

Cannabinoid Receptor 1 Regulates Zebrafish Renal Multiciliated Cell Development via cAMP Signaling
 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Zebrafish Husbandry
2.2. Animal Models
2.3. Whole Mount in Situ Hybridization (WISH)
2.4. Morpholino Knockdown
2.5. Immunofluorescence (IF)
2.6. Drug Treatments
2.7. PCT Phenotype Scoring
2.8. Image Acquisition
2.9. Quantification and Statistical Analysis
3. Results
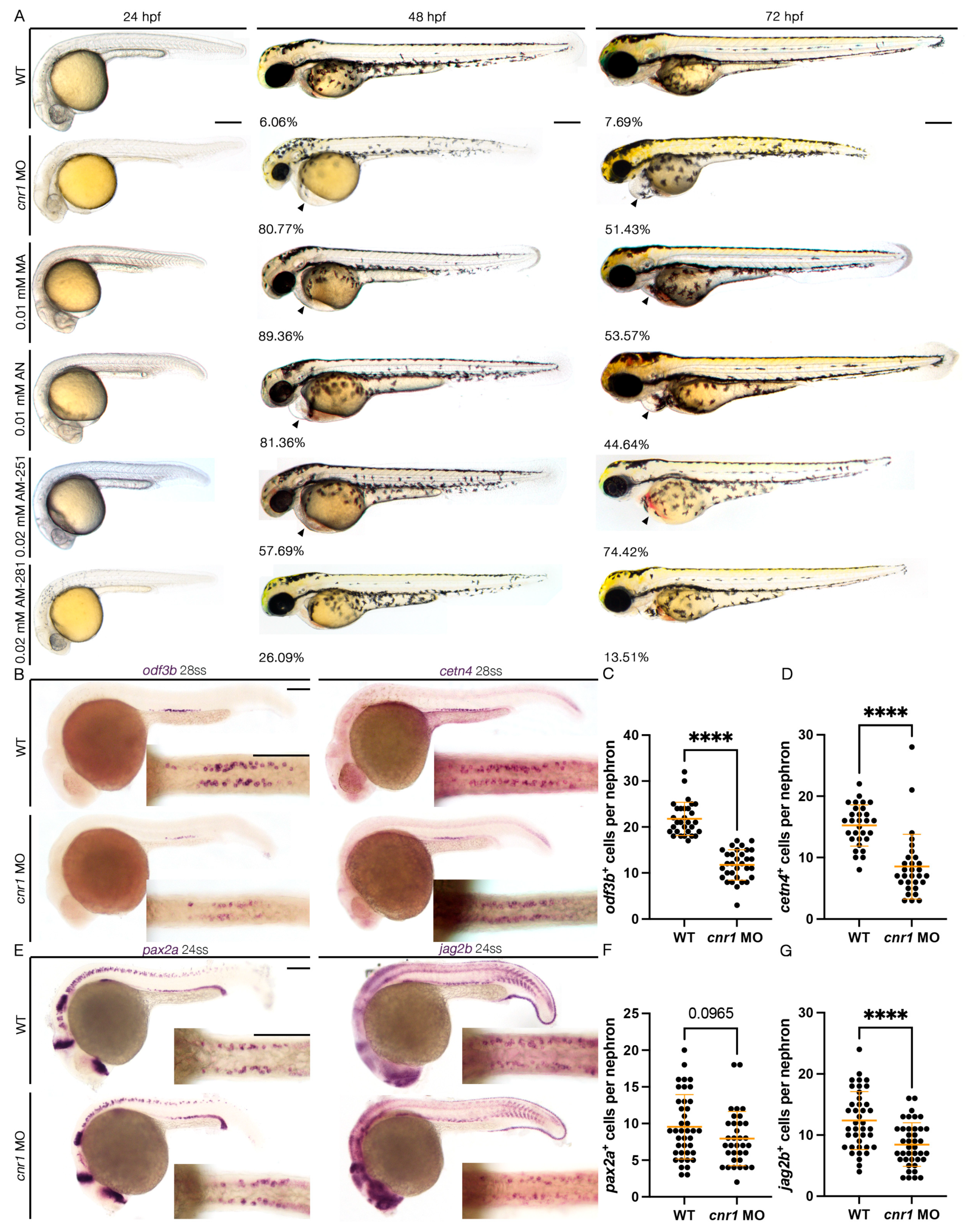
3.1. Loss of cnr1 Leads to Phenotypes Consistent with Renal MCC Defects
3.2. Cnr1 Agonism Leads to Phenotypes Consistent with Renal MCC Defects
3.3. Cnr1 Antagonism Leads to Phenotypes Consistent with Renal MCC Defects
3.4. Assessing Renal Segmental Differences with cnr1 Deficiency, Cnr1 Agonism and Antagonism
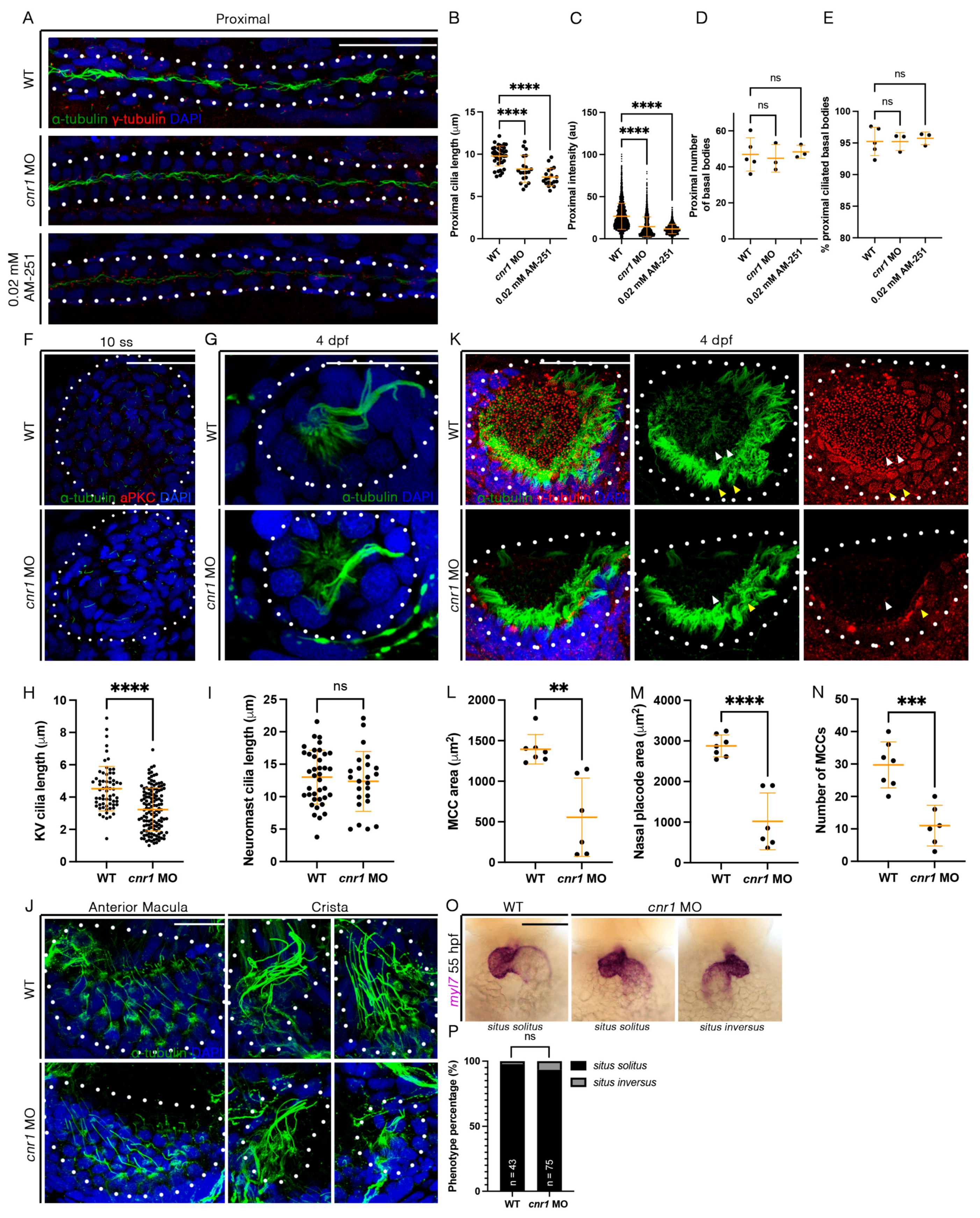
3.5. cnr1 Deficiency Leads to Reduced Cilia Development Across Zebrafish Tissues
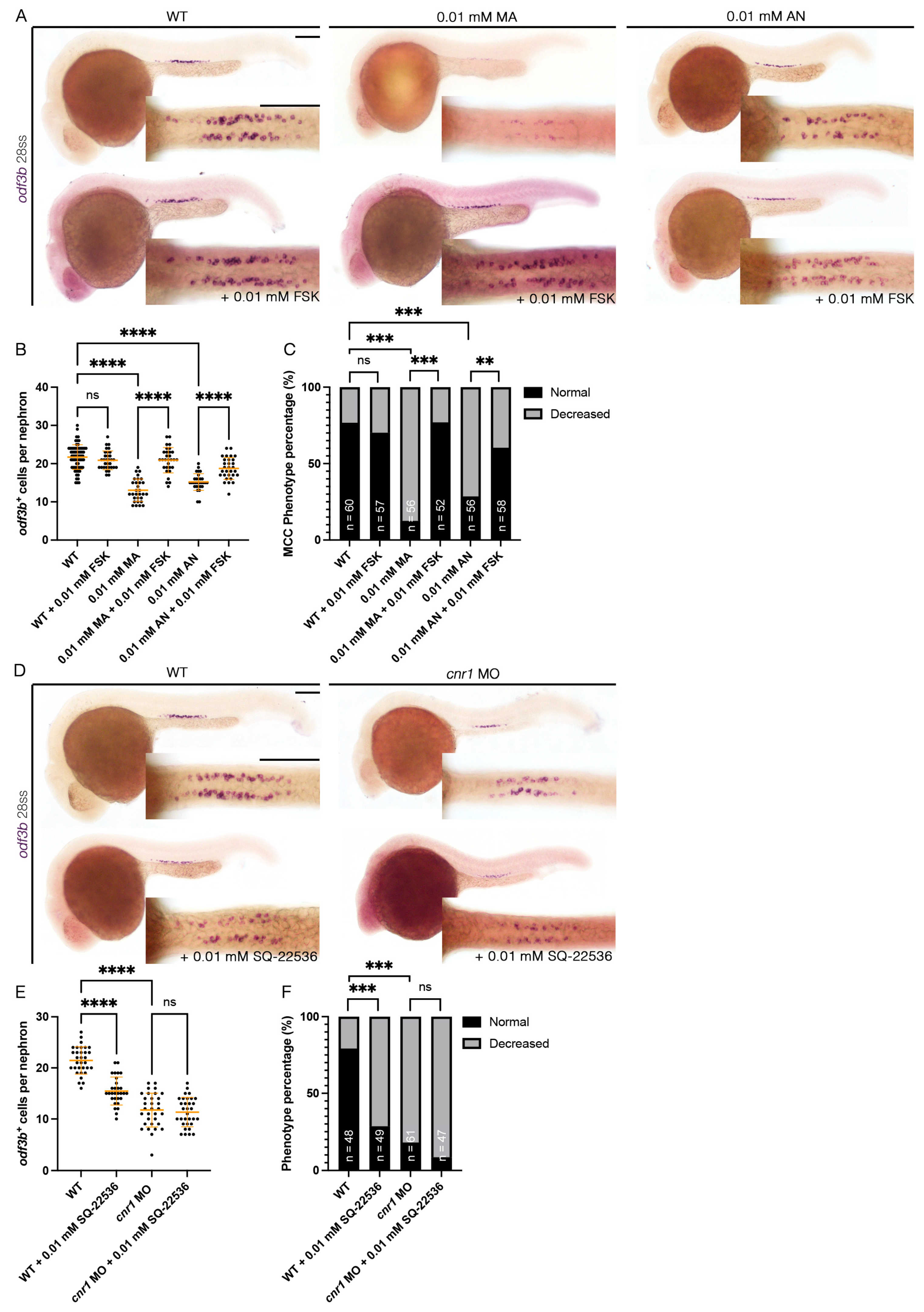
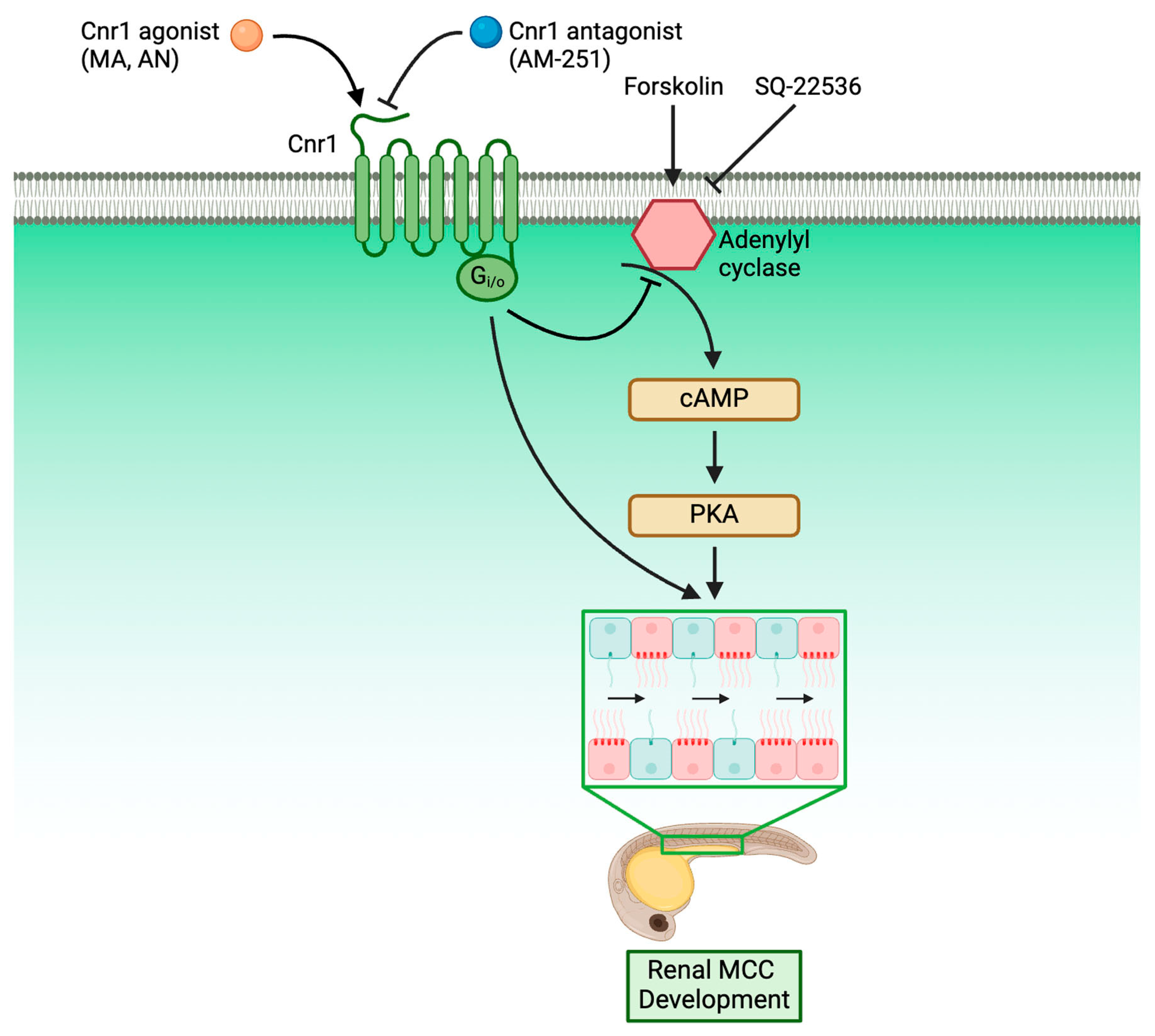
3.6. Forskolin Rescues Renal MCC Reduction in Cnr1 Agonist-Treated Embryos
3.7. cnr1 Likely Governs Renal MCC Development via Additional Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arceri, L.; Nguyen, T.K.; Gibson, S.; Baker, S.; Wingert, R.A. Cannabinoid Signaling in Kidney Disease. Cells 2023, 12, 1419. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of Cannabinoid Receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Kendall, D.A.; Yudowski, G.A. Cannabinoid Receptors in the Central Nervous System: Their Signaling and Roles in Disease. Front. Cell Neurosci. 2016, 10, 294. [Google Scholar] [CrossRef]
- Allam, S.; Paris, E.; Lazcano, I.; Bitterman, P.; Basu, S.; O’Donnell, J.; Barua, A. Detection of Cannabinoid Receptor Expression by Endometriotic Lesions in Women with Endometriosis as an Alternative to Opioid-Based Pain Medication. J. Immunol. Res. 2022, 2022, 4323259. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.; François, H. Cannabinoid Receptor 1 Inhibition in Chronic Kidney Disease: A New Therapeutic Toolbox. Front. Endocrinol. 2021, 12, 720734. [Google Scholar] [CrossRef] [PubMed]
- Hasenoehrl, C.; Taschler, U.; Storr, M.; Schicho, R. The Gastrointestinal Tract—A Central Organ of Cannabinoid Signaling in Health and Disease. Neurogastroenterol. Motil. 2016, 28, 1765–1780. [Google Scholar] [CrossRef]
- Haspula, D.; Clark, M.A. Cannabinoid Receptors: An Update on Cell Signaling, Pathophysiological Roles and Therapeutic Opportunities in Neurological, Cardiovascular, and Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 7693. [Google Scholar] [CrossRef]
- Osei-Hyiaman, D.; DePetrillo, M.; Pacher, P.; Liu, J.; Radaeva, S.; Bátkai, S.; Harvey-White, J.; Mackie, K.; Offertáler, L.; Wang, L.; et al. Endocannabinoid Activation at Hepatic CB1 Receptors Stimulates Fatty Acid Synthesis and Contributes to Diet-Induced Obesity. J. Clin. Investig. 2005, 115, 1298–1305. [Google Scholar] [CrossRef]
- Dalle, S.; Koppo, K. Cannabinoid Receptor 1 Expression Is Higher in Muscle of Old vs. Young Males, and Increases upon Resistance Exercise in Older Adults. Sci. Rep. 2021, 11, 18349. [Google Scholar] [CrossRef]
