Omega-3 Fatty Acids Prevent Post-Traumatic Stress Disorder-Induced Memory Impairment

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Induction of Single-Prolonged Stress Model
2.3. Radial Arm Water Maze
2.4. Brain Dissection
2.5. Molecular Assays
2.6. Statistical Analysis
3. Results
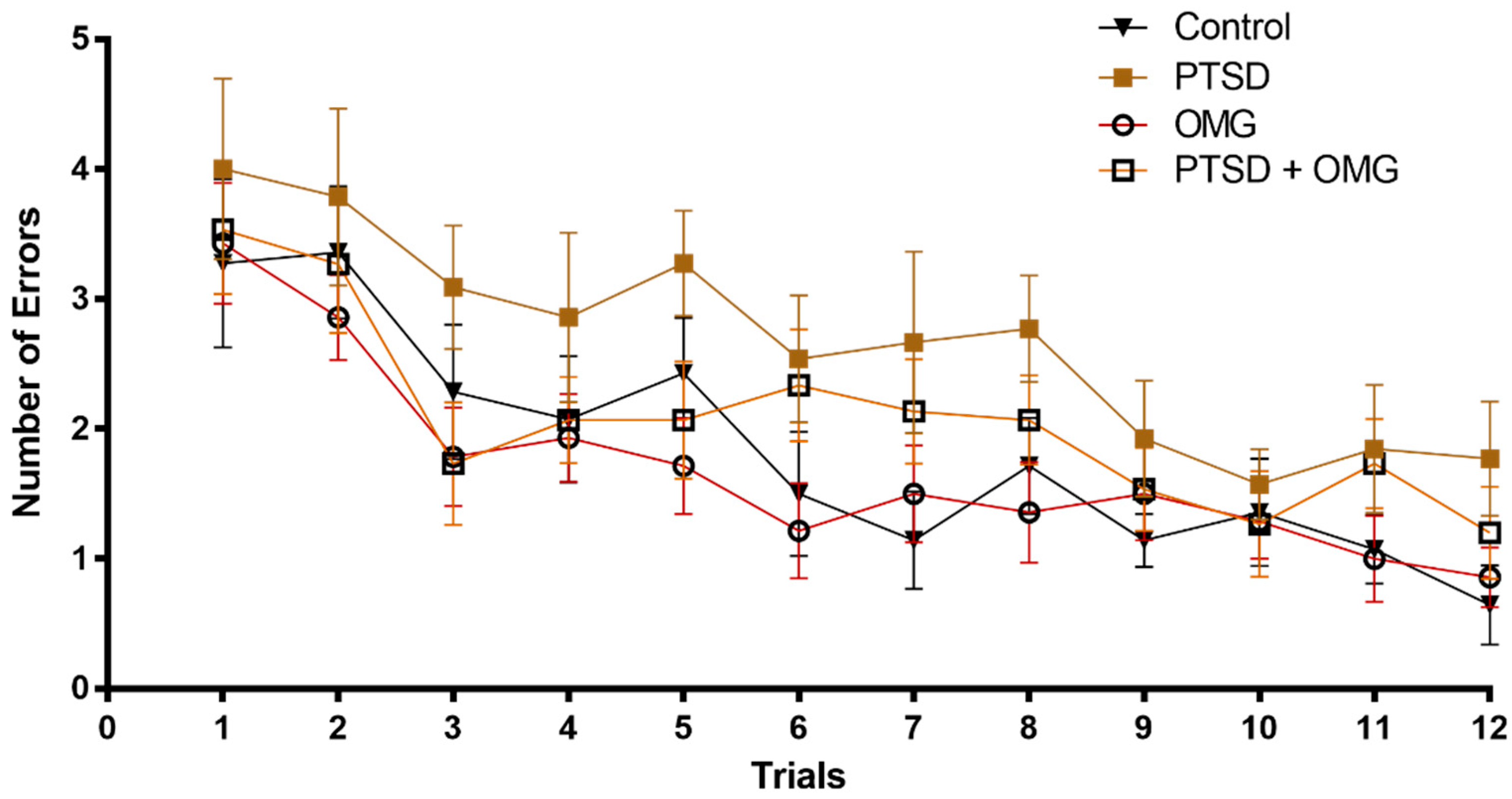
