Antioxidative Properties of Melanins and Ommochromes from Black Soldier Fly Hermetia illucens


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Melanin From BSF
- -
- homogenization of thawed biomass in a mixer (laboratory disperser HG-202, Vilitek, Moscow, Russia);
- -
- extraction of water-soluble melanin using 10% NaOH in a weight/volume ratio of 1:4 at room temperature for 16 h;
- -
- filtering the alkaline extract through a paper filter (ash less filter paper White Ribbon, Moscow, Russia);
- -
- precipitation of water-soluble melanin from the extract by acidifying it to pH 2.0 by concentrated hydrochloric acid. The precipitate was washed with distilled water to neutral pH, followed by centrifugation at least 3 times, and then dried at 70 °C to constant weight (water-soluble melanin WSM).
- -
- homogenization of thawed biomass in a mixer;
- -
- acid hydrolysis of biomass with sulfuric acid. For this, 20 g of homogenized biomass of the flies was mixed with 150 mL of 25% sulfuric acid and boiled at reflux for 3 h;
- -
- the hydrolysate was cooled to 40 °C and filtered on a Buchner funnel with a water-jet pump, then washed with distilled water. After drying in a desiccator (Medsteclo, Klin, Russia), a black, oily to the touch powder was obtained. The mass of this powder was 5.8 g, which corresponds to a yield approximately 28% (AHM preparation).
2.2. Isolation of BSF Ommochromes
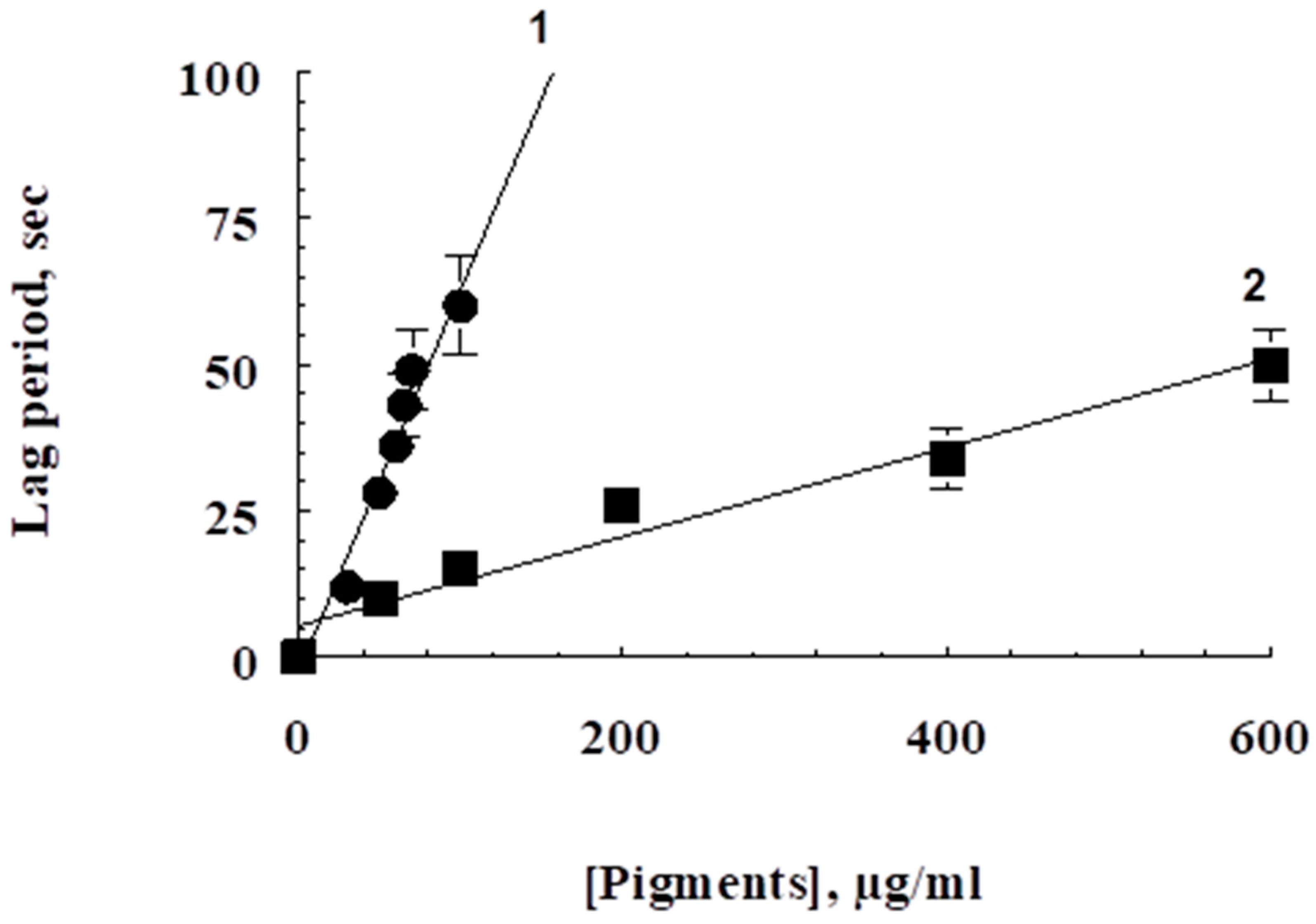
2.3. The Measurements of Antiradical and Antioxidant Activities
2.4. The Measurements of Sorption Capacity of Melanins
2.5. Oxidative Destruction of Pigments
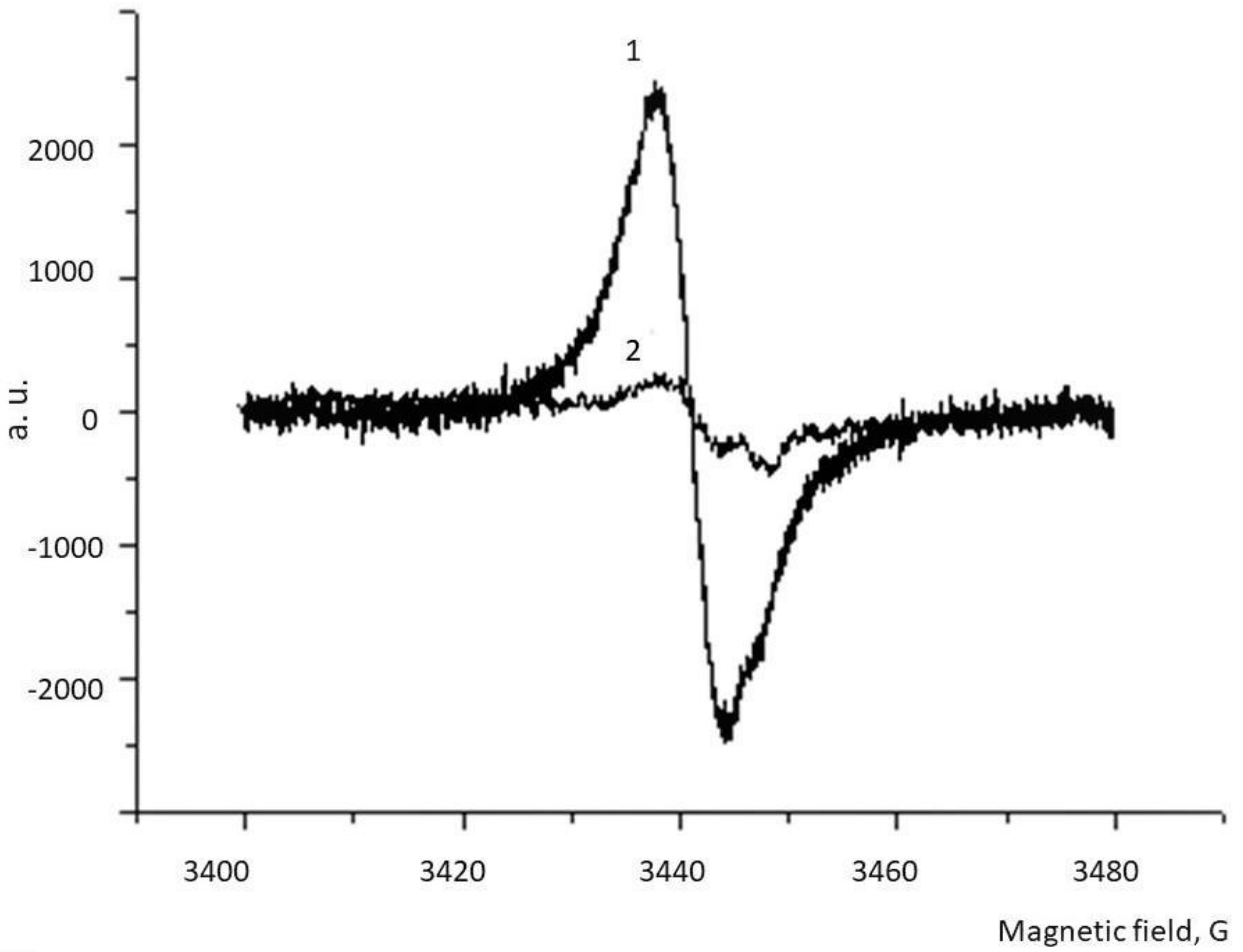
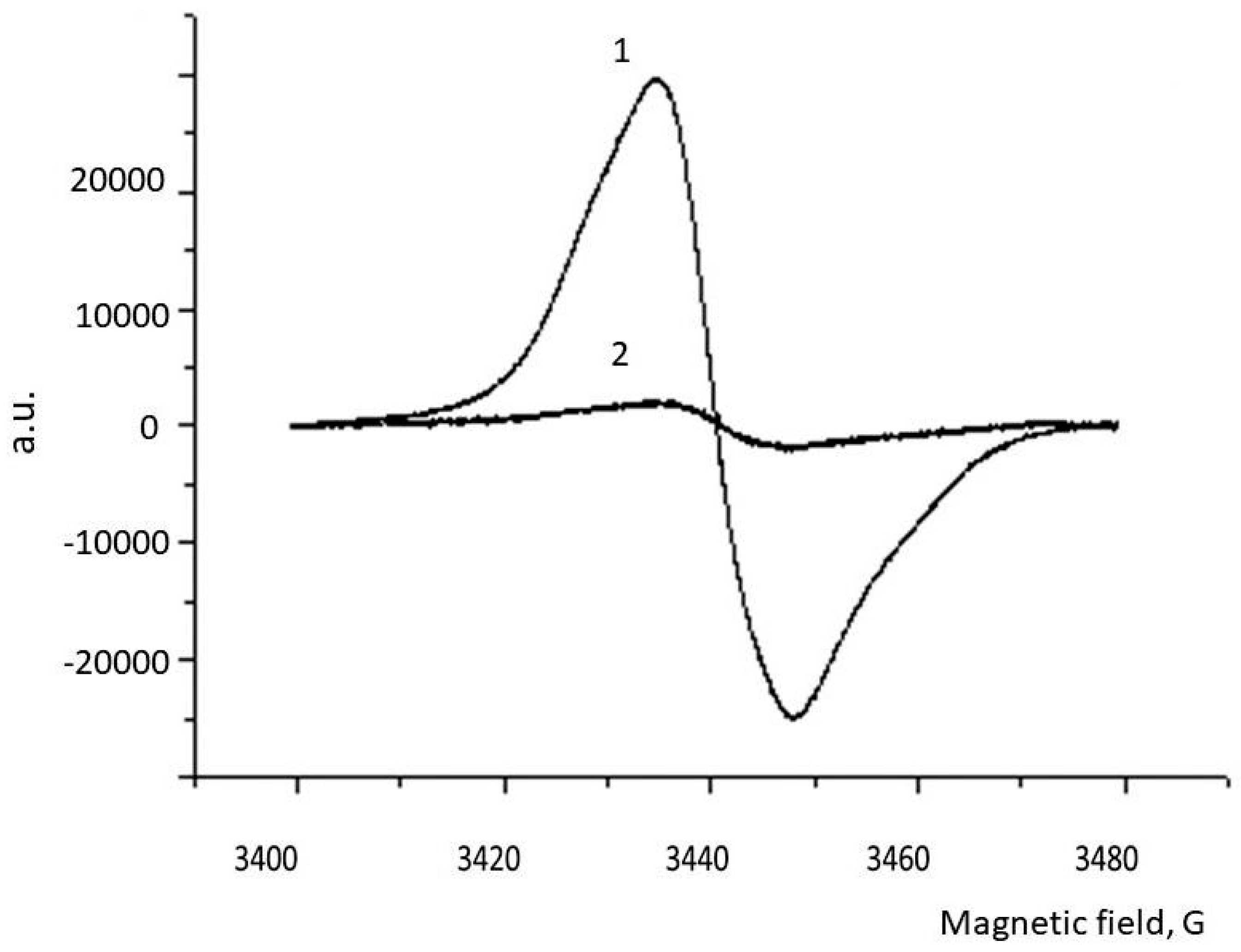