- Zottola, A.C.P.; Severi, I.; Cannich, A.; Ciofi, P.; Cota, D.; Marsicano, G.; Giordano, A.; Bellocchio, L. Expression of Functional Cannabinoid Type-1 (CB1) Receptor in Mitochondria of White Adipocytes. Cells 2022, 11, 2582. [Google Scholar] [CrossRef]
- Liu, J.; Gao, B.; Mirshahi, F.; Sanyal, A.J.; Khanolkar, A.D.; Makriyannis, A.; Kunos, G. Functional CB1 Cannabinoid Receptors in Human Vascular Endothelial Cells. Biochem. J. 2000, 346, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Puhl, S.-L. Cannabinoid-Sensitive Receptors in Cardiac Physiology and Ischaemia. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2020, 1867, 118462. [Google Scholar] [CrossRef] [PubMed]
- Drori, A.; Permyakova, A.; Hadar, R.; Udi, S.; Nemirovski, A.; Tam, J. Cannabinoid-1 Receptor Regulates Mitochondrial Dynamics and Function in Renal Proximal Tubular Cells. Diabetes Obes. Metab. 2019, 21, 146–159. [Google Scholar] [CrossRef]
- Larrinaga, G.; Varona, A.; Pérez, I.; Sanz, B.; Ugalde, A.; Cándenas, M.L.; Pinto, F.M.; Gil, J.; López, J.I. Expression of Cannabinoid Receptors in Human Kidney. Histol. Histopathol. 2010, 25, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Koura, Y.; Ichihara, A.; Tada, Y.; Kaneshiro, Y.; Okada, H.; Temm, C.J.; Hayashi, M.; Saruta, T. Anandamide Decreases Glomerular Filtration Rate through Predominant Vasodilation of Efferent Arterioles in Rat Kidneys. J. Am. Soc. Nephrol. 2004, 15, 1488. [Google Scholar] [CrossRef]
- Lin, C.-L.; Hsu, Y.-C.; Lee, P.-H.; Lei, C.-C.; Wang, J.-Y.; Huang, Y.-T.; Wang, S.-Y.; Wang, F.-S. Cannabinoid Receptor 1 Disturbance of PPARγ2 Augments Hyperglycemia Induction of Mesangial Inflammation and Fibrosis in Renal Glomeruli. J. Mol. Med. 2014, 92, 779–792. [Google Scholar] [CrossRef]
- Lecru, L.; Desterke, C.; Grassin-Delyle, S.; Chatziantoniou, C.; Vandermeersch, S.; Devocelle, A.; Vernochet, A.; Ivanovski, N.; Ledent, C.; Ferlicot, S.; et al. Cannabinoid Receptor 1 Is a Major Mediator of Renal Fibrosis. Kidney Int. 2015, 88, 72–84. [Google Scholar] [CrossRef]
- Oltrabella, F.; Melgoza, A.; Nguyen, B.; Guo, S. Role of the Endocannabinoid System in Vertebrates: Emphasis on the Zebrafish Model. Dev. Growth Differ. 2017, 59, 194–210. [Google Scholar] [CrossRef]
- Liu, L.Y.; Alexa, K.; Cortes, M.; Schatzman-Bone, S.; Kim, A.J.; Mukhopadhyay, B.; Cinar, R.; Kunos, G.; North, T.E.; Goessling, W. Cannabinoid Receptor Signaling Regulates Liver Development and Metabolism. Development 2016, 143, 609–622. [Google Scholar] [CrossRef]
- Tam, J. The Emerging Role of the Endocannabinoid System in the Pathogenesis and Treatment of Kidney Diseases. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 267–276. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Baker, S.; Rodriguez, J.-M.; Arceri, L.; Wingert, R.A. Using Zebrafish to Study Multiciliated Cell Development and Disease States. Cells 2024, 13, 1749. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.K.; Petrikas, M.; Chambers, B.E.; Wingert, R.A. Principles of Zebrafish Nephron Segment Development. J. Dev. Biol. 2023, 11, 14. [Google Scholar] [CrossRef]
