
Gait Transition from Pacing by a Quadrupedal Simulated Model and Robot with Phase Modulation by Vestibular Feedback

Abstract
:1. Introduction
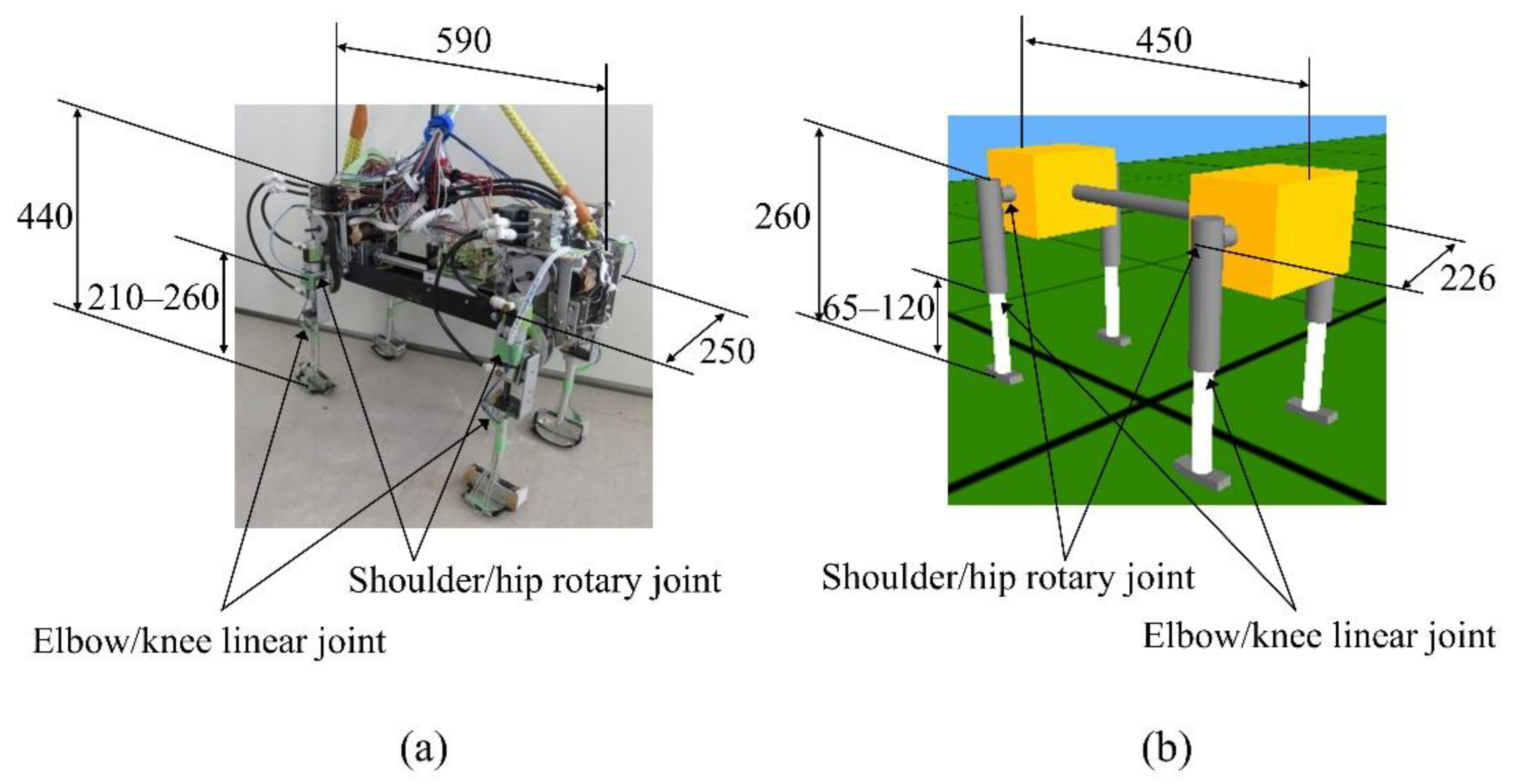
2. Spinalbot Quadruped Robot and Simulated Model
3. Motion Generation Control
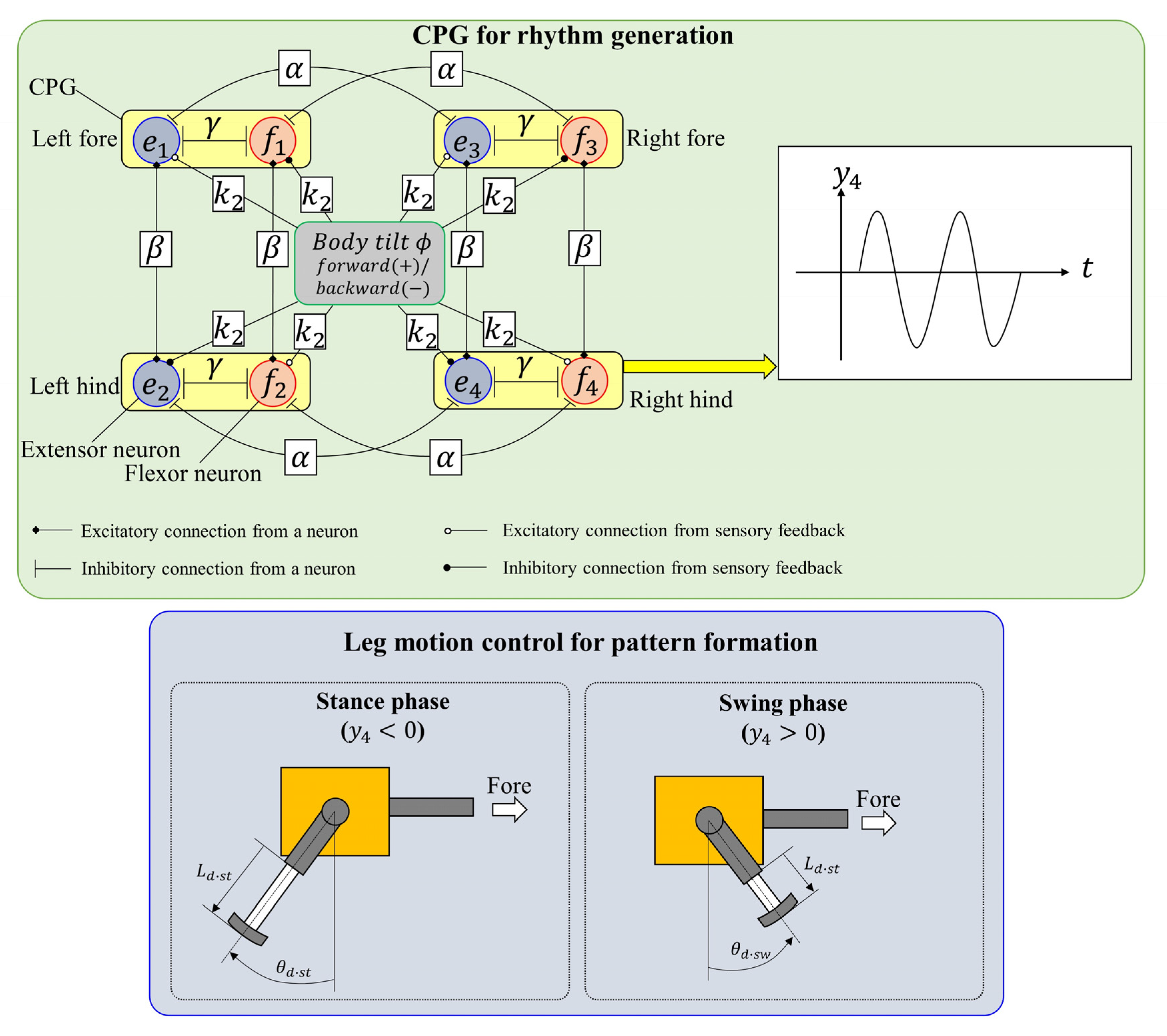
3.1. Rhythm Generation
3.1.1. CPG Model and CPG Network
3.1.2. Vestibular Modulation
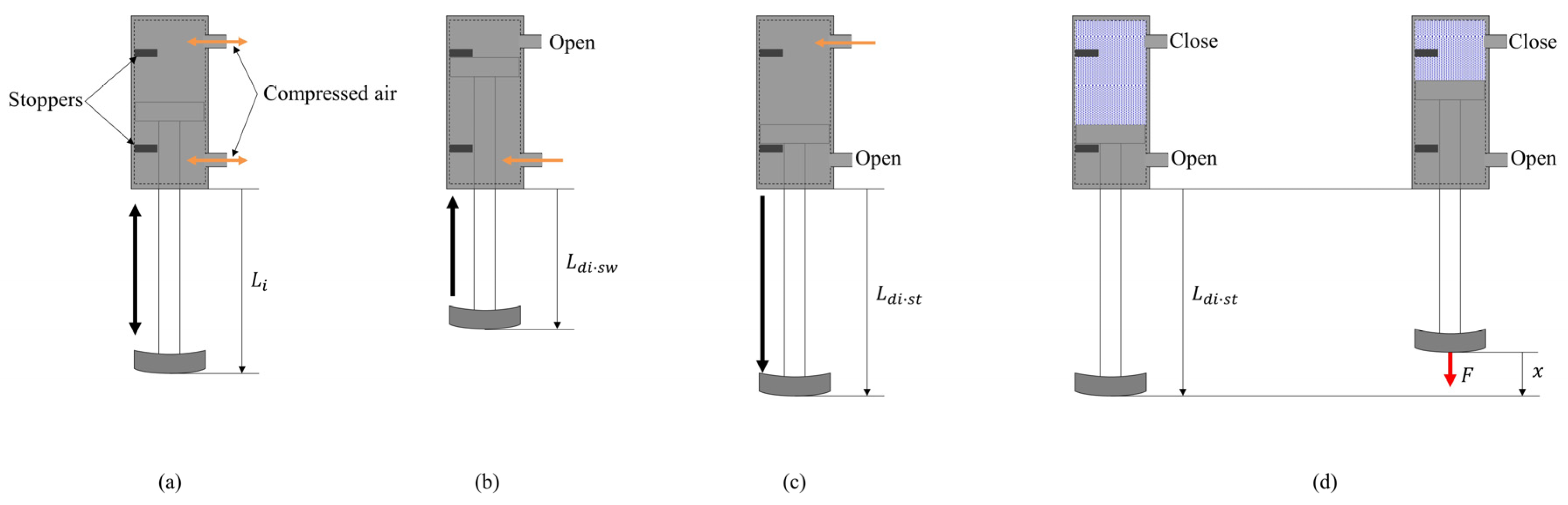
3.2. Leg Motion Generation
4. Results
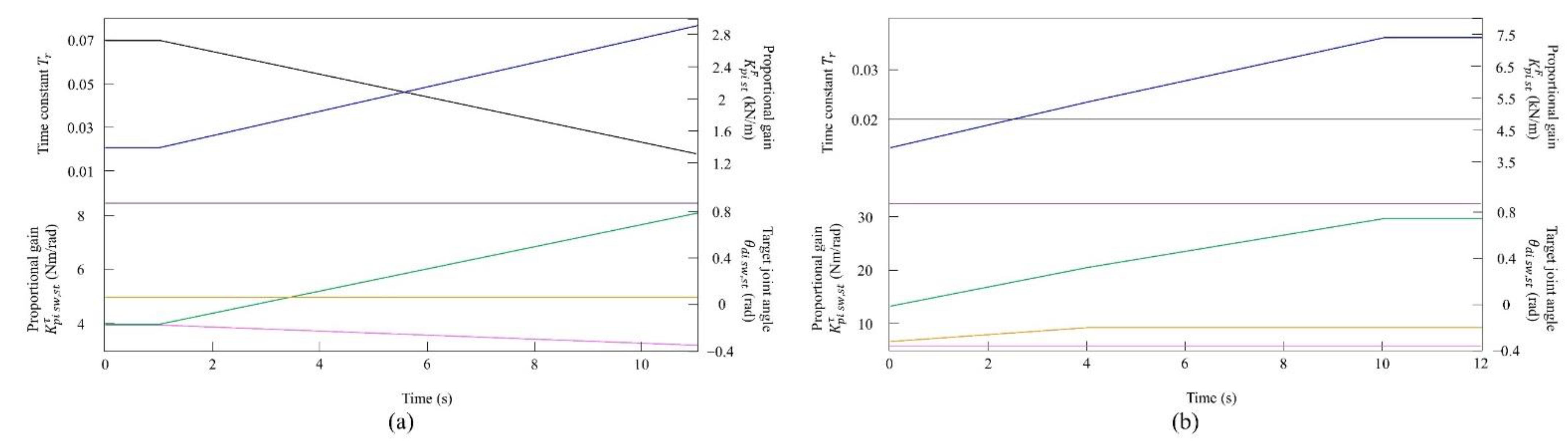
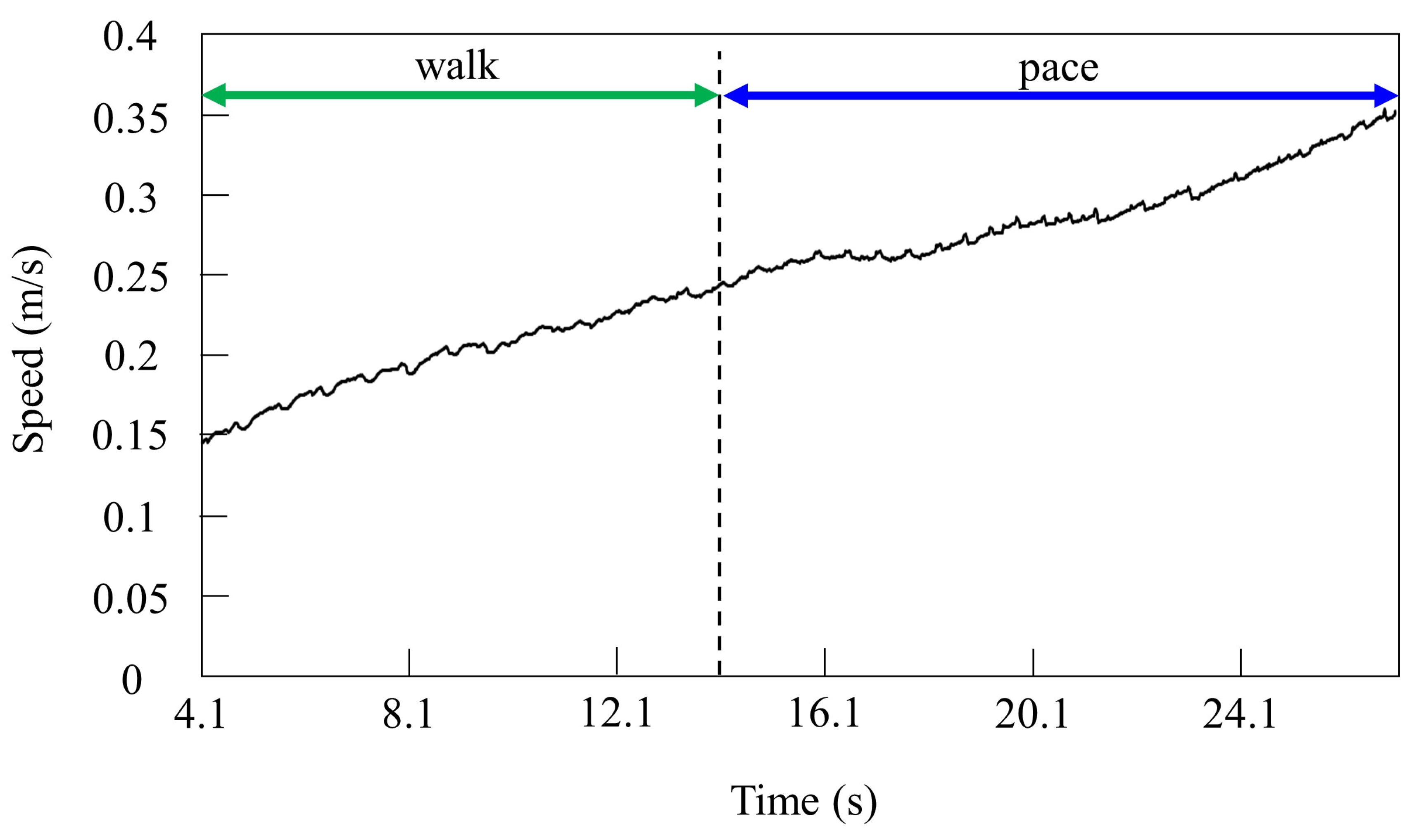
4.1. Speed Parameter
4.2. Simulated Results
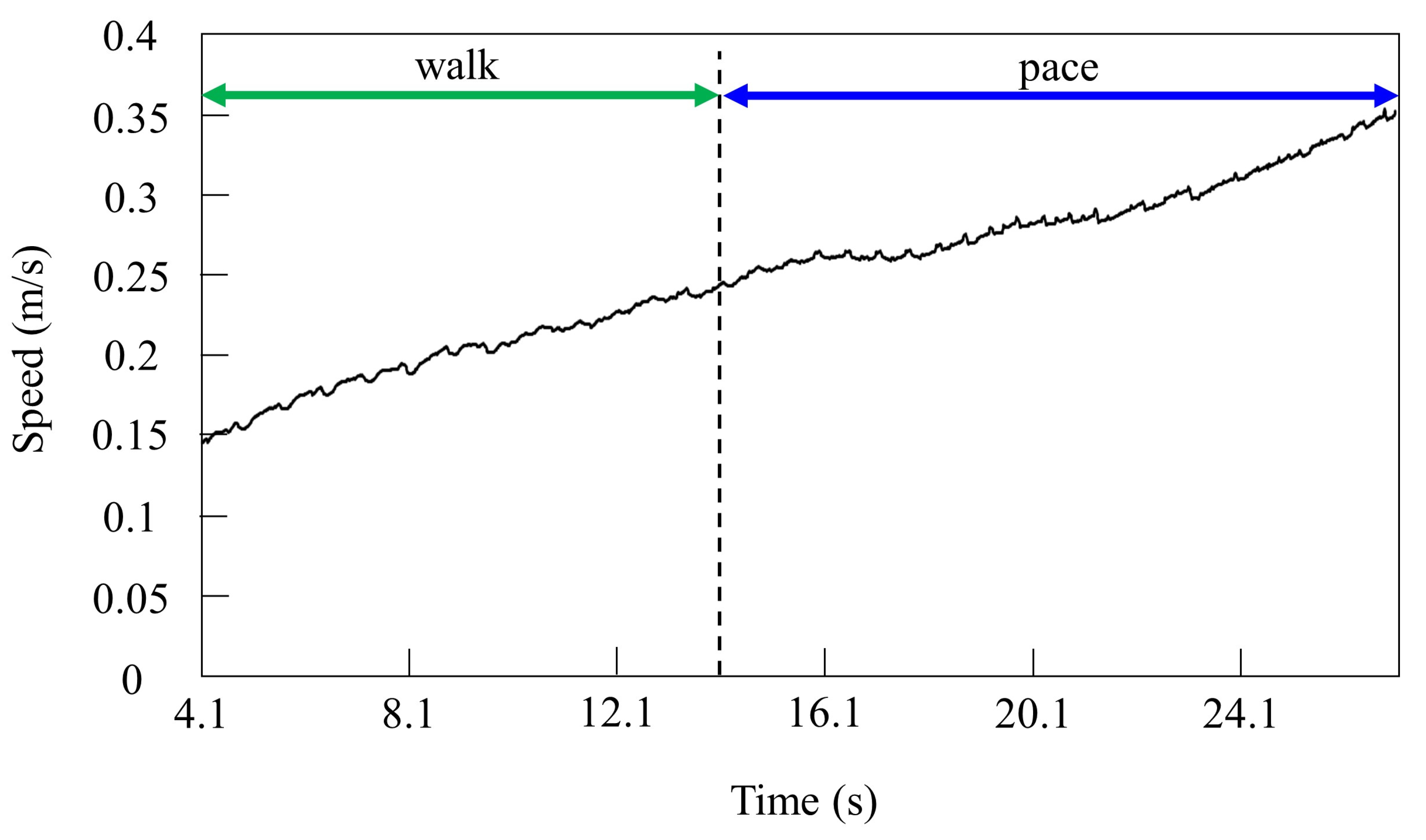
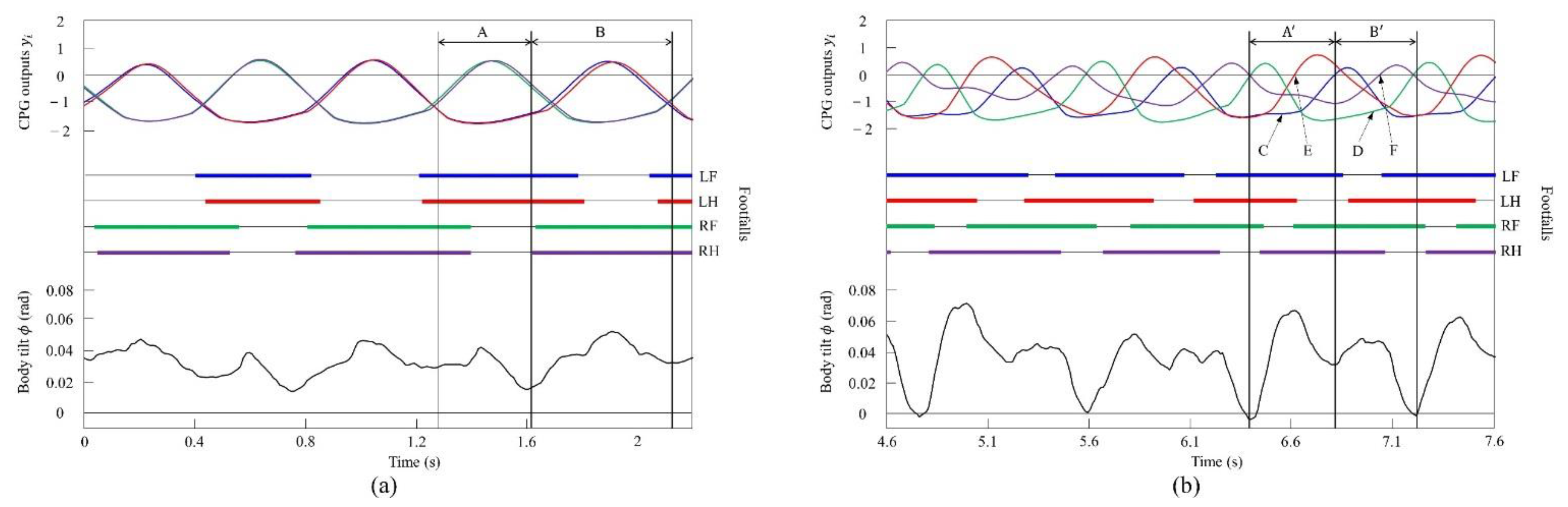
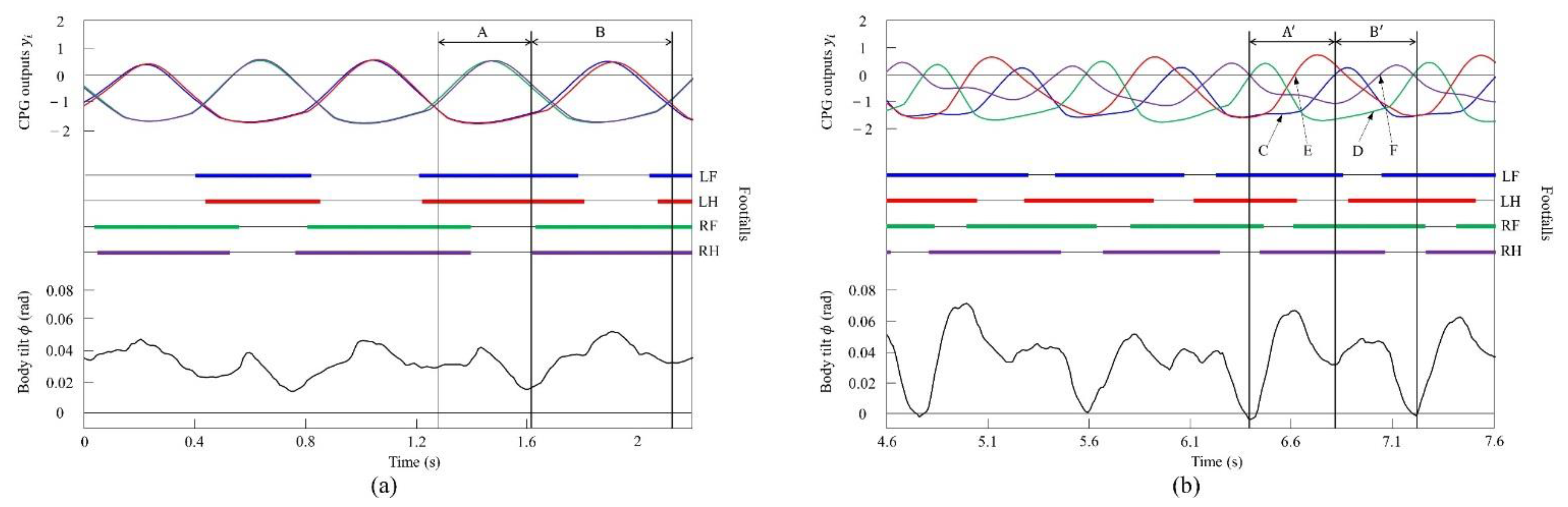
4.2.1. Gait Transition from Walking to Pacing with Vestibular Modulation in Simulation