2.6. ESR Spectroscopy
2.7. Statistical Analyses
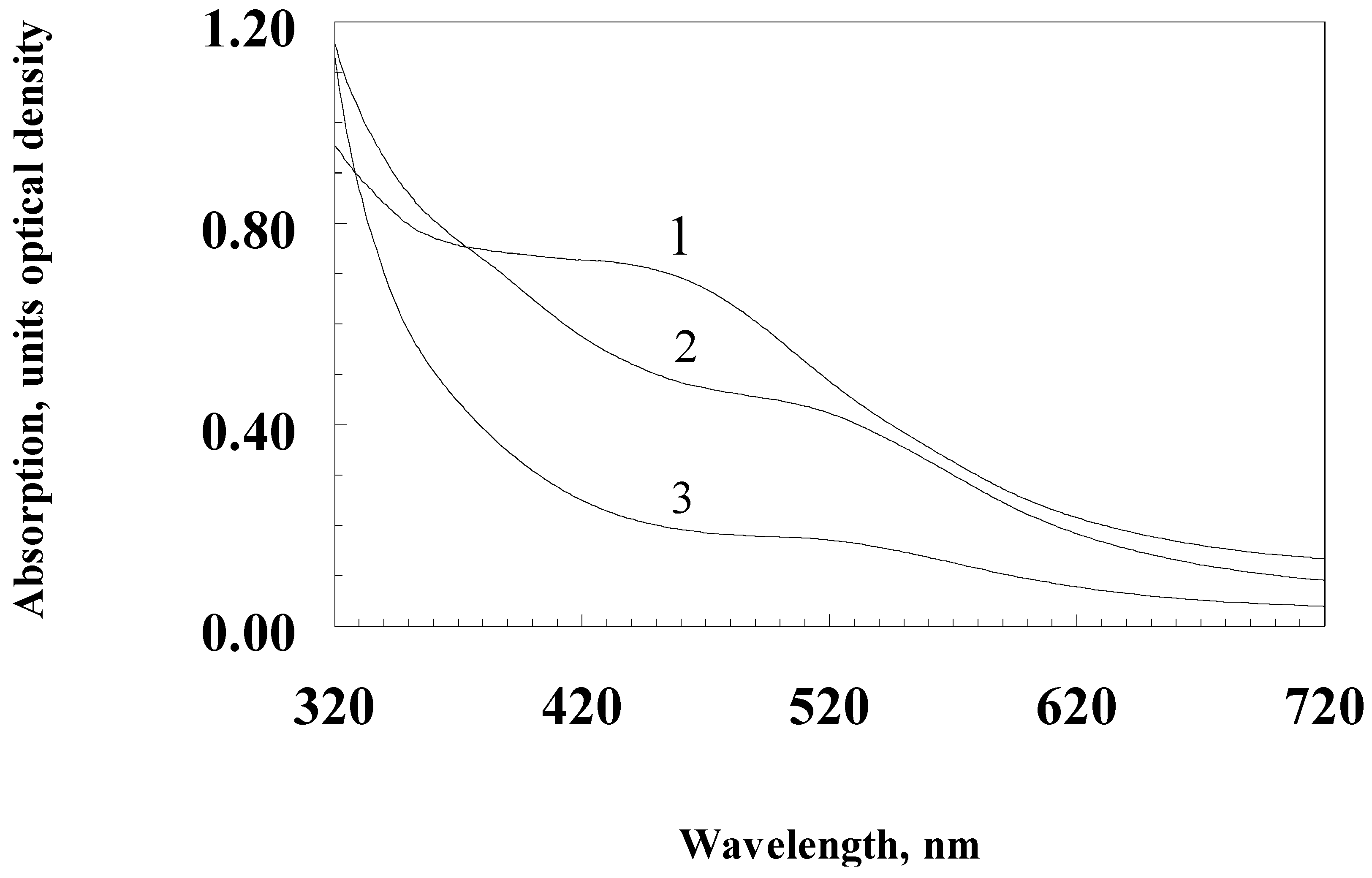
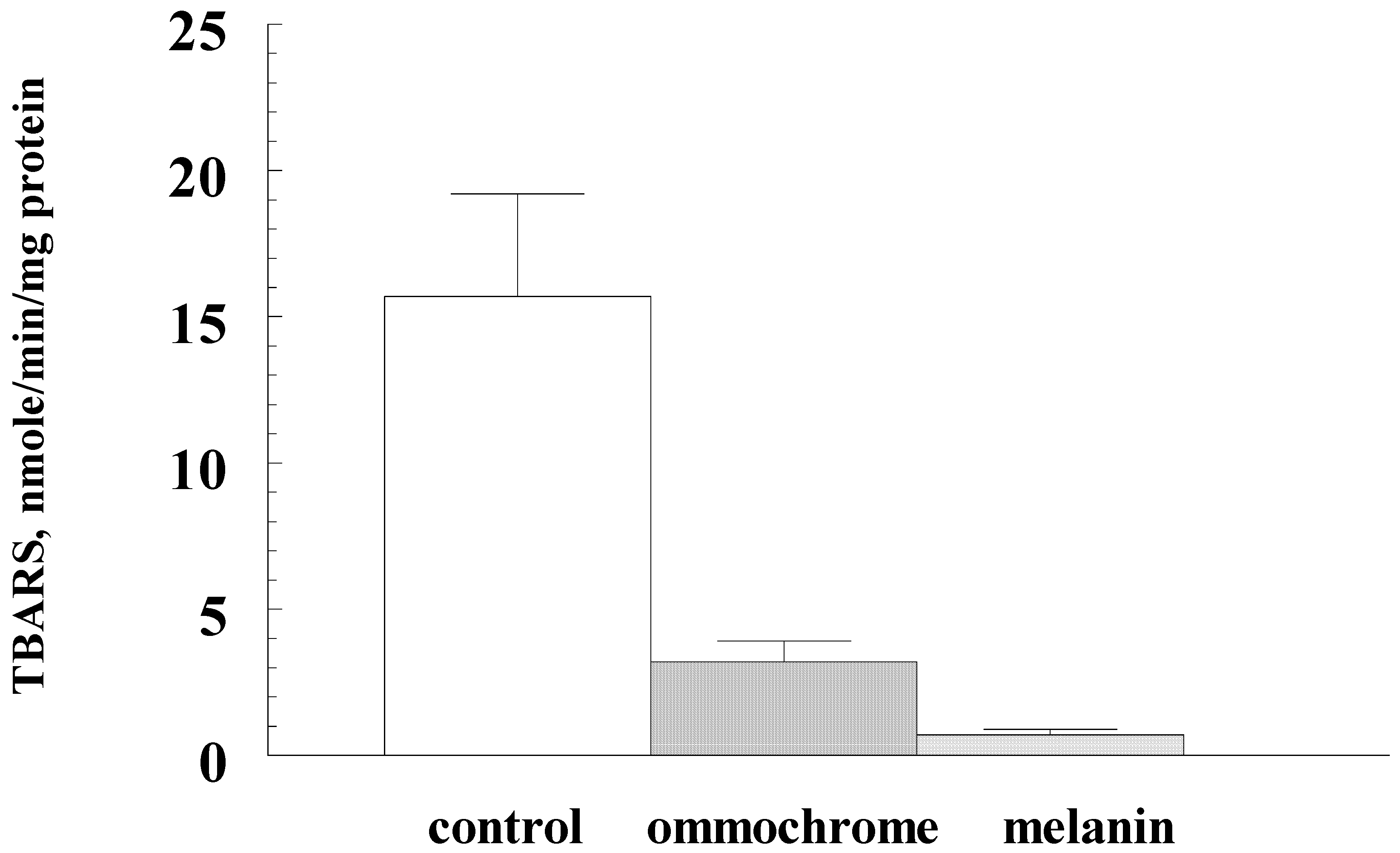
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- NIIR Board of Consultants & Engineers. The Complete Book on Natural Dyes & Pigments; Asia Pacific Business Press Inc: Delhi, India, 2005; p. 448. [Google Scholar]
- Dontsov, A. The Protective Effect of Melanins in Oxidative Stress; Lambert Academic Publishing: Saarbrűcken, Germany, 2014; p. 165. [Google Scholar]
- Ostrovsky, M.A.; Dontsov, A.E. Physiological functions of melanin in the body. Hum. Physiol. 1985, 11, 670–678. [Google Scholar]
- Bagmanyan, I.A.; Myamin, V.E.; Giginyak, Y.G.; Borodin, O.I.; Kurchenko, V.P. The possible role of melanin lichens in the adaptation to extreme factors in Antarctica. BSU 2014, 9, 82–89. [Google Scholar]
- Gessler, N.N.; Egorova, A.S.; Belozerskaya, T.A. Melanin pigments of fungi under extreme environmental conditions (Review). Prikl. Biokhim. Mikrobiol. 2014, 50, 105–113. [Google Scholar]
- Ostrovsky, M.A.; Dontsov, A.E. Vertebrate Eye Melanosomes and Invertebrate Eye Ommochromes as Antioxidant Cell Organelles: Part 2. Biol. Rev. Camb. 2019, 46, 105–116. [Google Scholar] [CrossRef]
- Bilgihan, A.; Bilgihan, M.K.; Akata, R.F.; Aricioglu, A.; Hasanreisoglu, B. Antioxidative role of ocular melanin pigment in the model of lenz induced uveitis. Free Radic. Biol. Med. 1995, 19, 883–885. [Google Scholar] [CrossRef]
- Ostrovsky, M.A.; Sakina, N.L.; Dontsov, A.E. An antioxidative role of ocular screening pigments. Vision Res. 1987, 27, 893–899. [Google Scholar] [CrossRef]