- Bertram, J.F.; Douglas-Denton, R.N.; Diouf, B.; Hughson, M.D.; Hoy, W.E. Human Nephron Number: Implications for Health and Disease. Pediatr. Nephrol. 2011, 26, 1529–1533. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.L.; Suzuki, Y. Ciliated Human Renal Proximal Tubular Cells. Observations in Three Cases of Hypercalcemia. Am. J. Pathol. 1968, 53, 609. [Google Scholar]
- Larsen, T.E.; Ghadially, F.N. Cilia in Lupus Nephritis. J. Pathol. 1974, 114, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Katz, S.M.; Morgan, J.J. Cilia in the Human Kidney. Ultrastruct. Pathol. 1984, 6, 285–294. [Google Scholar] [CrossRef]
- Lungarella, G.; de Santi, M.M.; Tosi, P. Ultrastructural Study of the Ciliated Cells from Renal Tubular Epithelium in Acute Progressive Glomerulonephritis. Ultrastruct. Pathol. 1984, 6, 1–7. [Google Scholar] [CrossRef]
- Eymael, J.; Willemsen, B.; Xu, J.; Mooren, F.; Steenbergen, E.; Wetzels, J.F.; Dijkman, H.; Jansen, J.; Van der Vlag, J.; Smeets, B. Motile Cilia on Kidney Proximal Tubular Epithelial Cells Are Associated With Tubular Injury and Interstitial Fibrosis. Front. Cell Dev. Biol. 2022, 10, 765887. [Google Scholar] [CrossRef]
- Arévalo-Martín, Á.; García-Ovejero, D.; Rubio-Araiz, A.; Gómez, O.; Molina-Holgado, F.; Molina-Holgado, E. Cannabinoids Modulate Olig2 and Polysialylated Neural Cell Adhesion Molecule Expression in the Subventricular Zone of Post-Natal Rats through Cannabinoid Receptor 1 and Cannabinoid Receptor 2. Eur. J. Neurosci. 2007, 26, 1548–1559. [Google Scholar] [CrossRef]
- Pirone, A.; Lenzi, C.; Briganti, A.; Abbate, F.; Levanti, M.; Abramo, F.; Miragliotta, V. Spatial Distribution of Cannabinoid Receptor 1 and Fatty Acid Amide Hydrolase in the Cat Ovary and Oviduct. Acta Histochem. 2017, 119, 417–422. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.L.; Wingert, R.A.; Thisse, C.; Thisse, B.; Zon, L.I. Combinatorial Regulation of Novel Erythroid Gene Expression in Zebrafish. Exp. Hematol. 2008, 36, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Lengerke, C.; Wingert, R.; Beeretz, M.; Grauer, M.; Schmidt, A.G.; Konantz, M.; Daley, G.Q.; Davidson, A.J. Interactions between Cdx Genes and Retinoic Acid Modulate Early Cardiogenesis. Dev. Biol. 2011, 354, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.N.; Li, Y.; Marra, A.N.; Verdun, V.; Wingert, R.A. Flat Mount Preparation for Observation and Analysis of Zebrafish Embryo Specimens Stained by Whole Mount In Situ Hybridization. J. Vis. Exp. JoVE 2014, e51604. [Google Scholar] [CrossRef]
- Wingert, R.A.; Selleck, R.; Yu, J.; Song, H.-D.; Chen, Z.; Song, A.; Zhou, Y.; Thisse, B.; Thisse, C.; McMahon, A.P.; et al. The Cdx Genes and Retinoic Acid Control the Positioning and Segmentation of the Zebrafish Pronephros. PLoS Genet. 2007, 3, 1922–1938. [Google Scholar] [CrossRef]
- O’Brien, L.L.; Grimaldi, M.; Kostun, Z.; Wingert, R.A.; Selleck, R.; Davidson, A.J. Wt1a, Foxc1a, and the Notch Mediator Rbpj Physically Interact and Regulate the Formation of Podocytes in Zebrafish. Dev. Biol. 2011, 358, 318–330. [Google Scholar] [CrossRef]
- Gerlach, G.F.; Wingert, R.A. Zebrafish Pronephros Tubulogenesis and Epithelial Identity Maintenance Are Reliant on the Polarity Proteins Prkc Iota and Zeta. Dev. Biol. 2014, 396, 183–200. [Google Scholar] [CrossRef]
