. Biol. 2016, 4, 30. https://doi.org/10.3390/jdb4040030
Bushman W. Hedgehog Signaling in Prostate Development, Regeneration and Cancer. Journal of Developmental Biology. 2016; 4(4):30. https://doi.org/10.3390/jdb4040030
Chicago/Turabian StyleBushman, Wade. 2016. "Hedgehog Signaling in Prostate Development, Regeneration and Cancer" Journal of Developmental Biology 4, no. 4: 30. https://doi.org/10.3390/jdb4040030