- Corsaro, C.; Scalia, M.; Blanco, A.R.; Aiello, I.; Sichel, G. Melanins in physiological conditions protect against lipo-peroxidation. Pigm. Cell Res. 1995, 8, 279–282. [Google Scholar] [CrossRef]
- Wang, Z.; Dillon, J.; Gaillard, E.R. Antioxidant properties of melanin in retinal pigment epithelium cells. Cell Biochem. Biophys. 2006, 82, 474–479. [Google Scholar]
- Potts, A.M.; Au, P.C. The affinity of melanin for inorganic ions. Exp. Eye Res. 1976, 22, 487–491. [Google Scholar] [CrossRef]
- Bagirov, R.M.; Stukan, R.A.; Dontsov, A.E.; Lapina, V.A.; Ostrovsky, M.A. Gamma-resonance spectroscopic study of ferrum ions binding by melanin granules of the eye pigment epithelium. Biophysics (Moscow) 1986, 31, 469–474. [Google Scholar]
- Butenandt, A.; Schafer, W. Recent Progress in the Chemistry of Natural and Synthetic Coloring Matters and Related Fields; Gore, T.S., Joshi, B.S., Sunthankar, S.V., Tilak, B.D., Eds.; Academic Press: New York, NY, USA, 1962; pp. 13–34. [Google Scholar]
- Figon, F.; Casas, J. Ommochromes in invertebrates: Biochemistry and cell biology. Biol. Rev. Camb. Philos. Soc. 2019, 94, 156–183. [Google Scholar] [CrossRef] [PubMed]
- Riou, M.; Christides, J.-P. Cryptic color change in a crab spider (Misumena vatia): Identification and quantification of precursors and ommochrome pigments by HPLC. J. Chem. Ecol. 2010, 36, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Stavenga, D.G.; Leertouwer, H.L.; Wilts, B.D. Coloration principles of nymphaline butterflies–thin films, melanin, ommochromes and wing scale stacking. J. Exp. Biol. 2014, 217, 2171–2180. [Google Scholar] [CrossRef]
- Panettieri, S.; Gjinaj, E.; John, G.; Lohman, D.J. Different ommochrome pigment mixtures enable sexually dimorphic Batesian mimicry in disjunct populations of the common palmfly butterfly, Elymnias hypermnestra. PLoS ONE 2018, 13, e0202465. [Google Scholar] [CrossRef] [PubMed]