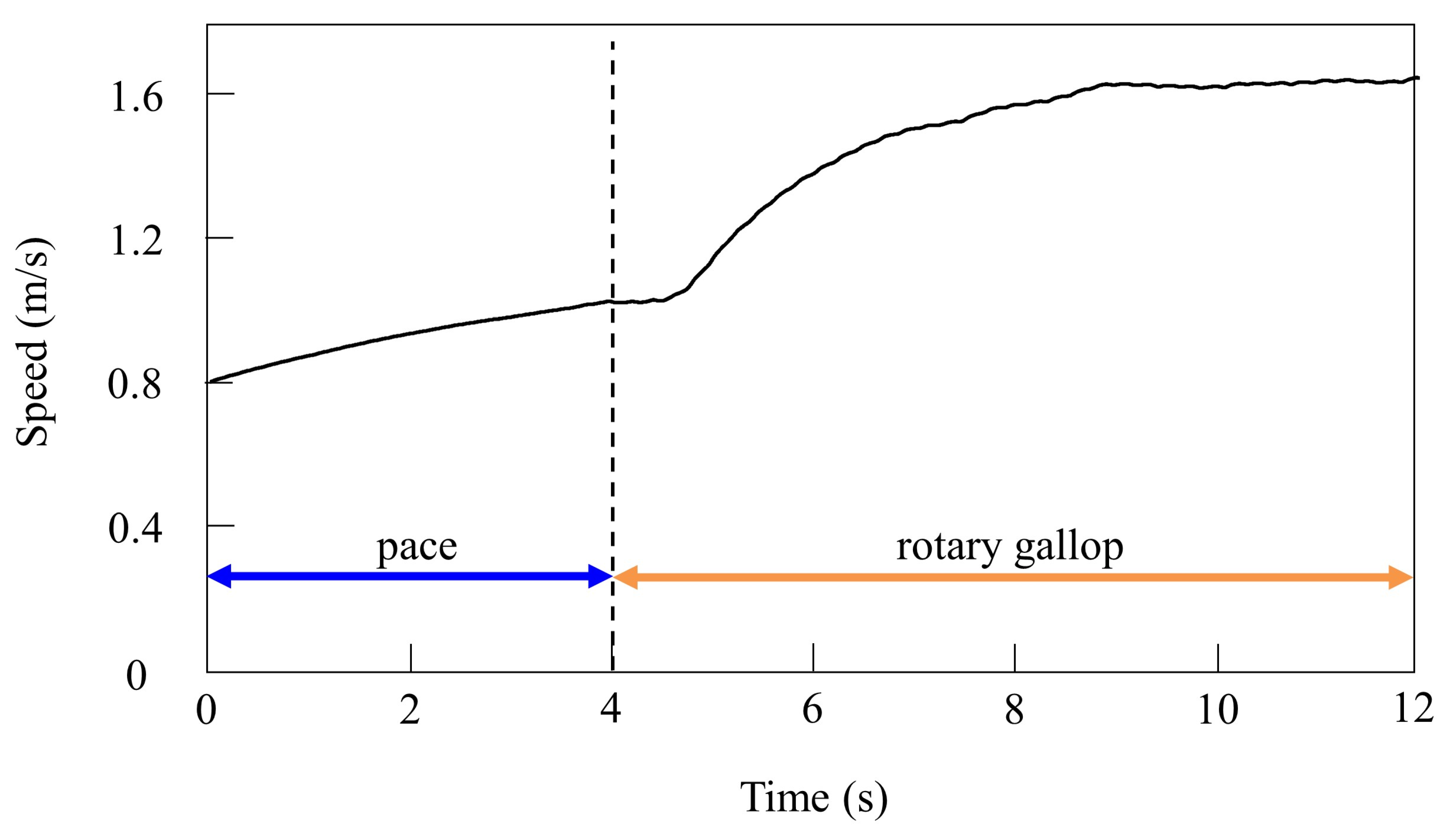
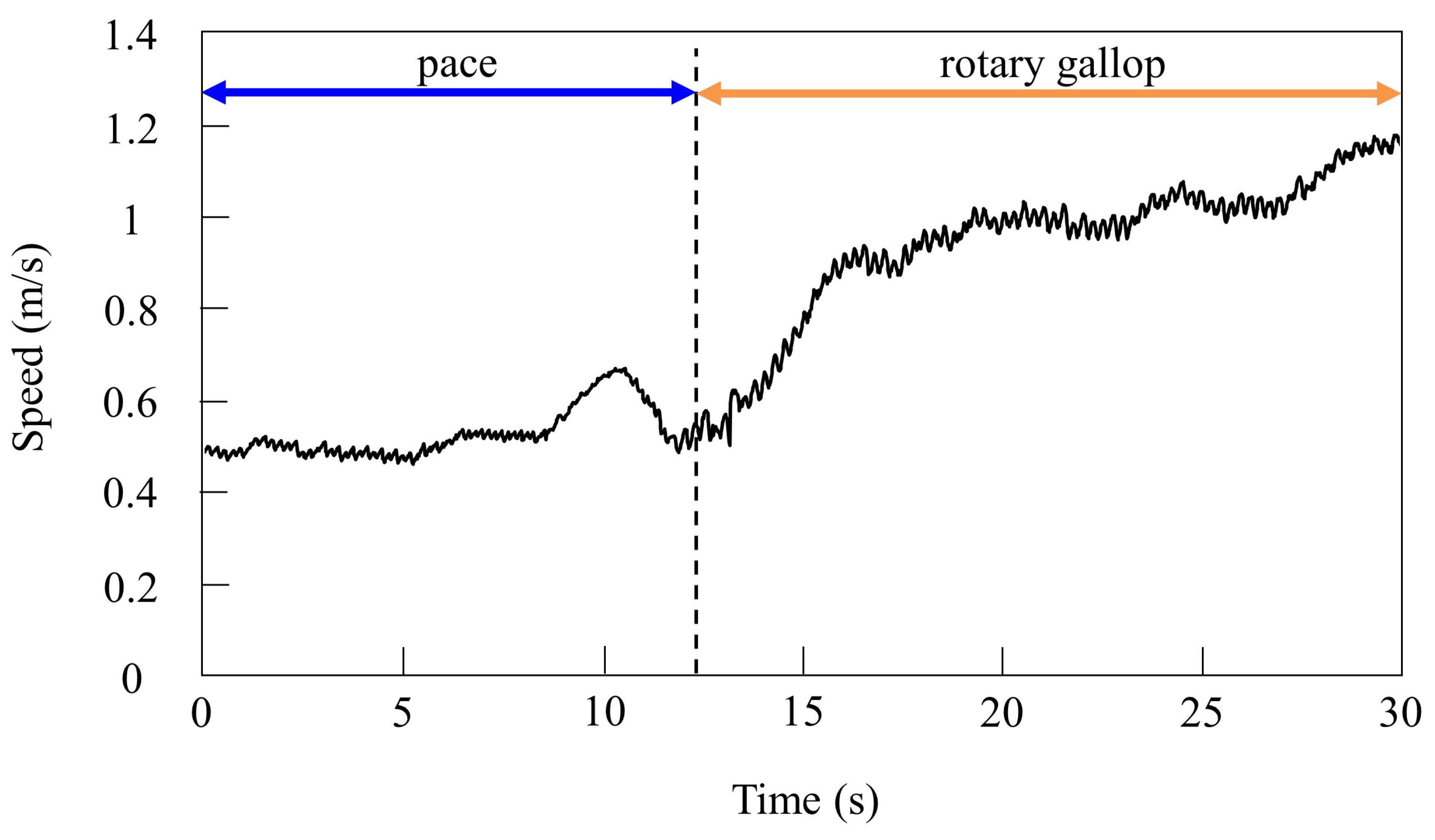
4.2.2. Gait Transition from Pacing to Galloping with Vestibular Modulation in Simulation
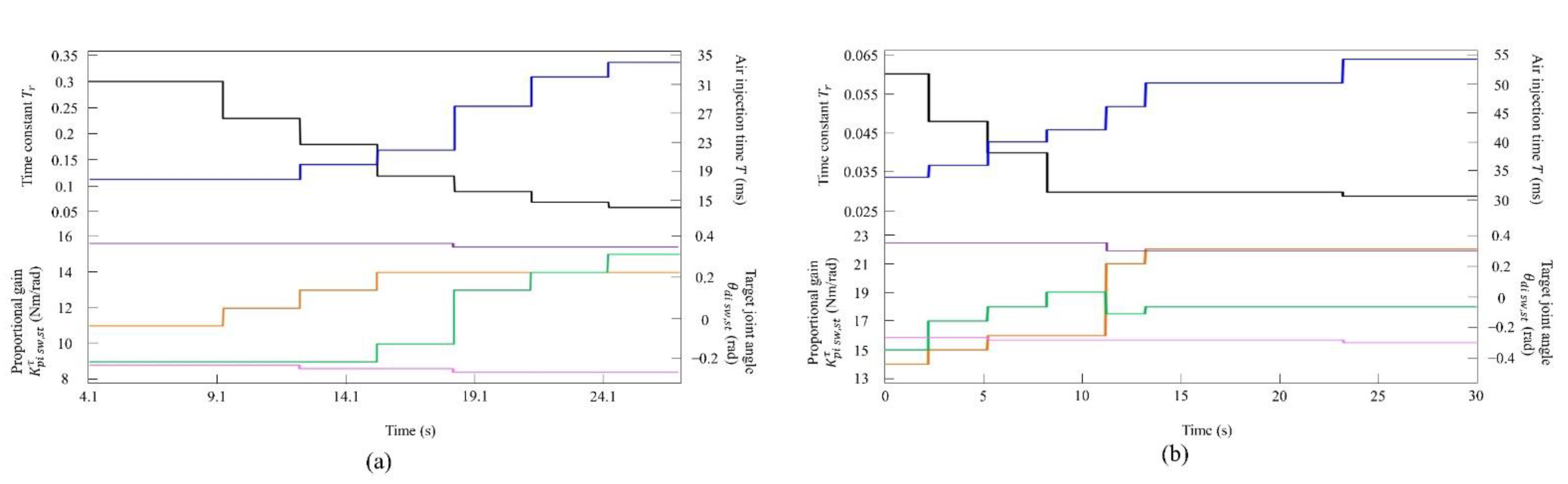
4.3. Experimental Results
4.3.1. Gait Transition from Walking to Pacing with Vestibular Modulation
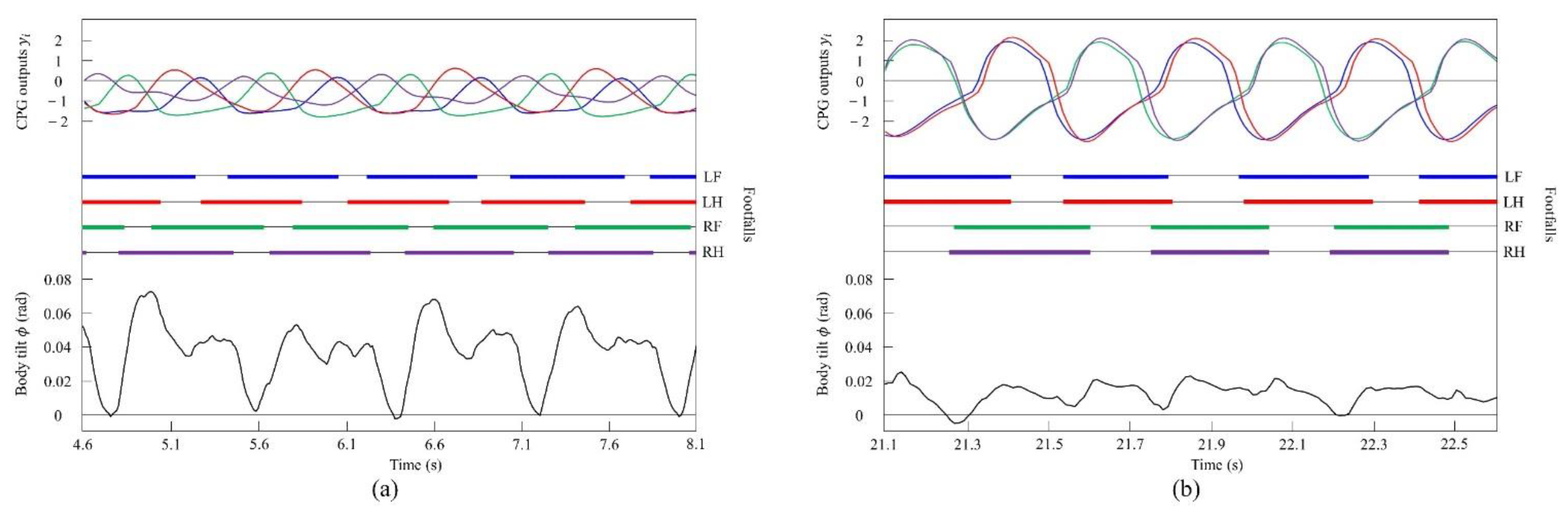
4.3.2. Gait Transition from Pacing to Galloping with Vestibular Modulation
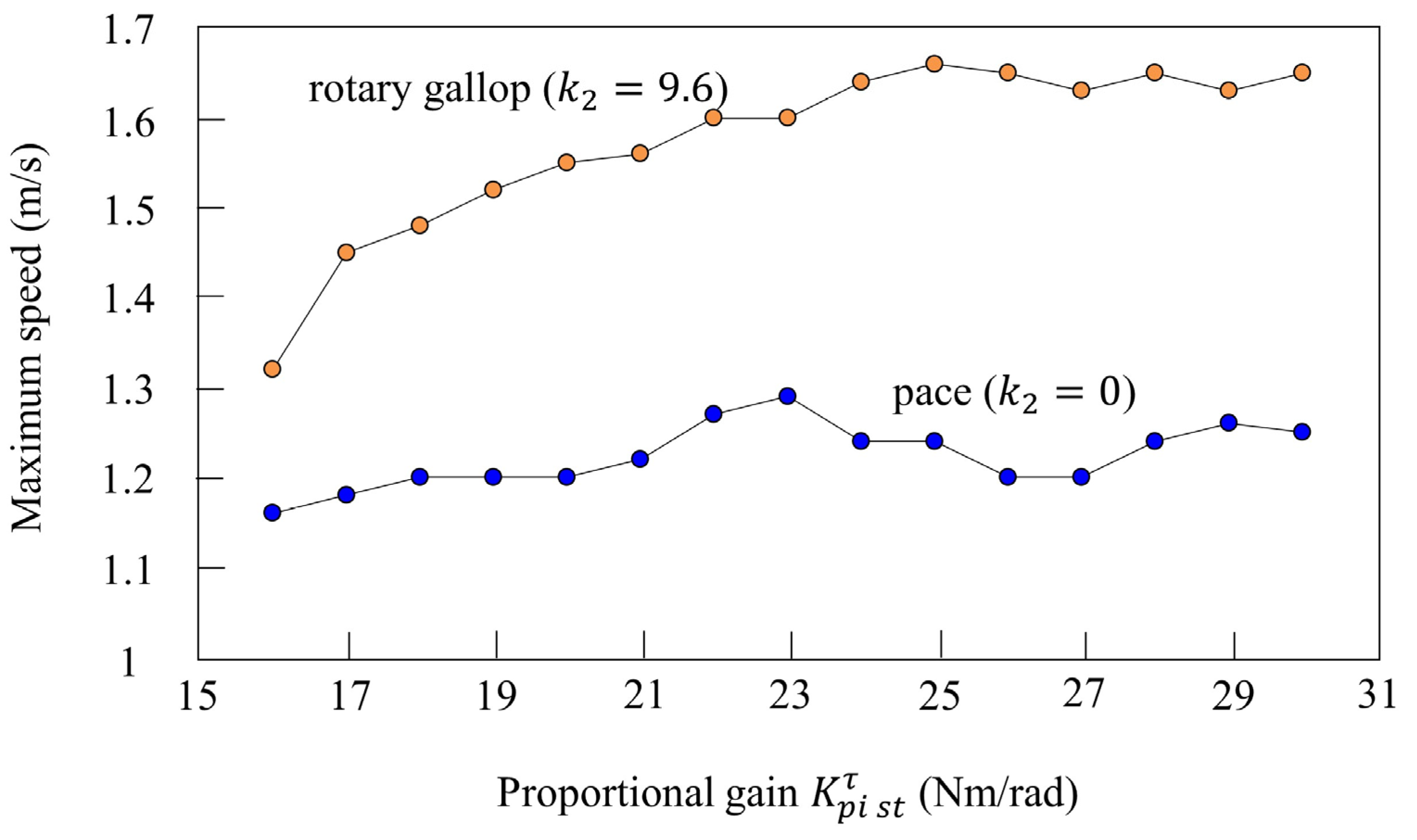
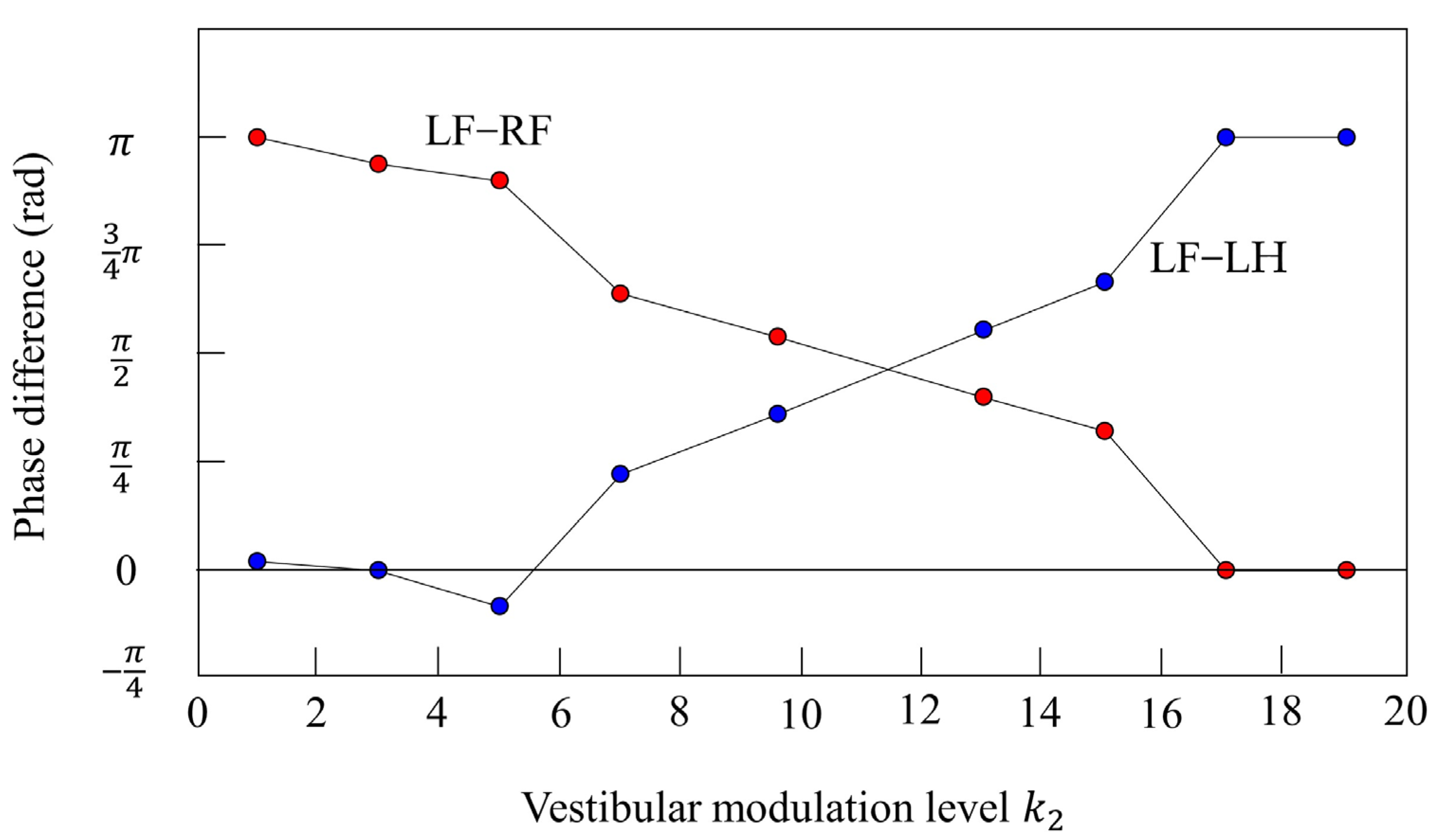
4.4. Vestibular Modulation Level
5. Discussion
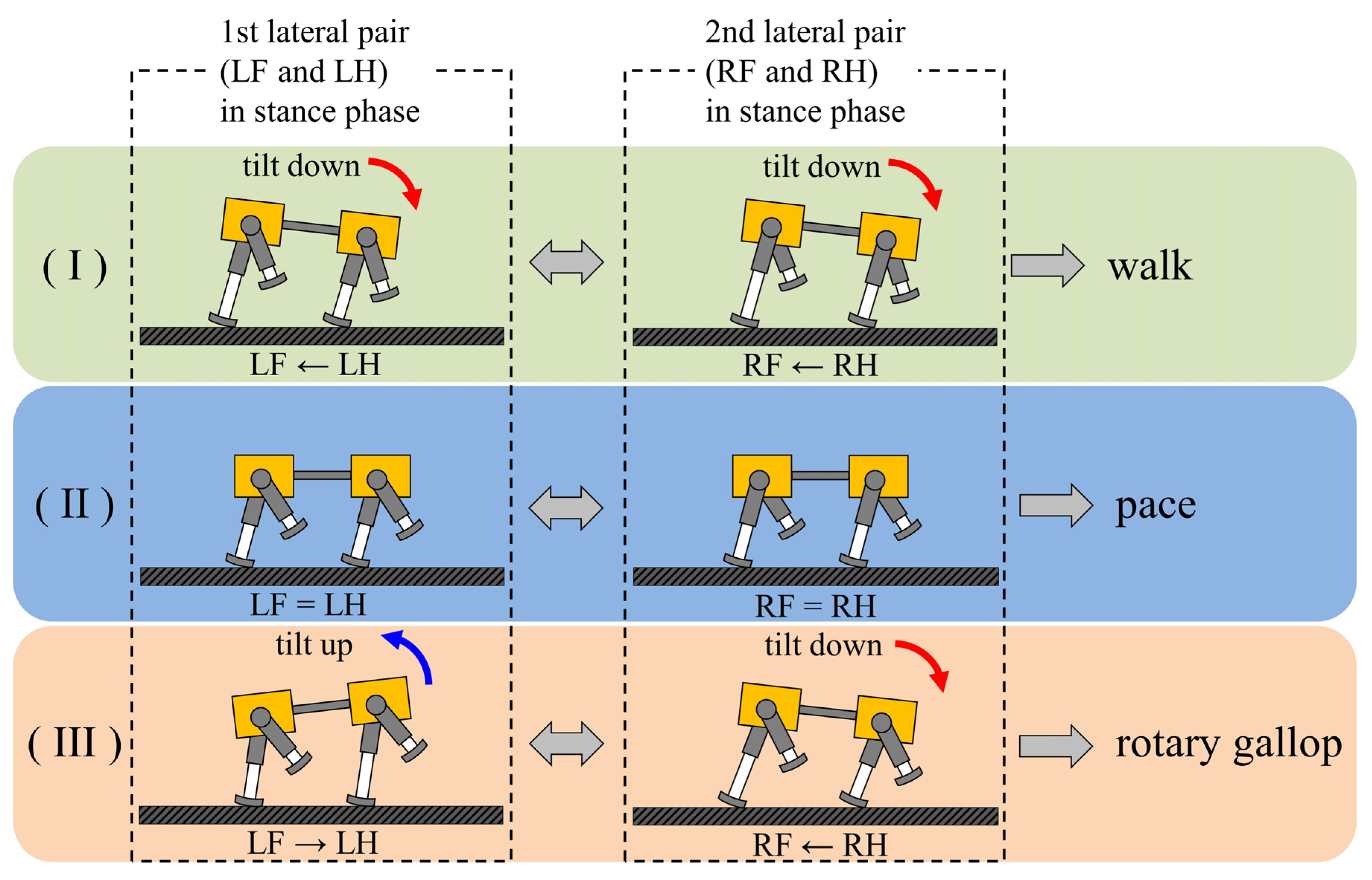
5.1. Principle Emergence of Walking
5.2. Principle Emergence of Galloping
5.3. Difference between Factors behind Emergence of Walking and Galloping
5.4. Transition Speed
5.5. Related Works