- Watson, S.; Chambers, D.; Hobbs, C.; Doherty, P.; Graham, A. The Endocannabinoid Receptor, CB1, Is Required for Normal Axonal Growth and Fasciculation. Mol. Cell. Neurosci. 2008, 38, 89–97. [Google Scholar] [CrossRef]
- Zuccarini, G.; D’Atri, I.; Cottone, E.; Mackie, K.; Shainer, I.; Gothilf, Y.; Provero, P.; Bovolin, P.; Merlo, G.R. Interference with the Cannabinoid Receptor CB1R Results in Miswiring of GnRH3 and AgRP1 Axons in Zebrafish Embryos. Int. J. Mol. Sci. 2020, 21, 168. [Google Scholar] [CrossRef]
- Zhou, F.; Rayamajhi, D.; Ravi, V.; Narasimhan, V.; Chong, Y.L.; Lu, H.; Venkatesh, B.; Roy, S. Conservation as Well as Divergence in Mcidas Function Underlies the Differentiation of Multiciliated Cells in Vertebrates. Dev. Biol. 2020, 465, 168–177. [Google Scholar] [CrossRef]
- Chambers, J.M.; Addiego, A.; Flores-Mireles, A.L.; Wingert, R.A. Ppargc1a Controls Ciliated Cell Development by Regulating Prostaglandin Biosynthesis. Cell Rep. 2020, 33, 108370. [Google Scholar] [CrossRef] [PubMed]
- Wesselman, H.M.; Flores-Mireles, A.L.; Bauer, A.; Pei, L.; Wingert, R.A. Esrrγa Regulates Nephron and Ciliary Development by Controlling Prostaglandin Synthesis. Development 2023, 150, dev201411. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Rodriguez, J.-M.; Wesselman, H.M.; Wingert, R.A. Emx2 Is an Essential Regulator of Ciliated Cell Development across Embryonic Tissues. iScience 2024, 27, 111271. [Google Scholar] [CrossRef]
- Poureetezadi, S.J.; Cheng, C.N.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Prostaglandin Signaling Regulates Nephron Segment Patterning of Renal Progenitors during Zebrafish Kidney Development. eLife 2016, 5, e17551. [Google Scholar] [CrossRef] [PubMed]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Abadji, V.; Lin, S.; Taha, G.; Griffin, G.; Stevenson, L.A.; Pertwee, R.G.; Makriyannis, A. (R)-Methanandamide: A Chiral Novel Anandamide Possessing Higher Potency and Metabolic Stability. J. Med. Chem. 1994, 37, 1889–1893. [Google Scholar] [CrossRef] [PubMed]
- Seely, K.A.; Brents, L.K.; Franks, L.N.; Rajasekaran, M.; Zimmerman, S.M.; Fantegrossi, W.E.; Prather, P.L. AM-251 and Rimonabant Act as Direct Antagonists at Mu-Opioid Receptors: Implications for Opioid/Cannabinoid Interaction Studies. Neuropharmacology 2012, 63, 905–915. [Google Scholar] [CrossRef]
- Wesselman, H.M.; Nguyen, T.K.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Advances in Understanding the Genetic Mechanisms of Zebrafish Renal Multiciliated Cell Development. J. Dev. Biol. 2023, 11, 1. [Google Scholar] [CrossRef]
- Wang, L.; Fu, C.; Fan, H.; Du, T.; Dong, M.; Chen, Y.; Jin, Y.; Zhou, Y.; Deng, M.; Gu, A.; et al. miR-34b Regulates Multiciliogenesis during Organ Formation in Zebrafish. Development 2013, 140, 2755–2764. [Google Scholar] [CrossRef]
- Zhou, F.; Narasimhan, V.; Shboul, M.; Chong, Y.L.; Reversade, B.; Roy, S. Gmnc Is a Master Regulator of the Multiciliated Cell Differentiation Program. Curr. Biol. 2015, 25, 3267–3273. [Google Scholar] [CrossRef]
