
The Immunoglobulin Superfamily Members syg-2 and syg-1 Regulate Neurite Development in C. elegans



Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Genetics
2.2. EMS Mutagenesis and Genome Resequencing
2.3. Imaging and Analysis
2.4. Data Availability
3. Results
3.1. syg-2 Mutations Cause Pdns
3.2. syg-2 and syg-1 Function in a Common Pathway in Parallel to Wnt Signaling
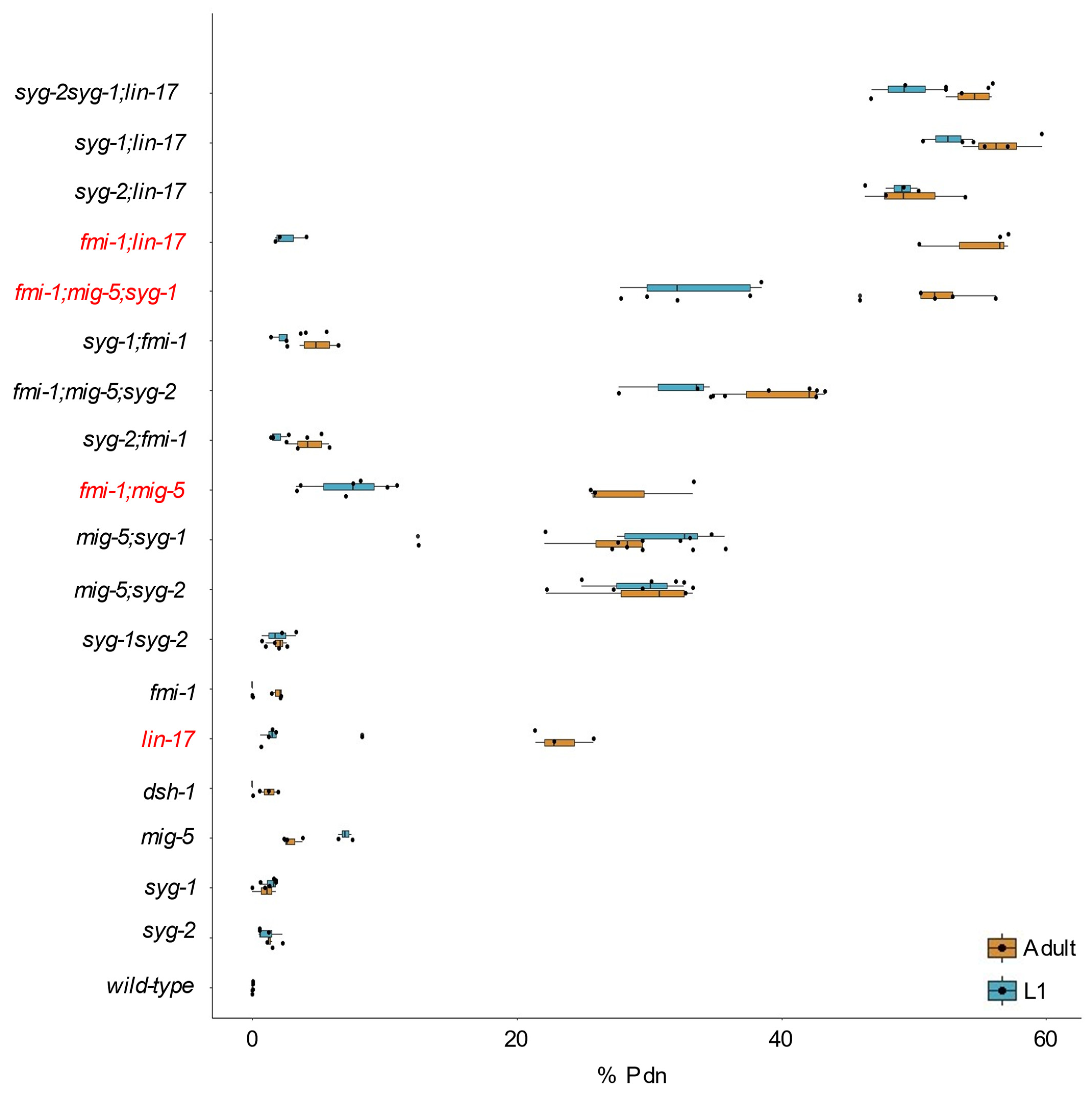
3.3. syg-2 and syg-1 Mutations are Additive with fmi-1
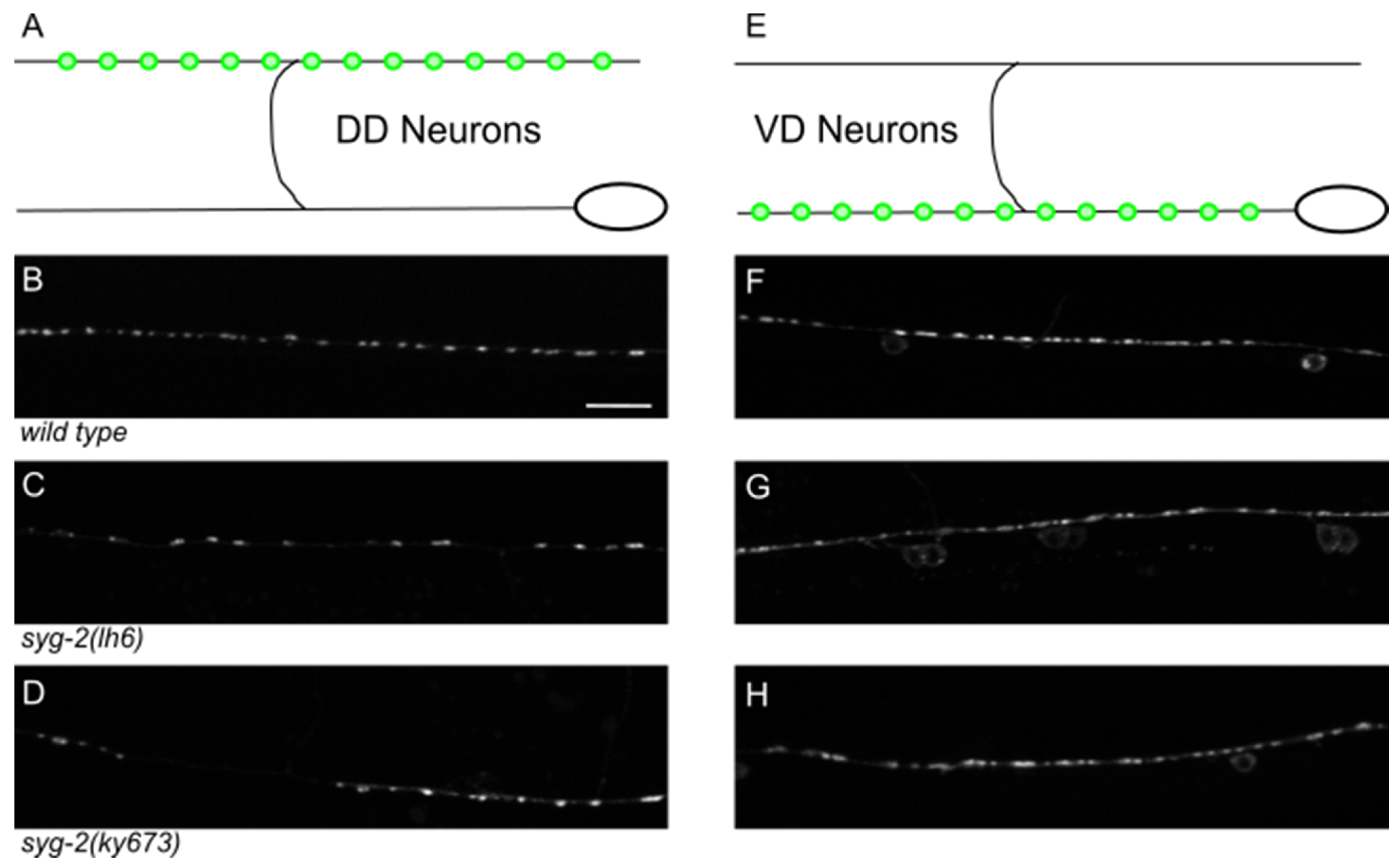
3.4. syg-1 and syg-2 Primarily Affect DD Neurons, While fmi-1 Affects VD Neurons
3.5. syg-2 and Wnt Signaling Mutations Result in Bipolar DD Neurons
3.6. syg-2 Mutations Result in Fewer DD Synapses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Najarro, E.H.; Ackley, B.D.C. Elegans fmi-1/flamingo and Wnt pathway components interact genetically to control the anteroposterior neurite growth of the VD GABAergic neurons. Dev. Biol. 2013, 377, 224–235. [Google Scholar] [CrossRef]
- Usui, T.; Shima, Y.; Shimada, Y.; Hirano, S.; Burgess, R.W.; Schwarz, T.L.; Takeichi, M.; Uemura, T. Flamingo, a seven-pass transmembrane cadherin, regulates planar cell polarity under the control of Frizzled. Cell 1999, 98, 585–595. [Google Scholar] [CrossRef]
- Liebscher, I.; Cevheroglu, O.; Hsiao, C.C.; Maia, A.F.; Schihada, H.; Scholz, N.; Soave, M.; Spiess, K.; Trajkovic, K.; Kosloff, M.; et al. A guide to adhesion GPCR research. FEBS J. 2021. [Google Scholar] [CrossRef]
- Zhou, L.; Bar, I.; Achouri, Y.; Campbell, K.; De Backer, O.; Hebert, J.M.; Jones, K.; Kessaris, N.; de Rouvroit, C.L.; O’Leary, D.; et al. Early forebrain wiring: Genetic dissection using conditional Celsr3 mutant mice. Science 2008, 320, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Usui, T.; Uemura, T.; Jan, L.; Jan, Y.N. Flamingo controls the planar polarity of sensory bristles and asymmetric division of sensory organ precursors in Drosophila. Curr. Biol. 1999, 9, 1247–1250. [Google Scholar] [CrossRef]