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muybridge, E. Animal in Motion; Dover Publications: New York, NY, USA, 1957. [Google Scholar]
- Hildebrand, M. The adaptive significance of tetrapod gait selection. Am. Zool. 1980, 20, 255–267. [Google Scholar] [CrossRef]
- Hudson, P.E.; Corr, S.A.; Wilson, A.M. High speed galloping in the cheetah (Acinonyx jubatus) and the racing greyhound (Cains familiaris): Spatio-temporal and kinetic characteristics. J. Exp. Biol. 2012, 215, 2425–2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmiedeler, J.P.; Waldron, K.J. The mechanics of quadrupedal galloping and the future of legged vehicles. Int. J. Robot. Res. 1999, 18, 1224–1234. [Google Scholar] [CrossRef]
- Hawker, G.; Buehler, M. Quadruped trotting with passive knees: Design, control, and experiments. In Proceedings of the IEEE International Conference on Robotics and Automation (ICRA), San Francisco, CA, USA, 24–28 April 2000; pp. 24–28. [Google Scholar]
- Raibert, M.; Blankespoor, K.; Nelson, G.; Playter, R. BigDog, the rough-terrain quadruped robot. In Proceedings of the 17th World Congress. The International Federation of Automatic Control (IFAC), Seoul, Korea, 6–11 July 2008; Volume 41, pp. 10822–10825. [Google Scholar]
- Nakatani, K.; Sugimoto, Y.; Osuka, K. Demonstration and analysis of quadrupedal passive dynamic walking. Adv. Robot. 2009, 23, 483–501. [Google Scholar] [CrossRef]
- Santos, C.P.; Matos, V. Gait transition and modulation in a quadruped robot: A brainstem-like modulation approach. Robot. Auton. Syst. 2011, 59, 620–634. [Google Scholar] [CrossRef] [Green Version]