- Pathak, N.; Obara, T.; Mangos, S.; Liu, Y.; Drummond, I.A. The Zebrafish Fleer Gene Encodes an Essential Regulator of Cilia Tubulin Polyglutamylation. MBoC 2007, 18, 4353–4364. [Google Scholar] [CrossRef] [PubMed]
- Delaval, B.; Covassin, L.; Lawson, N.D.; Doxsey, S. Centrin Depletion Causes Cyst Formation and Other Ciliopathy-Related Phenotypes in Zebrafish. Cell Cycle 2011, 10, 3964–3972. [Google Scholar] [CrossRef] [PubMed]
- Chong, Y.L.; Zhang, Y.; Zhou, F.; Roy, S. Distinct Requirements of E2f4 versus E2f5 Activity for Multiciliated Cell Development in the Zebrafish Embryo. Dev. Biol. 2018, 443, 165–172. [Google Scholar] [CrossRef]
- Jin, D.; Ni, T.T.; Sun, J.; Wan, H.; Amack, J.D.; Yu, G.; Fleming, J.; Chiang, C.; Li, W.; Papierniak, A.; et al. Prostaglandin Signaling Regulates Ciliogenesis by Modulating Intraflagellar Transport. Nat. Cell Biol. 2014, 16, 841–851. [Google Scholar] [CrossRef]
- Ramos, B.C.R.; Moraes, M.N.C.M.; Poletini, M.O.; Lima, L.H.R.G.; Castrucci, A.M.L. From Blue Light to Clock Genes in Zebrafish ZEM-2S Cells. PLoS ONE 2014, 9, e106252. [Google Scholar] [CrossRef]
- Dell, K.M. The Role of Cilia in the Pathogenesis of Cystic Kidney Disease. Curr. Opin. Pediatr. 2015, 27, 212–218. [Google Scholar] [CrossRef]
- Ma, M. Cilia and Polycystic Kidney Disease. Semin. Cell Dev. Biol. 2021, 110, 139–148. [Google Scholar] [CrossRef] [PubMed]
- McConnachie, D.J.; Stow, J.L.; Mallett, A.J. Ciliopathies and the Kidney: A Review. Am. J. Kidney Dis. 2021, 77, 410–419. [Google Scholar] [CrossRef]
- Marra, A.N.; Adeeb, B.D.; Chambers, B.E.; Drummond, B.E.; Ulrich, M.; Addiego, A.; Springer, M.; Poureetezadi, S.J.; Chambers, J.M.; Ronshaugen, M.; et al. Prostaglandin Signaling Regulates Renal Multiciliated Cell Specification and Maturation. Proc. Natl. Acad. Sci. USA 2019, 116, 8409–8418. [Google Scholar] [CrossRef]
- Park, F.; Potukuchi, P.K.; Moradi, H.; Kovesdy, C.P. Cannabinoids and the Kidney: Effects in Health and Disease. Am. J. Physiol. -Ren. Physiol. 2017, 313, F1124–F1132. [Google Scholar] [CrossRef]
- Chua, J.T.; Argueta, D.A.; DiPatrizio, N.V.; Kovesdy, C.P.; Vaziri, N.D.; Kalantar-Zadeh, K.; Moradi, H. Endocannabinoid System and the Kidneys: From Renal Physiology to Injury and Disease. Cannabis Cannabinoid Res. 2019, 4, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Jourdan, T.; Szanda, G.; Rosenberg, A.Z.; Tam, J.; Earley, B.J.; Godlewski, G.; Cinar, R.; Liu, Z.; Liu, J.; Ju, C.; et al. Overactive Cannabinoid 1 Receptor in Podocytes Drives Type 2 Diabetic Nephropathy. Proc. Natl. Acad. Sci. USA 2014, 111, E5420–E5428. [Google Scholar] [CrossRef]
- Udi, S.; Hinden, L.; Ahmad, M.; Drori, A.; Iyer, M.R.; Cinar, R.; Herman-Edelstein, M.; Tam, J. Dual Inhibition of Cannabinoid CB1 Receptor and Inducible NOS Attenuates Obesity-Induced Chronic Kidney Disease. Br. J. Pharmacol. 2020, 177, 110–127. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.H.; Lee, M.H.; Kim, J.E.; Song, H.K.; Kang, Y.S.; Lee, J.E.; Kim, H.W.; Cha, J.J.; Hyun, Y.Y.; Kim, S.H.; et al. Blockade of Cannabinoid Receptor 1 Improves Insulin Resistance, Lipid Metabolism, and Diabetic Nephropathy in Db/Db Mice. Endocrinology 2012, 153, 1387–1396. [Google Scholar] [CrossRef]
