
Ribosome Incorporation Transdifferentiates Chick Primary Cells and Induces Their Proliferation by Secreting Growth Factors

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Chick Muscle-Derived Cells
2.2. Isolation and Purification of Ribosomes
2.3. Ribosome-Induced Cell Clusters (RICs) Formation of CMCs
2.4. Immunocytochemistry
2.5. Alkaline Phosphatase Staining
2.6. Transdifferentiation of RICs-CMC
2.7. Proliferation Assay
2.8. RNA-Sequencing (RNA-Seq)
2.9. Statistical Analysis
3. Results
3.1. Incorporation of Ribosomes into Chick Muscle-Derived Cells Forms Cell Clusters
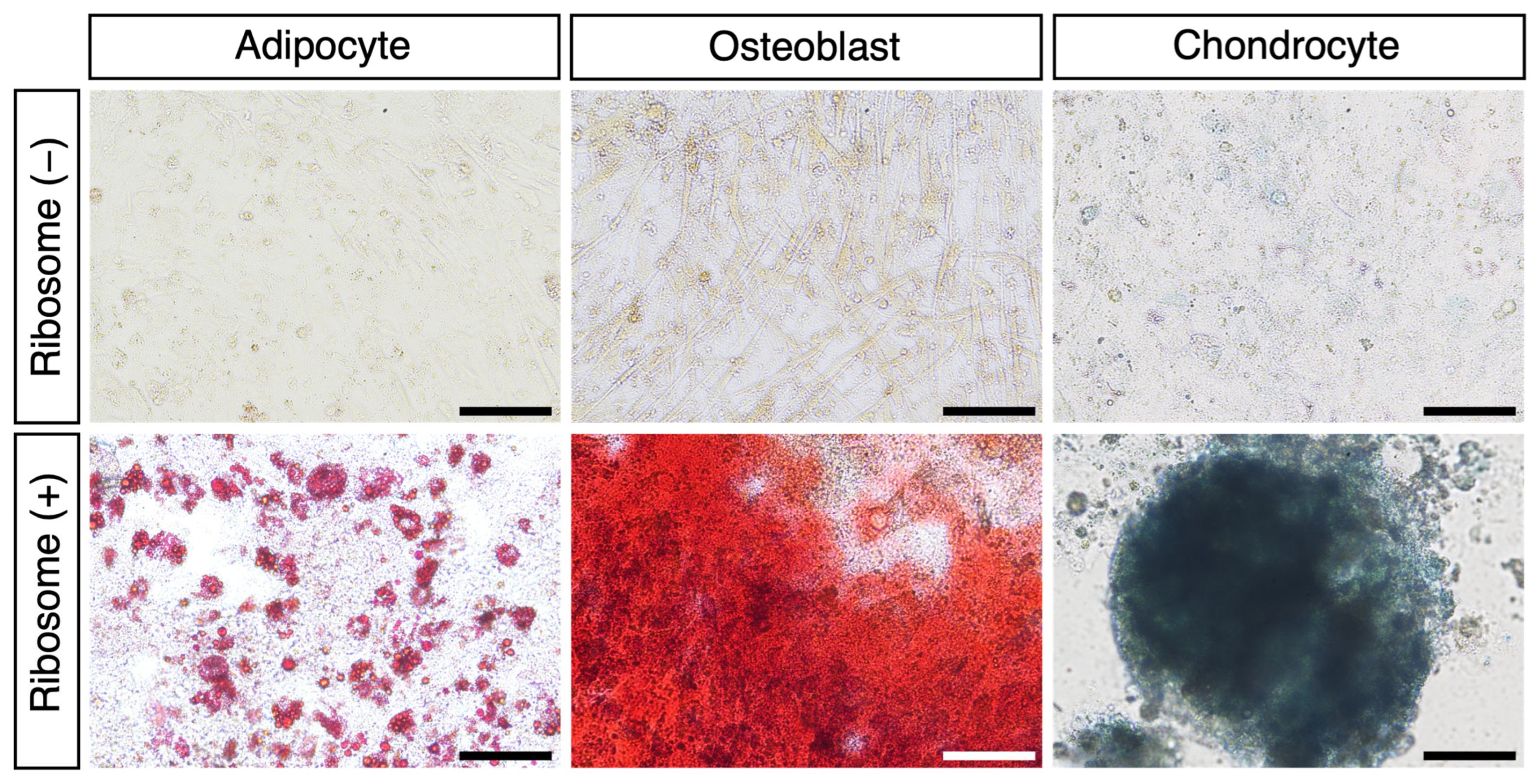
3.2. RICs-CMC into Mesodermal Lineage Cells In Vitro
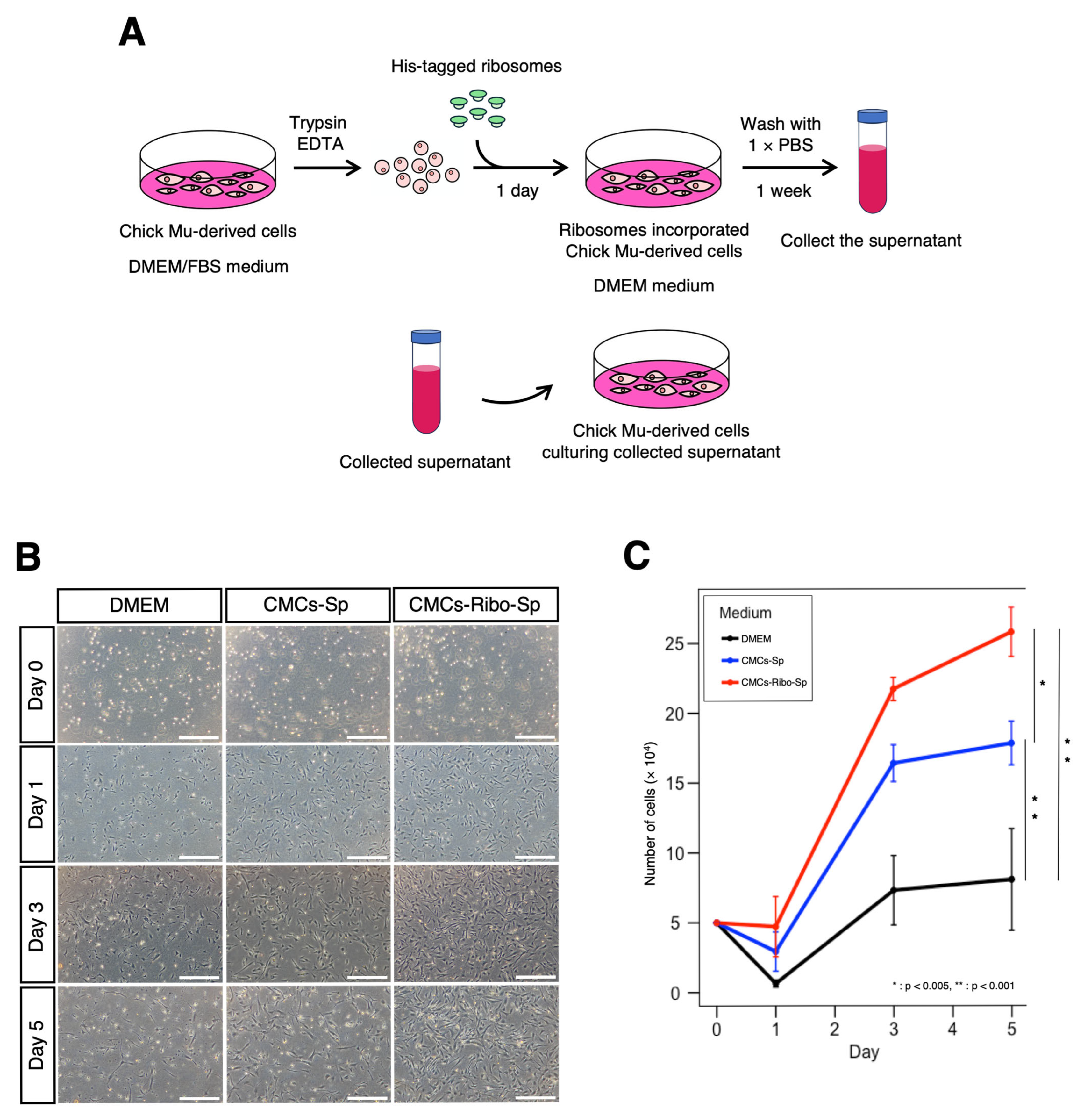
3.3. Cell Culture Supernatant from Ribosome-Incorporated CMCs Enhances Cell Proliferation
3.4. Ribosome-Incorporated CMCs Express Several Cell Growth Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martinez, J.; Dabert, P.; Barrington, S.; Burton, C. Livestock Waste Treatment Systems for Environmental Quality, Food Safety, and Sustainability. Bioresour. Technol. 2009, 100, 5527–5536. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Bahamondes, P.; Oba, M.; Beauchemin, K.A. Universally Applicable Methane Prediction Equations for Beef Cattle Fed High- or Low-Forage Diets. Can. J. Anim. Sci. 2017, 97, 83–94. [Google Scholar] [CrossRef]
- Forster, P.; Ramaswamy, V.; Artaxo, P.; Berntsen, T.; Betts, R.; Fahey, D.W.; Haywood, J.; Lean, J.; Lowe, D.C.; Raga, G.; et al. Changes in Atmospheric Constituents and in Radiative Forcing; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Shukla, P.R.; Skea, J.; Reisinger, A.; IPCC (Eds.) Climate Change 2022: Mitigation of Climate Change; IPCC: Geneva, Switzerland, 2022; ISBN 978-92-9169-160-9. [Google Scholar]
- Poore, J.; Nemecek, T. Reducing Food’s Environmental Impacts through Producers and Consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Kawano, R.; Ito, N. Lactic Acid Bacteria Convert Human Fibroblasts to Multipotent Cells. PLoS ONE 2012, 7, e51866. [Google Scholar] [CrossRef]