- Gribakin, F.G.; Chesnokova, E.G. Using of eye mutants to study the physiology of insect vision. Adv. Curr. Biol. 1989, 7, 69–82. [Google Scholar]
- Ostrovsky, M.A.; Zak, P.P.; Dontsov, A.E. Vertebrate Eye Melanosomes and Invertebrate Eye Ommochromes as Screening Cell Organelles. Biol. Bull. 2018, 45, 570–579. [Google Scholar] [CrossRef]
- Dontsov, A.E.; Fedorovich, I.B.; Lindström, M.; Ostrovsky, M.A. Comparative study of spectral and antioxidant properties of pigments from the eyes of two Mysis relicta (Crustacea, Mysidacea) populations, with different light damage resistance. J. Comp. Physiol. B 1999, 169, 157–164. [Google Scholar] [CrossRef]
- Insausti, T.C.; LeGall, M.; Lazzari, C.R. Oxidative stress, photodamage and the role of screening pigments in insect eyes. J. Exp. Biol. 2013, 216, 3200–3207. [Google Scholar] [CrossRef]
- Romero, Y.; Martinez, A. Antiradical capacity of ommochromes. J. Mol. Model. 2015, 21, 220. [Google Scholar] [CrossRef]
- Zhuravlev, A.V.; Zakharov, G.A.; Shchegolev, B.F.; Savvateeva-Popova, E.V. Antioxidant properties of kynurenines: Density functional theory calculations. PLoS Comput. Biol. 2016, 12, e1005213. [Google Scholar] [CrossRef]
- Chio, S.-S.; Hyde, J.S.; Sealy, R.C. Paramagnetism in Melanins: pH Dependence. Arch. Biochem. Biophys. 1982, 215, 100–106. [Google Scholar] [CrossRef]
- Borg, D.C. Application of electron spin resonance in biology. In Free Radicals in Biology; Pryor, W.A., Ed.; Acad. Press: Cambridge, MA, USA, 1976; Volume 1, pp. 88–177. [Google Scholar]