- Barutta, F.; Corbelli, A.; Mastrocola, R.; Gambino, R.; Di Marzo, V.; Pinach, S.; Rastaldi, M.P.; Perin, P.C.; Gruden, G. Cannabinoid Receptor 1 Blockade Ameliorates Albuminuria in Experimental Diabetic Nephropathy. Diabetes 2010, 59, 1046–1054. [Google Scholar] [CrossRef]
- Jenkin, K.A.; McAinch, A.J.; Zhang, Y.; Kelly, D.J.; Hryciw, D.H. Elevated Cannabinoid Receptor 1 and G Protein-Coupled Receptor 55 Expression in Proximal Tubule Cells and Whole Kidney Exposed to Diabetic Conditions. Clin. Exp. Pharmacol. Physiol. 2015, 42, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.c.; Lim, S.k.; Han, H.j.; Park, S.h. Cannabinoid Receptor 1 Mediates Palmitic Acid-Induced Apoptosis via Endoplasmic Reticulum Stress in Human Renal Proximal Tubular Cells. J. Cell. Physiol. 2010, 225, 654–663. [Google Scholar] [CrossRef]
- Janiak, P.; Poirier, B.; Bidouard, J.-P.; Cadrouvele, C.; Pierre, F.; Gouraud, L.; Barbosa, I.; Dedio, J.; Maffrand, J.-P.; Le Fur, G.; et al. Blockade of Cannabinoid CB1 Receptors Improves Renal Function, Metabolic Profile, and Increased Survival of Obese Zucker Rats. Kidney Int. 2007, 72, 1345–1357. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, T.; Dou, Z.; Wang, M.; Hu, Z.; Wang, B. CB1 Receptor Antagonist Rimonabant Protects against Chronic Intermittent Hypoxia-Induced Renal Injury in Rats. BMC Nephrol. 2021, 22, 153. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Pan, H.; Rajesh, M.; Bátkai, S.; Patel, V.; Harvey-White, J.; Mukhopadhyay, B.; Haskó, G.; Gao, B.; Mackie, K.; et al. CB1 Cannabinoid Receptors Promote Oxidative/Nitrosative Stress, Inflammation and Cell Death in a Murine Nephropathy Model. Br. J. Pharmacol. 2010, 160, 657–668. [Google Scholar] [CrossRef]
- Mick, D.U.; Rodrigues, R.B.; Leib, R.D.; Adams, C.M.; Chien, A.S.; Gygi, S.P.; Nachury, M.V. Proteomics of Primary Cilia by Proximity Labeling. Dev. Cell 2015, 35, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, V.A.; Mayrhofer, J.E.; Ilouz, R.; Tschaikner, P.; Raffeiner, P.; Röck, R.; Courcelles, M.; Apelt, F.; Lu, T.-W.; Baillie, G.S.; et al. Gpr161 Anchoring of PKA Consolidates GPCR and cAMP Signaling. Proc. Natl. Acad. Sci. USA 2016, 113, 7786–7791. [Google Scholar] [CrossRef] [PubMed]
- Hilgendorf, K.I.; Johnson, C.T.; Jackson, P.K. The Primary Cilium as a Cellular Receiver: Organizing Ciliary GPCR Signaling. Curr. Opin. Cell Biol. 2016, 39, 84–92. [Google Scholar] [CrossRef]
- Wachten, D.; Mick, D.U. Signal Transduction in Primary Cilia—Analyzing and Manipulating GPCR and Second Messenger Signaling. Pharmacol. Ther. 2021, 224, 107836. [Google Scholar] [CrossRef]
- Mykytyn, K.; Askwith, C. G-Protein-Coupled Receptor Signaling in Cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028183. [Google Scholar] [CrossRef]
- Somatilaka, B.N.; Hwang, S.-H.; Palicharla, V.R.; White, K.A.; Badgandi, H.; Shelton, J.M.; Mukhopadhyay, S. Ankmy2 Prevents Smoothened-Independent Hyperactivation of the Hedgehog Pathway via Cilia-Regulated Adenylyl Cyclase Signaling. Dev. Cell 2020, 54, 710–726.e8. [Google Scholar] [CrossRef]