- Ito, N.; Ohta, K. Reprogramming of Human Somatic Cells by Bacteria. Dev. Growth Differ. 2015, 57, 305–312. [Google Scholar] [CrossRef]
- Ito, N.; Katoh, K.; Kushige, H.; Saito, Y.; Umemoto, T.; Matsuzaki, Y.; Kiyonari, H.; Kobayashi, D.; Soga, M.; Era, T.; et al. Ribosome Incorporation into Somatic Cells Promotes Lineage Transdifferentiation towards Multipotency. Sci. Rep. 2018, 8, 1634. [Google Scholar] [CrossRef]
- Istiaq, A.; Ohta, K. Ribosome-Induced Cellular Multipotency, an Emerging Avenue in Cell Fate Reversal. Cells 2021, 10, 2276. [Google Scholar] [CrossRef]
- Istiaq, A.; Ishtiyaq Ahmad, S.A.; Anam, M.B.; Kudo, M.; Nakayama, S.; Ito, N.; Ohta, K. Chapter 9—Bacteria to Form Induced Pluripotent Stem Cells. In Methods in iPSC Technology; Birbrair, A., Ed.; Advances in Stem Cell Biology; Academic Press: New York, NY, USA, 2021; Volume 9, pp. 231–247. ISBN 978-0-323-85766-6. [Google Scholar]
- Datta, A.; Istiaq, A.; Tamura, S.; Ohta, K. Bacterial Ribosomes Induce Plasticity in Mouse Adult Fibroblasts. Cells 2024, 13, 1116. [Google Scholar] [CrossRef]
- Anam, M.B.; Istiaq, A.; Kariya, R.; Kudo, M.; Ishtiyaq Ahmad, S.A.; Ito, N.; Okada, S.; Ohta, K. Ribosome Induces Transdifferentiation of A549 and H-111-TC Cancer Cell Lines. Biochem. Biophys. Rep. 2021, 26, 100946. [Google Scholar] [CrossRef]
- Kudo, M.; Anam, M.B.; Istiaq, A.; Ahmad, S.A.I.; Ito, N.; Ohta, K. Ribosome Incorporation Induces EMT-like Phenomenon with Cell Cycle Arrest in Human Breast Cancer Cell. Cells Tissues Organs 2021, 211, 212–221. [Google Scholar] [CrossRef]
- Shirakawa, Y.; Hide, T.; Yamaoka, M.; Ito, Y.; Ito, N.; Ohta, K.; Shinojima, N.; Mukasa, A.; Saito, H.; Jono, H. Ribosomal Protein S6 Promotes Stem-like Characters in Glioma Cells. Cancer Sci. 2020, 111, 2041–2051. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, Y.; Ohta, K.; Miyake, S.; Kanemaru, A.; Kuwano, A.; Yonemaru, K.; Uchino, S.; Yamaoka, M.; Ito, Y.; Ito, N.; et al. Glioma Cells Acquire Stem-like Characters by Extrinsic Ribosome Stimuli. Cells 2021, 10, 2970. [Google Scholar] [CrossRef]