- Moro, F.L.; Spröwitz, A.; Tuleu, A.; Vespignani, M.; Tsagarakis, N.G.; Ijspeert, A.J.; Caldwell, D.G. Horse-like walking, trotting and galloping derived from kinematic motion primitives(kMPs) and their application to walk/trot transitions in a compliant quadruped robot. Biol. Cybern. 2013, 107, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Gehring, C.; Coros, S.; Hutter, M.; Bloesch, M.; Hoepflinger, M.A.; Siegwart, R. Control of dynamic gaits for a quadrupedal robot. In Proceedings of the IEEE International Conference on Robotics and Automation (ICRA), Karlsruhe, Germany, 6–10 May 2013. [Google Scholar] [CrossRef] [Green Version]
- Spröwitz, A.; Tuleu, A.; Vespignani, M.; Ajallooeian, M.; Badri, E.; Ijspeert, A.J. Towards dynamic trot gait locomotion: Design, control, and experiments with Cheetah-cub, a compliant quadruped robot. Int. J. Robot. Res. 2013, 32, 932–950. [Google Scholar] [CrossRef] [Green Version]
- Hutter, M.; Gehring, C.; Höpflinger, M.A.; Blösch, M.; Siegwart, R. Toward combining speed, efficiency, versatility, and robustness in an autonomous quadruped. IEEE Trans. Robot. 2014, 30, 1427–1440. [Google Scholar] [CrossRef]
- Li, M.; Jiang, Z.; Wang, P.; Sun, L.; Ge, S.S. Control of a quadruped robot with bionic springy legs in trotting gait. J. Bionic Eng. 2014, 11, 188–198. [Google Scholar] [CrossRef]
- Koo, I.M.; Trong, T.D.; Lee, Y.H.; Moon, H.; Koo, J.; Park, S.; Choi, H.R. Biologically inspired gait transition control for a quadruped walking robot. Auton. Robot. 2015, 39, 169–182. [Google Scholar] [CrossRef]
- Semini, C.; Barasuol, V.; Boaventura, T.; Frigerio, M.; Focchi, M.; Caldwell, D.G.; Buchli, J. Towards versatile legged robots through active impedance control. Int. J. Robot. Res. 2015, 34, 1003–1020. [Google Scholar] [CrossRef]
- Hyun, D.J.; Lee, J.; Park, S.; Kim, S. Implementation of trot-to-gallop transition and subsequent gallop on the MIT Cheetah I. Int. J. Robot. Res. 2016, 35, 1627–1650. [Google Scholar] [CrossRef]
- Kitano, S.; Hirose, S.; Horigome, A.; Endo, G. TITAN-XIII: Sprawling-type quadruped robot with ability of fast and energy-efficient walking. ROBOMECH J. 2016, 3. [Google Scholar] [CrossRef]
- Hutter, M.; Gehring, C.; Lauber, A.; Gunther, F.; Bellicoso, C.D.; Tsounis, V.; Fankhauser, P.; Diethelm, R.; Bachmann, S.; Bloesch, M.; et al. ANYmal—Toward legged robots for harsh environments. Adv. Robot. 2017, 31, 918–931. [Google Scholar] [CrossRef]
- Boston Dynamics, Spot. Available online: http://www.bostondynamics.com/ (accessed on 9 December 2021).