- Kalová, M.; Borkovcová, M. Voracious Larvae Hermetia illucens and Treatment of Selected Types of Biodegradable Waste. Acta Univ. Agric. Et Silvicult. Mendel. Brun. 2013, 61, 77–83. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish Offal Recycling by the Black Soldier Fly Produces a Foodstuff High in Omega-3 Fatty Acids. J. World Aquacult. Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). J. Econ. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Porebska-Budny, M.; Sakina, N.; Stepien, K.; Dontsov, A.; Wilczok, T. Antioxidative activity of synthetic melanins. Cardiolipin liposome model. Biochim. Biophys. ACTA Gen. Subj. 1992, 1116, 11–16. [Google Scholar] [CrossRef]
- Teselkin, Y.O.; Babenkova, I.V.; Lyubitsky, O.B.; Klebanov, G.I.; Vladimirov, Y.A. The measurement of serum antioxidant activity in the system hemoglobin–hydrogen peroxide–luminol. J. Med. Chem. 1997, 43, 87–92. [Google Scholar]
- Ottolenghi, A. Interaction of ascorbic acid and mitochondrial lipids. Arch. Biochem. Biophys. 1959, 7, 355–363. [Google Scholar] [CrossRef]
- Sarna, T.; Burke, J.M.; Korytowski, W.; Rozanowska, M.; Skumatz, C.M.B.; Zareba, A.; Zareba, M. Loss of melanin from human RPE with aging: Possible role of melanin photooxidation. Exp. Eye Res. 2003, 76, 89–98. [Google Scholar] [CrossRef]
- Dontsov, A.E.; Sakina, N.L.; Ostrovsky, M.A. Loss of melanin by eye retinal pigment epithelium cells is associated with its oxidative destruction in melanolipofuscin granules. Biochemistry (Moscow) 2017, 82, 916–924. [Google Scholar] [CrossRef]
- Ephrussi, B.; Herold, J.L. Studies of eye pigments of drosophila. I. Methods of extraction and quantitative estimation of the pigment components. Genetics 1944, 39, 148–175. [Google Scholar]