- Chen, L.; Guttieres, D.; Koenigsberg, A.; Barone, P.W.; Sinskey, A.J.; Springs, S.L. Large-Scale Cultured Meat Production: Trends, Challenges and Promising Biomanufacturing Technologies. Biomaterials 2022, 280, 121274. [Google Scholar] [CrossRef] [PubMed]
- Garrison, G.L.; Biermacher, J.T.; Brorsen, B.W. How Much Will Large-Scale Production of Cell-Cultured Meat Cost? J. Agric. Food Res. 2022, 10, 100358. [Google Scholar] [CrossRef]
- Benny, A.; Pandi, K.; Upadhyay, R. Techniques, Challenges and Future Prospects for Cell-Based Meat. Food Sci. Biotechnol. 2022, 31, 1225–1242. [Google Scholar] [CrossRef]
- Woelken, L.; Weckowska, D.M.; Dreher, C.; Rauh, C. Toward an Innovation Radar for Cultivated Meat: Exploring Process Technologies for Cultivated Meat and Claims about Their Social Impacts. Front. Sustain. Food Syst. 2024, 8, 1390720. [Google Scholar] [CrossRef]
- ABOUT US | IntegriCulture Inc. Available online: https://integriculture.com/en/aboutus/ (accessed on 23 May 2025).
- Liu, Q.; Xie, L.; Chen, W. Recombinant Porcine FGF1 Promotes Muscle Stem Cell Proliferation and Mitochondrial Function for Cultured Meat Production. J. Agric. Food Chem. 2025, 73, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Stout, A.J.; Mirliani, A.B.; Rittenberg, M.L.; Shub, M.; White, E.C.; Yuen, J.S.K.; Kaplan, D.L. Simple and Effective Serum-Free Medium for Sustained Expansion of Bovine Satellite Cells for Cell Cultured Meat. Commun. Biol. 2022, 5, 466. [Google Scholar] [CrossRef]
- Yamanaka, K.; Haraguchi, Y.; Takahashi, H.; Kawashima, I.; Shimizu, T. Development of Serum-Free and Grain-Derived-Nutrient-Free Medium Using Microalga-Derived Nutrients and Mammalian Cell-Secreted Growth Factors for Sustainable Cultured Meat Production. Sci. Rep. 2023, 13, 498. [Google Scholar] [CrossRef]