- Das, G.; Reynolds-Kenneally, J.; Mlodzik, M. The atypical cadherin Flamingo links Frizzled and Notch signaling in planar polarity establishment in the Drosophila eye. Dev. Cell 2002, 2, 655–666. [Google Scholar] [CrossRef]
- Chen, W.S.; Antic, D.; Matis, M.; Logan, C.Y.; Povelones, M.; Anderson, G.A.; Nusse, R.; Axelrod, J.D. Asymmetric homotypic interactions of the atypical cadherin flamingo mediate intercellular polarity signaling. Cell 2008, 133, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Mouri, K.; Horiuchi, S.Y.; Uemura, T. Cohesin controls planar cell polarity by regulating the level of the seven-pass transmembrane cadherin Flamingo. Genes Cells 2012, 17, 509–524. [Google Scholar] [CrossRef]
- Bao, H.; Berlanga, M.L.; Xue, M.; Hapip, S.M.; Daniels, R.W.; Mendenhall, J.M.; Alcantara, A.A.; Zhang, B. The atypical cadherin flamingo regulates synaptogenesis and helps prevent axonal and synaptic degeneration in Drosophila. Mol. Cell Neurosci. 2007, 34, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Hakeda-Suzuki, S.; Berger-Muller, S.; Tomasi, T.; Usui, T.; Horiuchi, S.Y.; Uemura, T.; Suzuki, T. Golden Goal collaborates with Flamingo in conferring synaptic-layer specificity in the visual system. Nat. Neurosci. 2011, 14, 314–323. [Google Scholar] [CrossRef]
- Najarro, E.H.; Wong, L.; Zhen, M.; Carpio, E.P.; Goncharov, A.; Garriga, G.; Lundquist, E.A.; Jin, Y.; Ackley, B.D. Caenorhabditis elegans flamingo cadherin fmi-1 regulates GABAergic neuronal development. J. Neurosci. 2012, 32, 4196–4211. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Huang, Y.; Feng, J.; Alvarez-Bolado, G.; Grove, E.A.; Yang, Y.; Tissir, F.; Zhou, L.; Goffinet, A.M. Genetic evidence that Celsr3 and Celsr2, together with Fzd3, regulate forebrain wiring in a Vangl-independent manner. Proc. Natl. Acad. Sci. USA 2014, 111, E2996–E3004. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, Q.; Li, C.; Yu, P.; Qu, Y.; Zhou, L. The role of Celsr3 in the development of central somatosensory projections from dorsal root ganglia. Neuroscience 2017, 359, 267–276. [Google Scholar] [CrossRef]