- Siljee, J.E.; Wang, Y.; Bernard, A.A.; Ersoy, B.A.; Zhang, S.; Marley, A.; Von Zastrow, M.; Reiter, J.F.; Vaisse, C. Subcellular Localization of MC4R with ADCY3 at Neuronal Primary Cilia Underlies a Common Pathway for Genetic Predisposition to Obesity. Nat. Genet. 2018, 50, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bernard, A.; Comblain, F.; Yue, X.; Paillart, C.; Zhang, S.; Reiter, J.F.; Vaisse, C. Melanocortin 4 Receptor Signals at the Neuronal Primary Cilium to Control Food Intake and Body Weight. J. Clin. Investig. 2021, 131, e142064. [Google Scholar] [CrossRef]
- Mehta, Y.R.; Lewis, S.A.; Leo, K.T.; Chen, L.; Park, E.; Raghuram, V.; Chou, C.-L.; Yang, C.-R.; Kikuchi, H.; Khundmiri, S.; et al. “ADPKD-Omics”: Determinants of Cyclic AMP Levels in Renal Epithelial Cells. Kidney Int. 2022, 101, 47–62. [Google Scholar] [CrossRef]
- Paolocci, E.; Zaccolo, M. Compartmentalised cAMP Signalling in the Primary Cilium. Front. Physiol. 2023, 14, 1187134. [Google Scholar] [CrossRef]
- Hansen, J.N.; Kaiser, F.; Leyendecker, P.; Stüven, B.; Krause, J.; Derakhshandeh, F.; Irfan, J.; Sroka, T.J.; Preval, K.M.; Desai, P.B.; et al. A cAMP Signalosome in Primary Cilia Drives Gene Expression and Kidney Cyst Formation. EMBO Rep. 2022, 23, e54315. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Majeed, S.; Moloney, B.C.; Nauli, S.M. Mechanisms Regulating Cilia Growth and Cilia Function in Endothelial Cells. Cell. Mol. Life Sci. 2012, 69, 165–173. [Google Scholar] [CrossRef]
- Pei, S.-J.; Zhu, H.-Y.; Guo, J.-H.; Zhang, X.; Deng, Z.-J. Knockout of CNR1 Prevents Metabolic Stress-Induced Cardiac Injury through Improving Insulin Resistance (IR) Injury and Endoplasmic Reticulum (ER) Stress by Promoting AMPK-Alpha Activation. Biochem. Biophys. Res. Commun. 2018, 503, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Boals, A.G.; Collier, D.M.; Romero, J.R.; Hillard, C.J.; Park, F. Lack of Cannabinoid Type 2 Promoter Activity in Normal or Injured Kidneys Using a Cnr2-GFP Reporter Mouse. Cannabis Cannabinoid Res. 2024, 10, 400–408. [Google Scholar] [CrossRef]
- Zhou, S.; Wu, Q.; Lin, X.; Ling, X.; Miao, J.; Liu, X.; Hu, C.; Zhang, Y.; Jia, N.; Hou, F.F.; et al. Cannabinoid Receptor Type 2 Promotes Kidney Fibrosis through Orchestrating β-Catenin Signaling. Kidney Int. 2021, 99, 364–381. [Google Scholar] [CrossRef]
- Zhou, S.; Ling, X.; Liang, Y.; Feng, Q.; Xie, C.; Li, J.; Chen, Q.; Miao, J.; Zhang, M.; Li, Z.; et al. Cannabinoid Receptor 2 Plays a Key Role in Renal Fibrosis through Inhibiting Lipid Metabolism in Renal Tubular Cells. Metabolism 2024, 159, 155978. [Google Scholar] [CrossRef]
- Rezende, B.; Alencar, A.K.N.; de Bem, G.F.; Fontes-Dantas, F.L.; Montes, G.C. Endocannabinoid System: Chemical Characteristics and Biological Activity. Pharmaceuticals 2023, 16, 148. [Google Scholar] [CrossRef] [PubMed]
- Imperatore, R.; D’Angelo, L.; Safari, O.; Motlagh, H.A.; Piscitelli, F.; de Girolamo, P.; Cristino, L.; Varricchio, E.; di Marzo, V.; Paolucci, M. Overlapping Distribution of Orexin and Endocannabinoid Receptors and Their Functional Interaction in the Brain of Adult Zebrafish. Front. Neuroanat. 2018, 12, 62. [Google Scholar] [CrossRef]