- Pilawa, B.; Zdybel, M.; Chodurek, E. Application of Electron Paramagnetic Resonance Spectroscopy to Examine Free Radicals in Melanin Polymers and Human Melanoma Malignum Cells; Intechopen Limited: London, UK, 2017; Volume 5, pp. 79–103. [Google Scholar] [CrossRef]
- Enochs, W.S.; Nilges, M.J.; Swartz, H.M. A standardized test for the identification and characterization of melanins using electron paramagnetic resonance (EPR) spectroscopy. Pigm. Cell Res. 1993, 6, 91–99. [Google Scholar] [CrossRef]
- Dontsov, A.E.; Lapina, V.A.; Mordvintsev, P.I. Dark and light induced ESR signals of the ommochromes of the invertebrate eye. Biophysics (Moscow) 1985, 30, 6–8. [Google Scholar]
- Caligiani, A.; Marsegli, A.; Leni, G.; Baldassarr, S.; Maistrello, L.; Dossena, A.; Sforza, S. Composition of black soldier fly prepupae and systematic approaches for extraction and fractionation of proteins, lipids and chitin. Food Res. Int. 2018, 105, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Bilinska, B.; Pilava, B.; Zawada, Z.; Wilegala, E.; Wilczok, T.; Dontsov, A.E.; Sakina, N.L.; Ostrovsky, M.A.; Ilyasova, V.B. Electron spin resonance investigations of human retinal pigment epithelium melanosomes from young and old donors. Spectrochimica ACTA 2002, 58, 2257–2264. [Google Scholar] [CrossRef]
- Bolton, J.R. Experimental aspects of biological electron spin resonance studies. In Biological Application of Electron Spin Resonance; Swartz, H.M., Bolton, J.R., Borg, D.C., Eds.; Wiley (Interscience): New York, NY, USA, 1972; p. 11. [Google Scholar]
- Meredith, P.; Sarna, T. The Physical and Chemical Properties of Eumelanin. Pigm. Cell Res. 2006, 19, 572–594. [Google Scholar] [CrossRef]