- Thakar, S.; Wang, L.; Yu, T.; Ye, M.; Onishi, K.; Scott, J.; Qi, J.; Fernandes, C.; Han, X.; Yates, J.R., 3rd; et al. Evidence for opposing roles of Celsr3 and Vangl2 in glutamatergic synapse formation. Proc. Natl. Acad. Sci. USA 2017, 114, E610–E618. [Google Scholar] [CrossRef]
- Chai, G.; Goffinet, A.M.; Tissir, F. Celsr3 and Fzd3 in axon guidance. Int. J. Biochem. Cell Biol. 2015, 64, 11–14. [Google Scholar] [CrossRef]
- Formstone, C.J.; Mason, I. Combinatorial activity of Flamingo proteins directs convergence and extension within the early zebrafish embryo via the planar cell polarity pathway. Dev. Biol. 2005, 282, 320–335. [Google Scholar] [CrossRef][Green Version]
- Shen, K.; Fetter, R.D.; Bargmann, C.I. Synaptic specificity is generated by the synaptic guidepost protein SYG-2 and its receptor, SYG-1. Cell 2004, 116, 869–881. [Google Scholar] [CrossRef]
- Shen, K.; Bargmann, C.I. The immunoglobulin superfamily protein SYG-1 determines the location of specific synapses in C. elegans. Cell 2003, 112, 619–630. [Google Scholar] [CrossRef]
- Chao, D.L.; Shen, K. Functional dissection of SYG-1 and SYG-2, cell adhesion molecules required for selective synaptogenesis in C. elegans. Mol. Cell Neurosci. 2008, 39, 248–257. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Gruning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed]
- Minevich, G.; Park, D.S.; Blankenberg, D.; Poole, R.J.; Hobert, O. CloudMap: A cloud-based pipeline for analysis of mutant genome sequences. Genetics 2012, 192, 1249–1269. [Google Scholar] [CrossRef] [PubMed]
- Ackley, B.D.; Kang, S.H.; Crew, J.R.; Suh, C.; Jin, Y.; Kramer, J.M. The basement membrane components nidogen and type XVIII collagen regulate organization of neuromuscular junctions in Caenorhabditis elegans. J. Neurosci 2003, 23, 3577–3587. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; R Foundation: Vienna, Austria, 2013. [Google Scholar]
- Jorgensen, E.M. Gaba. WormBook 2005, 1–13. [Google Scholar] [CrossRef]