- Chu, S.; Haraguchi, Y.; Asahi, T.; Kato, Y.; Kondo, A.; Hasunuma, T.; Shimizu, T. A Serum-Free Culture Medium Production System by Co-Culture Combining Growth Factor-Secreting Cells and l-Lactate-Assimilating Cyanobacteria for Sustainable Cultured Meat Production. Sci. Rep. 2024, 14, 19578. [Google Scholar] [CrossRef]
- Ederth, J.; Mandava, C.S.; Dasgupta, S.; Sanyal, S. A Single-Step Method for Purification of Active His-Tagged Ribosomes from a Genetically Engineered Escherichia Coli. Nucleic Acids Res. 2009, 37, e15. [Google Scholar] [CrossRef] [PubMed]
- Chen, S. Ultrafast One-Pass FASTQ Data Preprocessing, Quality Control, and Deduplication Using Fastp. iMeta 2023, 2, e107. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- In, H.L.Y.; Pincket, R. Transcripts per Million Ratio: Applying Distribution-Aware Normalisation over the Popular TPM Method 2022. arXiv 2022, arXiv:2205.02844. [Google Scholar]
- Yan, J.; Yang, H.; Gan, L.; Sun, C. Adiponectin-Impaired Adipocyte Differentiation Negatively Regulates Fat Deposition in Chicken. J. Anim. Physiol. Anim. Nutr. 2014, 98, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Pei, W.; Wang, K.; Zhang, S.; Chen, F.; Wu, Y.; Guan, W. In Vitro Culture and Biological Properties of Broiler Adipose-derived Stem Cells. Exp. Ther. Med. 2018, 16, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Adhikari, R.; White, D.L.; Kim, W.K. Role of 1,25-Dihydroxyvitamin D3 on Osteogenic Differentiation and Mineralization of Chicken Mesenchymal Stem Cells. Front. Physiol. 2021, 12, 479596. [Google Scholar] [CrossRef]
- PiPE LiNE | IntegriCulture Inc. Available online: https://integriculture.com/en/service/pipeline/ (accessed on 23 May 2025).