- Ushakova, N.A.; Dontsov, A.E.; Sakina, N.L.; Brodsky, E.S.; Ratnikova, I.A.; Gavrilova, N.N.; Bastrakov, A.I.; Kozlova, A.A.; Nekrasov, R.V. Melanin properties at the different stages towards life cycle of the fly Hermetia illucens. Ukr. J. Ecol. 2017, 7, 424–431. [Google Scholar] [CrossRef]
- Devic, E.; Fahmi, M.R. Technical Handbook of Domestication and Production of Diptera Black Soldier Fly Hermetia Illucens, Stratiomyidae; Caruso, D., Devic, E., Subamia, I.W., Talamond, P., Baras, E., Eds.; IRD-DKP: Kuala Lumpur, Malaysia, 2013; p. 137. [Google Scholar]
- Becker, E. Über Eigenschaften, Verbreitung und die genetisch-entwicklungsphysiologische Bedeutung der Pigmente der Ommatin- und Ommingruppe (Ommochrome) bei den Arthropoden. Z.i.A. 1942, 80, 157–204. [Google Scholar] [CrossRef]
- Lhoste, J.-M.; Haug, A.; Ptak, M. Electron paramagnetic resonance studies of photoselected triplet molecules. I. Phenoxazine. J. Chem. Phys. 1966, 44, 648–654. [Google Scholar] [CrossRef]
- Bolognese, A.; Bonomo, R.P.; Chillemi, R.; Sciuto, S. Oxidation of 3-hydroxykynurewnine. An EPR investigation. J. Heterocyclic Chem. 1990, 27, 2207–2208. [Google Scholar] [CrossRef]
- Farmer, L.A.; Haidasz, E.A.; Griesser, M.; Pratt, D.A. Phenoxazine: A privileged scaffold for radical–trapping antioxidants. J. Org. Chem. 2017, 82, 10523–10536. [Google Scholar] [CrossRef] [PubMed]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef] [PubMed]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed–a review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Melanin Sample | The Concentration of Paramagnetic Centers, 1017 spin/g Dry Weight | g-Factor | ∆Hpp |
|---|---|---|---|