- Eastman, C.; Horvitz, H.R.; Jin, Y. Coordinated transcriptional regulation of the unc-25 glutamic acid decarboxylase and the unc-47 GABA vesicular transporter by the Caenorhabditis elegans UNC-30 homeodomain protein. J. Neurosci. 1999, 19, 6225–6234. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Hoskins, R.; Horvitz, H.R. Control of type-D GABAergic neuron differentiation by C. elegans UNC-30 homeodomain protein. Nature 1994, 372, 780–783. [Google Scholar] [CrossRef]
- Huang, X.; Huang, P.; Robinson, M.K.; Stern, M.J.; Jin, Y. UNC-71, a disintegrin and metalloprotease (ADAM) protein, regulates motor axon guidance and sex myoblast migration in C. elegans. Development 2003, 130, 3147–3161. [Google Scholar] [CrossRef] [PubMed]
- Steimel, A.; Wong, L.; Najarro, E.H.; Ackley, B.D.; Garriga, G.; Hutter, H. The Flamingo ortholog FMI-1 controls pioneer-dependent navigation of follower axons in C. elegans. Development 2010, 137, 3663–3673. [Google Scholar] [CrossRef]
- Sawa, H.; Lobel, L.; Horvitz, H.R. The Caenorhabditis elegans gene lin-17, which is required for certain asymmetric cell divisions, encodes a putative seven-transmembrane protein similar to the Drosophila frizzled protein. Genes Dev. 1996, 10, 2189–2197. [Google Scholar] [CrossRef]
- Ackley, B.D.C. Elegans fmi-1/flamingo and Wnt pathway components interact genetically to control the anteroposterior neurite growth of the VD GABAergic neurons. Worm 2013, 2, e25715. [Google Scholar] [CrossRef]
- Wanner, N.; Noutsou, F.; Baumeister, R.; Walz, G.; Huber, T.B.; Neumann-Haefelin, E. Functional and spatial analysis of C. elegans SYG-1 and SYG-2, orthologs of the Neph/nephrin cell adhesion module directing selective synaptogenesis. PLoS ONE 2011, 6, e23598. [Google Scholar] [CrossRef] [PubMed]
- Kurland, M.; O’Meara, B.; Tucker, D.K.; Ackley, B.D. The Hox Gene egl-5 Acts as a Terminal Selector for VD13 Development via Wnt Signaling. J. Dev. Biol 2020, 8, 5. [Google Scholar] [CrossRef]