- Ohuchi, H.; Nakagawa, T.; Yamamoto, A.; Araga, A.; Ohata, T.; Ishimaru, Y.; Yoshioka, H.; Kuwana, T.; Nohno, T.; Yamasaki, M.; et al. The Mesenchymal Factor, FGF10, Initiates and Maintains the Outgrowth of the Chick Limb Bud through Interaction with FGF8, an Apical Ectodermal Factor. Development 1997, 124, 2235–2244. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Sanchez, J.-C.; Gooley, A.A.; Appel, R.D.; Humphery-Smith, I.; Hochstrasser, D.F.; Williams, K.L. Progress with Proteome Projects: Why All Proteins Expressed by a Genome Should Be Identified and How To Do It. Biotechnol. Genet. Eng. Rev. 1996, 13, 19–50. [Google Scholar] [CrossRef] [PubMed]
- Clish, C.B. Metabolomics: An Emerging but Powerful Tool for Precision Medicine. Cold Spring Harb. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, S.; Istiaq, A.; Datta, A.; Lu, M.; Nakayama, S.; Takashi, K.; Nakajo, N.; Tamura, S.; Kawashima, I.; Ohta, K. Ribosome Incorporation Transdifferentiates Chick Primary Cells and Induces Their Proliferation by Secreting Growth Factors. J. Dev. Biol. 2025, 13, 19. https://doi.org/10.3390/jdb13020019
Inoue S, Istiaq A, Datta A, Lu M, Nakayama S, Takashi K, Nakajo N, Tamura S, Kawashima I, Ohta K. Ribosome Incorporation Transdifferentiates Chick Primary Cells and Induces Their Proliferation by Secreting Growth Factors. Journal of Developmental Biology. 2025; 13(2):19. https://doi.org/10.3390/jdb13020019
Chicago/Turabian StyleInoue, Shota, Arif Istiaq, Anamika Datta, Mengxue Lu, Shintaro Nakayama, Kousei Takashi, Nobushige Nakajo, Shigehiko Tamura, Ikko Kawashima, and Kunimasa Ohta. 2025. "Ribosome Incorporation Transdifferentiates Chick Primary Cells and Induces Their Proliferation by Secreting Growth Factors" Journal of Developmental Biology 13, no. 2: 19. https://doi.org/10.3390/jdb13020019
APA StyleInoue, S., Istiaq, A., Datta, A., Lu, M., Nakayama, S., Takashi, K., Nakajo, N., Tamura, S., Kawashima, I., & Ohta, K. (2025). Ribosome Incorporation Transdifferentiates Chick Primary Cells and Induces Their Proliferation by Secreting Growth Factors. Journal of Developmental Biology, 13(2), 19. https://doi.org/10.3390/jdb13020019