- Zhou, S.; Ling, X.; Meng, P.; Liang, Y.; Shen, K.; Wu, Q.; Zhang, Y.; Chen, Q.; Chen, S.; Liu, Y.; et al. Cannabinoid Receptor 2 Plays a Central Role in Renal Tubular Mitochondrial Dysfunction and Kidney Ageing. J. Cell. Mol. Med. 2021, 25, 8957–8972. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Rajesh, M.; Pan, H.; Patel, V.; Mukhopadhyay, B.; Bátkai, S.; Gao, B.; Haskó, G.; Pacher, P. Cannabinoid-2 Receptor Limits Inflammation, Oxidative/Nitrosative Stress, and Cell Death in Nephropathy. Free Radic. Biol. Med. 2010, 48, 457–467. [Google Scholar] [CrossRef]
- Swanson, M.L.; Regner, K.R.; Moore, B.M.; Park, F. Cannabinoid Type 2 Receptor Activation Reduces the Progression of Kidney Fibrosis Using a Mouse Model of Unilateral Ureteral Obstruction. Cannabis Cannabinoid Res. 2022, 7, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Rokicki, M.; Żurowski, J.; Sawicki, S.; Ocłoń, E.; Szmatoła, T.; Jasielczuk, I.; Mizera-Szpilka, K.; Semik-Gurgul, E.; Gurgul, A. Impact of Long-Term Cannabidiol (CBD) Treatment on Mouse Kidney Transcriptome. Genes. 2024, 15, 1640. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Cao, X.; Zhang, K.; Li, Y.; Zheng, Q.; Li, G.; He, Q.; Li, S.; Xu, G.; Zhang, K. Celastrol Alleviates Renal Fibrosis by Upregulating Cannabinoid Receptor 2 Expression. Cell Death Dis. 2018, 9, 601. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.R.; Ahmed, K.T.; Ali, D.W. Cannabinoid Receptor 2 (Cb2r) Mediates Cannabinol (CBN) Induced Developmental Defects in Zebrafish. Sci. Rep. 2022, 12, 20251. [Google Scholar] [CrossRef]
- Colón-Cruz, L.; Rodriguez-Morales, R.; Santana-Cruz, A.; Cantres-Velez, J.; Torrado-Tapias, A.; Lin, S.-J.; Yudowski, G.; Kensler, R.; Marie, B.; Burgess, S.M.; et al. Cnr2 Is Important for Ribbon Synapse Maturation and Function in Hair Cells and Photoreceptors. Front. Mol. Neurosci. 2021, 14, 624265. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.K.; Baker, S.; Angtuaco, J.; Arceri, L.; Kaczor, S.; Fitzsimonds, B.; Hawkins, M.R.; Wingert, R.A. Cannabinoid Receptor 1 Regulates Zebrafish Renal Multiciliated Cell Development via cAMP Signaling. J. Dev. Biol. 2025, 13, 20. https://doi.org/10.3390/jdb13020020
Nguyen TK, Baker S, Angtuaco J, Arceri L, Kaczor S, Fitzsimonds B, Hawkins MR, Wingert RA. Cannabinoid Receptor 1 Regulates Zebrafish Renal Multiciliated Cell Development via cAMP Signaling. Journal of Developmental Biology. 2025; 13(2):20. https://doi.org/10.3390/jdb13020020
Chicago/Turabian StyleNguyen, Thanh Khoa, Sophia Baker, Julienne Angtuaco, Liana Arceri, Samuel Kaczor, Bram Fitzsimonds, Matthew R. Hawkins, and Rebecca A. Wingert. 2025. "Cannabinoid Receptor 1 Regulates Zebrafish Renal Multiciliated Cell Development via cAMP Signaling" Journal of Developmental Biology 13, no. 2: 20. https://doi.org/10.3390/jdb13020020
APA StyleNguyen, T. K., Baker, S., Angtuaco, J., Arceri, L., Kaczor, S., Fitzsimonds, B., Hawkins, M. R., & Wingert, R. A. (2025). Cannabinoid Receptor 1 Regulates Zebrafish Renal Multiciliated Cell Development via cAMP Signaling. Journal of Developmental Biology, 13(2), 20. https://doi.org/10.3390/jdb13020020