- Poon, V.Y.; Klassen, M.P.; Shen, K. UNC-6/netrin and its receptor UNC-5 locally exclude presynaptic components from dendrites. Nature 2008, 455, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Shen, K. WNTs in synapse formation and neuronal circuitry. EMBO J. 2012, 31, 2697–2704. [Google Scholar] [CrossRef] [PubMed]
- Hilliard, M.A.; Bargmann, C.I. Wnt signals and frizzled activity orient anterior-posterior axon outgrowth in C. elegans. Dev. Cell 2006, 10, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Durbin, R.M. Studies on the Development and Organisation of the Nervous System of Caenorhabditis Elegans; Kings College: Cambridge, UK, 1987. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Pdns | Total | Average | St. Dev. | Additive a | p Value (Comparison) |
|---|---|---|---|---|---|---|
| wild-type | 0 | 362 | 0.0% | 0.0% | ND | |
| mig-5(rh94) | 9 | 323 | 2.9% | 0.7% | ND | 0.011 (wt) |
| syg-2(lh6) | 7 | 504 | 1.3% | 0.2% | ND | 0.046 (wt) |
| mig-5;syg-2(lh6) | 234 | 814 | 29.5% | 4.2% | 4.1% | <0.001 (additive) |
| syg-2(ky673) | 26 | 1094 | 2.1% | 1.2% | ND | 0.001 (wt) |
| mig-5;syg-2(ky673) | 106 | 391 | 27.1% | 1.9% | 4.9% | <0.001 (additive) |
| syg-1(ky652) | 8 | 735 | 1.0% | 0.8% | ND | |
| mig-5;syg-1 | 129 | 451 | 28.7% | 1.3% | 3.9% | <0.001 (additive) |
| syg-2(lh6);syg-1 | 8 | 406 | 2% | 1% | 2.3% | 1.00 (additive) |
| lin-17(n671) | 63 | 273 | 23.3% | 2.2% | ND | <0.001 (wt) |
| syg-2(lh6);lin-17 | 131 | 260 | 49.8% | 3.9% | 24.3% | <0.001 (additive) |
| syg-1;lin-17 | 297 | 528 | 56.5% | 2.6% | 24.1% | <0.001 (additive) |
| syg-2(lh6)syg-1;lin17 | 302 | 559 | 54.4% | 1.7% | 24.8% | <0.001 (additive) |
| fmi-1(tm306) | 9 | 459 | 1.9% | 0.4% | ND | 0.006 (wt) |
| mig-5;fmi-1 | 153 | 527 | 28.3% | 4.4% | 4.8% | <0.001 (additive) |
| fmi-1;syg-2(lh6) | 33 | 780 | 4.3% | 1.3% | 3.2% | 0.349 (additive) |
| fmi-1;syg-1 | 31 | 607 | 4.9% | 1.3% | 2.9% | 0.079 (additive) |
| fmi-1;mig-5;syg-2(lh6) | 343 | 866 | 40.0% | 3.5% | 29.2% | <0.001 (additive) |
| fmi-1;mig-5;syg-1 | 292 | 570 | 51.4% | 3.7% | 29.0% | <0.001 (additive) |
| dsh-1(ok1445) | 8 | 560 | 1.3% | 0.7% | ND | 0.026 (wt) |