- Grillner, S. Handbook of Physiology; John Wiley & Sons: Hoboken, NJ, USA, 1981; pp. 1179–1236. [Google Scholar]
- Golubitsky, M.; Stewart, I.; Buono, P.L.; Collins, J.J. Symmetry in locomotor central pattern generators and animal gaits. Nature 1999, 401, 693–695. [Google Scholar] [CrossRef]
- Ito, S.; Yuasa, H.; Luo, Z.W.; Ito, M.; Yanagihara, D. A mathematical model of adaptive behavior in quadruped locomotion. Biol. Cybern. 1998, 78, 337–347. [Google Scholar] [CrossRef]
- Tsujita, K.; Tsuchiya, K.; Onat, A. Adaptive gait pattern control of a quadruped locomotion robot. In Proceedings of the IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Maui, HI, USA, 29 October–3 November 2001. [Google Scholar] [CrossRef]
- Tsujita, K.; Kobayashi, T.; Inoura, T.; Masuda, T. Gait transition by tuning muscle tones using pneumatic actuators in quadruped locomotion. In Proceedings of the IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Nice, France, 22–26 September 2008. [Google Scholar] [CrossRef]
- Harischandra, N.; Knuesel, J.; Kozlov, A.; Bicanski, A.; Cabelguen, J.M.; Ijspeert, A.; Ekeberg, Ö. Sensory feedback plays a significant role in generating walking gait and in gait transition in salamanders: A simulation study. Front. Neurorobotics 2011, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Aoi, S.; Yamashita, T.; Tsuchiya, K. Hysteresis in the gait transition of a quadruped investigated using simple body mechanical and oscillator network models. Phys. Rev. E 2011, 83, 061909. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H.; Akiyama, S.; Sakurama, K. Realization of dynamic walking and running of the quadruped using neural oscillator. Auton. Robot. 1999, 7, 247–258. [Google Scholar] [CrossRef]
- Fukuoka, Y.; Kimura, H.; Cohen, A.H. Adaptive dynamic walking of a quadruped robot on irregular terrain based on biological concepts. Int. J. Robot. Res. 2003, 22, 187–202. [Google Scholar] [CrossRef]
- Fukuoka, Y.; Kimura, H. Dynamic locomotion of a biomorphic quadruped‘Tekken ’robot using various gaits: Walk, Trot, Free-Gait and Bound. Appl. Bionics Biomech. 2009, 6, 63–71. [Google Scholar] [CrossRef]
- Aoi, S.; Katayama, D.; Fujiki, S.; Tomita, N.; Funato, T.; Yamashita, T.; Senda, K.; Tsuchiya, K. A stability-based mechanism for hysteresis in the walk-trot transition in quadruped locomotion. J. R. Soc. Interface 2013, 10, 20120908. [Google Scholar] [CrossRef] [Green Version]
- Fukuoka, Y.; Habu, Y.; Fukui, T. Analysis of the gait generation principle by a simulated quadruped model with a CPG incorporating vestibular modulation. Biol. Cybern. 2013, 107, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Fukui, T.; Fujisawa, H.; Otaka, K.; Fukuoka, Y. Autonomous gait transition and galloping over unperceived obstacles of a quadruped robot with CPG modulated by vestibular feedback. Robot. Auton. Syst. 2019, 111, 1–19. [Google Scholar] [CrossRef]
- Fukuoka, Y.; Habu, Y.; Fukui, T. A simple rule for quadrupedal gait generation determined by leg loading feedback: A modeling study. Sci. Rep. 2015, 5, 8169. [Google Scholar] [CrossRef] [Green Version]
- Owaki, D.; Ishiguro, A. A quadruped robot exhibiting spontaneous gait transitions from walking to trotting to galloping. Sci. Rep. 2017, 7, 277. [Google Scholar] [CrossRef]
- Kimura, H.; Shimoyama, I.; Miura, H. Dynamics in the dynamic walk of a quadruped robot. Adv. Robot. 1989, 4, 283–301. [Google Scholar] [CrossRef] [Green Version]
- Raibert, M.H. Trotting, pacing and bounding by a quadruped robot. J. Biomech. 1990, 23, 79–81, 83–98. [Google Scholar] [CrossRef]
- Sano, A.; Furusho, J. Realization of dynamic quadruped locomotion in pace gait by controlling walking cycle. In Experimental Robotics II; Springer: Berlin/Heidelberg, Germany, 2005; pp. 491–502. [Google Scholar]
- Homby, G.S.; Takamura, S.; Yamamoto, T.; Fujita, M. Autonomous evolution of dynamic gaits with two quadruped robots. IEEE Trans. Robot. 2005, 21, 402–410. [Google Scholar]
- Smith, J.A.; Poulakakis, I. Rotary gallop in the untethered quadrupedal robot scout II. In Proceedings of the IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Sendai, Japan, 28 September–2 October 2004. [Google Scholar] [CrossRef]
- Geyer, H.; Seyfarth, A.; Blickhan, R. Compliant leg behavior explains basic dynamics of walking and running. Proc. R. Soc. B 2006, 273, 861–2867. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, K. Mechanisms of frequency and pattern control in the neural rhythm generators. Biol. Cybern. 1987, 56, 345–353. [Google Scholar] [CrossRef]
- Andersson, O.; Grillner, S. Peripheral control of the cat’s step cycle: II. Entrainment of the central pattern generators for locomotion by sinusoidal hip movements during “fictive locomotion”. Acta Physiol. Scand. Banner 1983, 118, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Duysens, J.; Clarac, F.; Cruse, H. Load-regulating mechanisms in gait and posture: Comparative aspects. Physiol. Rev. 2000, 80, 83–133. [Google Scholar] [CrossRef] [Green Version]
- Roberts, T.D. Neurophysiology of Postural Mechanisms; Butterworths: Oxford, UK, 1967. [Google Scholar]
- Habu, Y.; Uta, K.; Fukuoka, Y. Three-dimensional walking of a simulated muscle-driven quadruped robot with neuromorphic two-level central pattern generators. Int. J. Adv. Robot. Syst. 2019, 16. [Google Scholar] [CrossRef]
- Fukuoka, Y.; Fukino, K.; Habu, Y.; Mori, Y. Energy evaluation of a bio-inspired gait modulation method for quadrupedal locomotion. Bioinspiration Biomim. 2015, 10, 046017. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Owaki, D.; Fukuhara, A.; Ishiguro, A. Quadruped gait transition from walk to pace to rotary gallop by exploiting head movement. In International Conference on Biomimetic and Biohybrid Systems; Springer: Cham, Switzerland, 2016; pp. 532–539. [Google Scholar]
- Suzuki, S.; Kano, T.; Ispeert, A.J.; Ishiguro, A. Sprawling quadruped robot driven by decentralized control with cross-coupled sensory feedback between legs and trunk. Front. Neurorobotics 2021, 14, 116. [Google Scholar] [CrossRef]
- Dürr, V.; Arena, P.P.; Cruse, H.; Dallmann, C.J.; Drimus, A.; Hoinville, T.; Krause, T.; Mátéfi-Tempfli, S.; Paskarbeit, J.; Patanè, L.; et al. Integrative biomimetics of autonomous hexapedal locomotion. Front. Neurorobotics 2019, 13, 88. [Google Scholar] [CrossRef]
- Manoonpong, P.; Patanè, L.; Xiong, X.; Brodoline, I.; Dupeyroux, J.; Viollet, S.; Arena, P.; Serres, J.R. Insect-inspired robots: Bridging biological and artificial systems. Sensors 2021, 21, 7609. [Google Scholar] [CrossRef] [PubMed]
- Calandra, M.; Patanè, L.; Sun, T.; Arena, P.; Manoonpong, P. Echo state networks for estimating exteroceptive conditions from proprioceptive states in quadruped robots. Front. Neurorobotics 2021, 15, 118. [Google Scholar] [CrossRef]
- Hwangbo, J.; Lee, J.; Dosovitskiy, A.; Bellicoso, D.; Tsounis, V.; Koltun, V.; Hutter, M. Learning agile and dynamic motor skills for legged robots. Sci. Robot. 2019, 4, 26. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value (Simulation) | Value (Robot Experiment) | Parameter | Value (Simulation) | Value (Robot Experiment) |
|---|---|---|---|---|---|
| 0.6 | 0.16 | −0.26 rad | −1.05 rad | ||
| −0.54 | −0.69 | 1.0 Nms/rad | 0.24 Nms/rad | ||
| 0.1 | 0.29 | 1.0 Nms/rad | 0.34 Nms/rad | ||
| −2.0 | −2.0 | 65 mm | 175 mm | ||
| 3.4 | 5.5 | 120 mm | 225 mm | ||
| 3.0 | 4.0 | 3.0 | 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukui, T.; Matsukawa, S.; Habu, Y.; Fukuoka, Y. Gait Transition from Pacing by a Quadrupedal Simulated Model and Robot with Phase Modulation by Vestibular Feedback. Robotics 2022, 11, 3. https://doi.org/10.3390/robotics11010003
Fukui T, Matsukawa S, Habu Y, Fukuoka Y. Gait Transition from Pacing by a Quadrupedal Simulated Model and Robot with Phase Modulation by Vestibular Feedback. Robotics. 2022; 11(1):3. https://doi.org/10.3390/robotics11010003
Chicago/Turabian StyleFukui, Takahiro, Souichiro Matsukawa, Yasushi Habu, and Yasuhiro Fukuoka. 2022. "Gait Transition from Pacing by a Quadrupedal Simulated Model and Robot with Phase Modulation by Vestibular Feedback" Robotics 11, no. 1: 3. https://doi.org/10.3390/robotics11010003
APA StyleFukui, T., Matsukawa, S., Habu, Y., & Fukuoka, Y. (2022). Gait Transition from Pacing by a Quadrupedal Simulated Model and Robot with Phase Modulation by Vestibular Feedback. Robotics, 11(1), 3. https://doi.org/10.3390/robotics11010003