| WSM | 3.4 ± 0.6 | 2.0038 | 6.2 |
| DL-WSM | 4.4 ± 0.7 | 2.0040 | 6.0 |
| AHM | 15.0 ± 1.7 | 2.0037 | 5.5 |
| DL-AHM | 26.0 ± 2.4 | 2.0037 | 5.5 |
| DOPA-melanin | 51.0 ± 3.5 | 2.0027 | 4.8 |
| Melanin Samples | Antiradical Activity, μM Ascorbate | Maximum Binding of Methylene Blue (MB), mg MB/g Dry Weight |
|---|---|---|
| WSM | 180 ± 16 | - |
| DL-WSM | 250 ± 19 | 390 ± 43 |
| AHM | 18.0 ± 3.5 | - |
| DL-AHM | 32.0 ± 5.0 | 180 ± 17 |
| DOPA-melanin | 6000 ± 460 | 360 ± 45 |
| Sample | The Concentration of Paramagnetic Centers, 1017 Spin/g Dry Weight | g-Factor | ∆Hpp |
|---|---|---|---|
| Suspension of ommochromes in 0.1 M K-phosphate buffer | 16.0 ± 2.4 | 2.0045 | 13.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ushakova, N.; Dontsov, A.; Sakina, N.; Bastrakov, A.; Ostrovsky, M. Antioxidative Properties of Melanins and Ommochromes from Black Soldier Fly Hermetia illucens. Biomolecules 2019, 9, 408. https://doi.org/10.3390/biom9090408
Ushakova N, Dontsov A, Sakina N, Bastrakov A, Ostrovsky M. Antioxidative Properties of Melanins and Ommochromes from Black Soldier Fly Hermetia illucens. Biomolecules. 2019; 9(9):408. https://doi.org/10.3390/biom9090408
Chicago/Turabian StyleUshakova, Nina, Alexander Dontsov, Natalia Sakina, Alexander Bastrakov, and Mikhail Ostrovsky. 2019. "Antioxidative Properties of Melanins and Ommochromes from Black Soldier Fly Hermetia illucens" Biomolecules 9, no. 9: 408. https://doi.org/10.3390/biom9090408
APA StyleUshakova, N., Dontsov, A., Sakina, N., Bastrakov, A., & Ostrovsky, M. (2019). Antioxidative Properties of Melanins and Ommochromes from Black Soldier Fly Hermetia illucens. Biomolecules, 9(9), 408. https://doi.org/10.3390/biom9090408