| dsh-1mig-5 | 234 | 671 | 34.9% | 0.0% | 4.2% | <0.001 (additive) |
| dsh-1;lin-17 | 74 | 321 | 24.2% | 6.0% | 24.3% | 0.781 (additive) |
| dsh-1;fmi-1 | 30 | 501 | 6.2% | 1.7% | 3.2% | 0.049 (additive) |
| lin-17;fmi-1 | 256 | 479 | 54.7% | 3.7% | 24.8% | <0.001 (additive) |
| dsh-1;lin-17;fmi-1 | 258 | 450 | 57.9% | 2.0% | 28.9% | <0.001 (additive) |
| Genotype | Pdns | Total | Average | St. Dev. | Additive a | p Value (Comparison) |
|---|---|---|---|---|---|---|
| wild-type | 0 | 416 | 0.0% | 0.0% | ND | |
| mig-5(rh94) | 15 | 219 | 7.0% | 0.7% | ND | <0.0001 (wt) |
| syg-2(lh6) | 11 | 1055 | 1.1% | 1.1% | ND | 0.041 (wt) |
| mig-5;syg-2(lh6) | 148 | 516 | 29.2% | 3.9% | 8.0% | <0.0001 (additive) |
| syg-2(ky673) | 19 | 1294 | 1.4% | 0.2% | ND | 0.007 (wt) |
| mig-5;syg-2(ky673) | 140 | 409 | 34.2% | 2.0% | 8.3% | <0.0001 (additive) |
| syg-1(ky652) | 17 | 1136 | 1.3% | 0.7% | ND | |
| mig-5;syg-1 | 228 | 702 | 32.1% | 3.1% | 8.2% | <0.0001 (additive) |
| syg-2(lh6);syg-1 | 8 | 464 | 1.9% | 1.3% | 2.4% | 0.644 (additive) |
| fmi-1(tm306) | 0 | 585 | 0.0% | 0.0% | ND | 1.000 (wt) |
| mig-5;fmi-1 | 51 | 754 | 7.3% | 2.9% | 7.0% | 0.919 (additive) |
| fmi-1;syg-2(lh6) | 13 | 729 | 1.9% | 0.6% | 1.1% | 0.380 (additive) |
| fmi-1;syg-1 | 12 | 563 | 2.2% | 0.7% | 1.3% | |
| fmi-1;mig-5;syg-2(lh6) | 189 | 597 | 31.9% | 3.7% | 8.3% | <0.0001 (additive) |
| fmi-1;mig-5;syg-1(ky652) | 264 | 804 | 33.2% | 4.7% | 8.5% | <0.0001 (additive) |
| lin-17(n671) | 54 | 1111 | 2.7% | 3.2% | ND | <0.0001 (wt) |
| dsh-1(ok1445) | 0 | 188 | 0% | 0.0% | ND | 1.000 (wt) |
| dsh-1(ok1445)mig-5(tm2639) | 42 | 157 | 27% | 0.0% | 7.0% | <0.0001 (additive) |
| syg-2(lh6);lin-17 | 103 | 207 | 49.1% | 1.7% | 2.4% | <0.0001 (additive) |
| syg-1;lin-17 | 172 | 325 | 52.6% | 2.7% | 2.6% | <0.0001 (additive) |
| syg-2(lh6)syg-1;lin17 | 383 | 775 | 49.5% | 2.8% | 3.2% | <0.0001 (additive) |
| lin-17;fmi-1 | 13 | 401 | 2.6% | 1.3% | 2.7% | 0.997 (additive) |
| Genotype | Pdns | Total | Unipolar | Bipolar | Ambiguous | Bipolar (%) |
|---|---|---|---|---|---|---|
| L1 | ||||||
| syg-2(lh6) | 7 | 454 | 0 | 7 | 0 | 100% |
| syg-2(lh6);mig-5(rh94) | 110 | 766 | 0 | 110 | 0 | 100% |
| lin-17(n671) | 47 | 567 | 0 | 47 | 0 | 100% |
| lin-17(n671);syg-2(lh6) | 318 | 913 | 4 | 314 | 0 | 99% |
| dsh-1(ok1445)mig-5(tm2639) | 42 | 157 | 0 | 42 | 0 | 100% |
| Adult | ||||||
| syg-2(lh6) | 3 | 373 | 0 | 2 | 1 | 67% |
| syg-2(lh6);mig-5(rh94) | 139 | 558 | 31 | 69 | 39 | 50% |
| lin-17(n671) | 60 | 417 | 31 | 29 | 0 | 48% |
| lin-17(n671);syg-2(lh6) | 104 | 295 | 13 | 40 | 51 | 38% |
| dsh-1(ok1445)mig-5(tm2639) | 234 | 671 | 0 | 234 | 0 | 100% |
| Genotype | Area (Avg ± SD) μm2 | N | p (vs. Wild Type) | Puncta (Avg ± SD) #/100 μm | N | p (vs. Wild Type) |
|---|---|---|---|---|---|---|
| Ventral Cord (VD Neurons) | ||||||
| wild type | 1.12 ± 1.17 | 517 | - | 24.0 ± 2.9 | 11 | - |
| syg-2(lh6) | 1.53 ± 2.11 | 402 | 4 × 10−4 | 21.9 ± 2.3 | 9 | 0.09 |
| Dorsal Cord (DD Neurons) | ||||||
| wild type | 0.68 ± 0.41 | 326 | 21.9 ± 4.7 | 9 | ||
| syg-2(lh6) | 1.01 ± 1.16 | 214 | 1 × 10−4 | 14.1 ± 3.9 | 8 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tucker, D.K.; Adams, C.S.; Prasad, G.; Ackley, B.D. The Immunoglobulin Superfamily Members syg-2 and syg-1 Regulate Neurite Development in C. elegans. J. Dev. Biol. 2022, 10, 3. https://doi.org/10.3390/jdb10010003
Tucker DK, Adams CS, Prasad G, Ackley BD. The Immunoglobulin Superfamily Members syg-2 and syg-1 Regulate Neurite Development in C. elegans. Journal of Developmental Biology. 2022; 10(1):3. https://doi.org/10.3390/jdb10010003
Chicago/Turabian StyleTucker, Dana K., Chloe S. Adams, Gauri Prasad, and Brian D. Ackley. 2022. "The Immunoglobulin Superfamily Members syg-2 and syg-1 Regulate Neurite Development in C. elegans" Journal of Developmental Biology 10, no. 1: 3. https://doi.org/10.3390/jdb10010003
APA StyleTucker, D. K., Adams, C. S., Prasad, G., & Ackley, B. D. (2022). The Immunoglobulin Superfamily Members syg-2 and syg-1 Regulate Neurite Development in C. elegans. Journal of Developmental Biology, 10(1), 3. https://doi.org/10.3390/jdb10010003