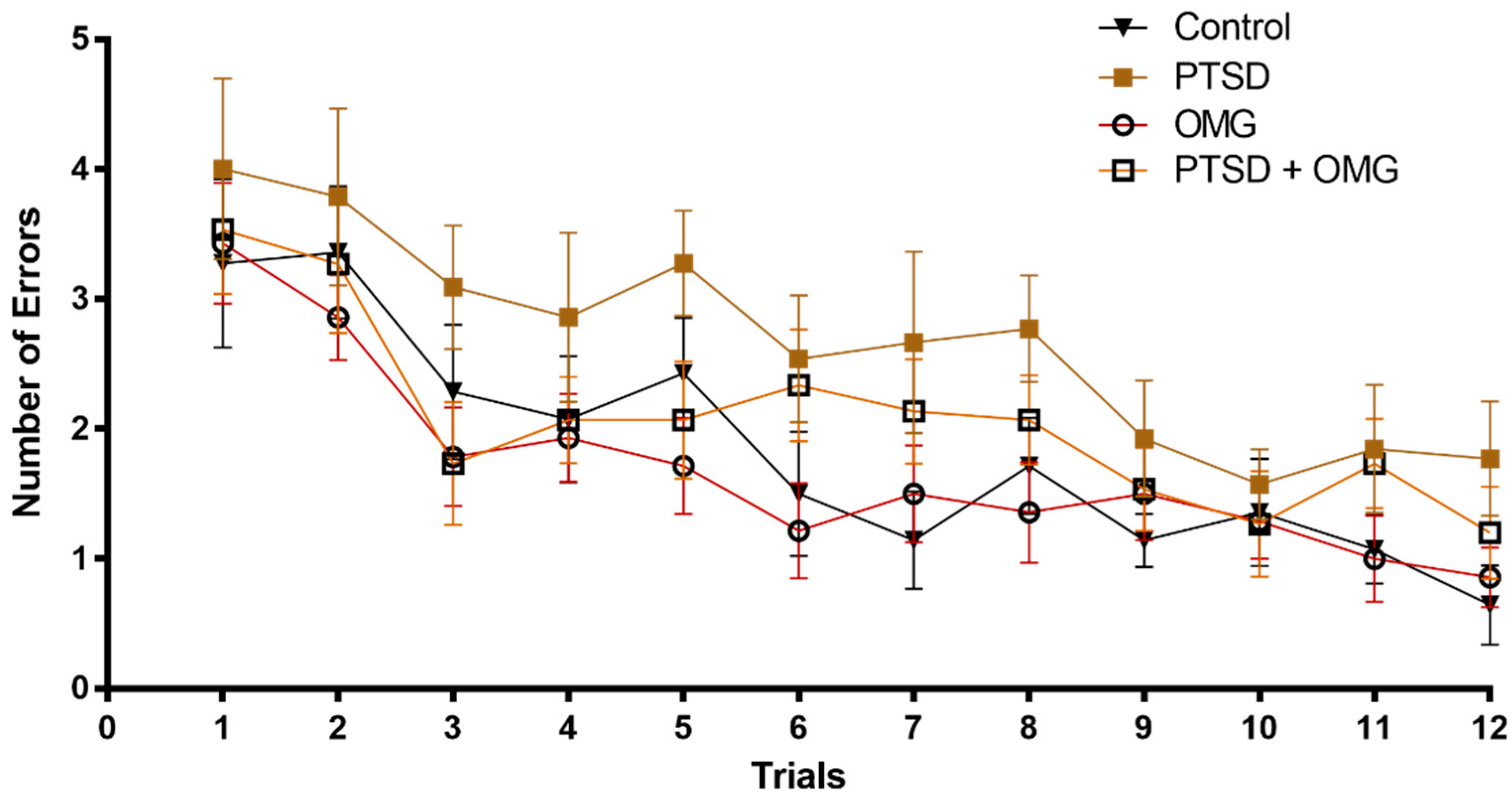
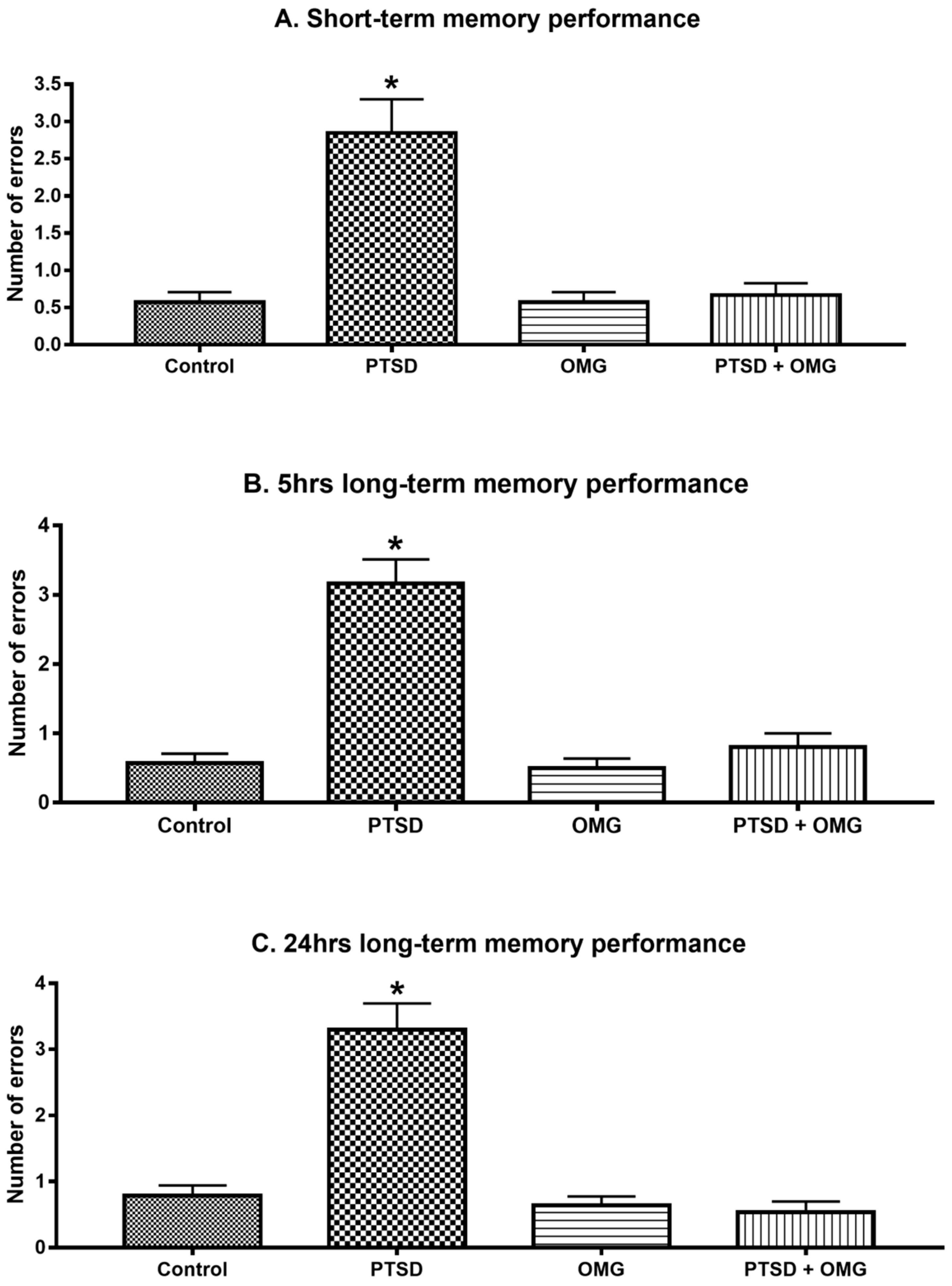
3.1. Effect of Post-Traumatic Stress Disorder and/or Omega-3 on Learning and Memory
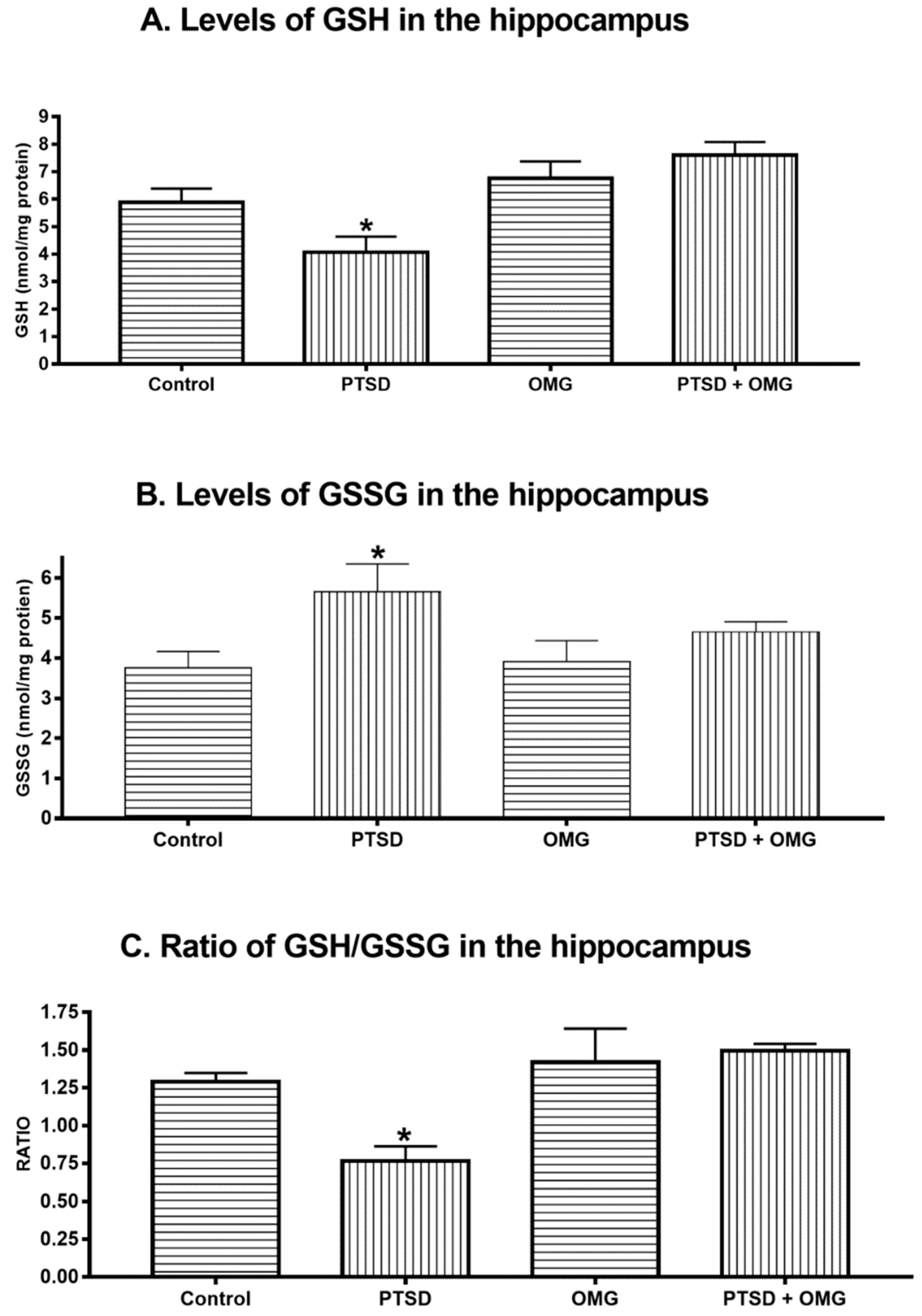
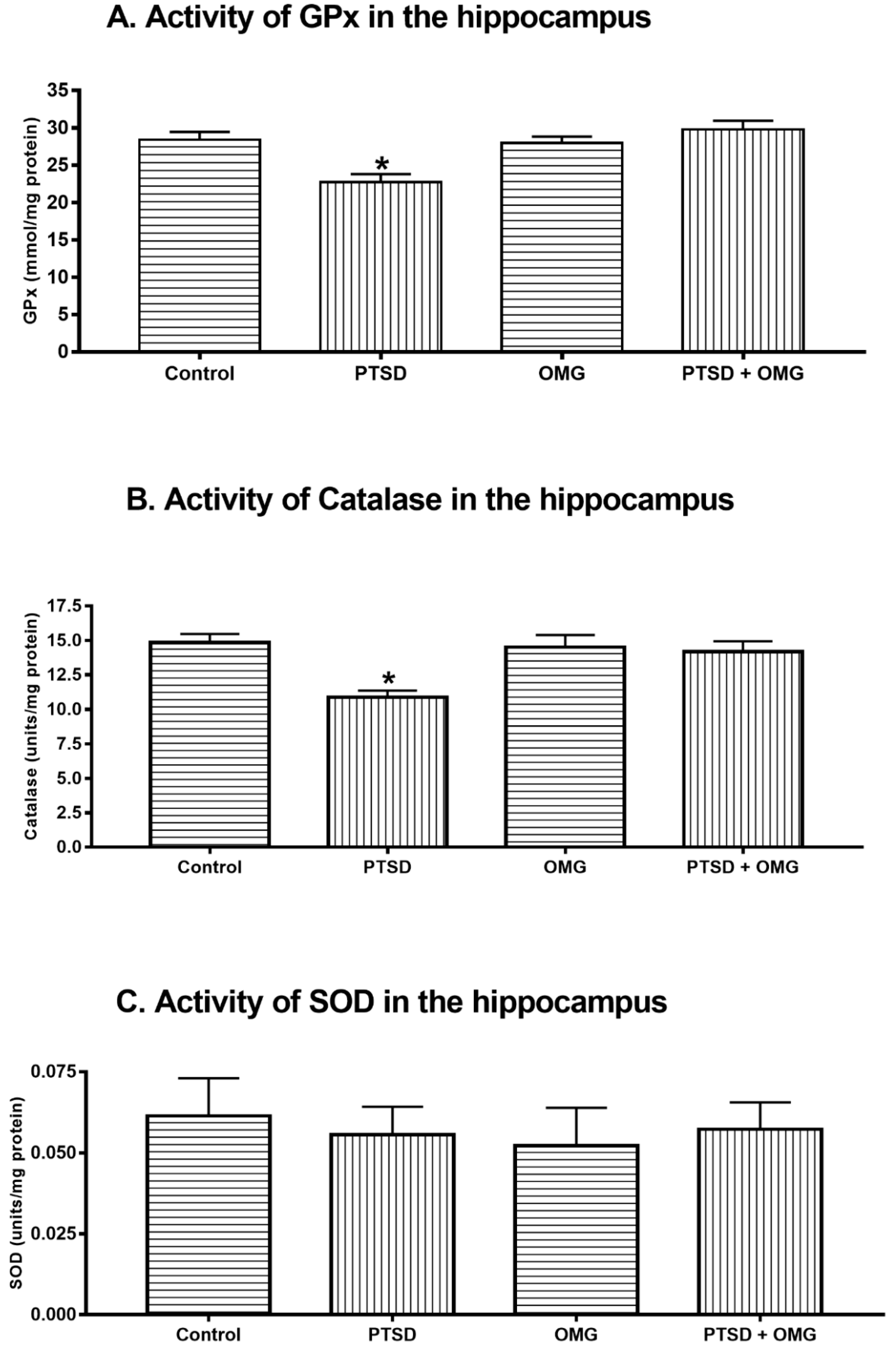
3.2. Effect of Post-Traumatic Stress Disorder and/or Omega-3 on Hippocampus Oxidative Stress Markers
3.3. Effect of Post-Traumatic Stress Disorder and/or Omega-3 on the Hippocampal Thiobarbituric Acid Reactive Species
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Lakhan, S.E.; Vieira, K.F. Nutritional therapies for mental disorders. Nutr. J. 2008, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Resick, P.A.; Monson, C.M.; Rizvi, S.L. Posttraumatic stress disorder. In Psychopathology: History, Diagnosis, and Empirical Foundations; Wiley: Hoboken, NJ, USA, 2013; pp. 244–284. [Google Scholar]
- Peterson, K.C.; Prout, M.F.; Schwarz, R.A. Post-Traumatic Stress Disorder: A Clinician’s Guide; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Atwoli, L.; Stein, D.J.; Koenen, K.C.; McLaughlin, K.A. Epidemiology of posttraumatic stress disorder: Prevalence, correlates and consequences. Curr. Opin. Psychiatry 2015, 28, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Sorce, S.; Krause, K.-H. NOX enzymes in the central nervous system: From signaling to disease. Antioxid. Redox Signal. 2009, 11, 2481–2504. [Google Scholar] [CrossRef] [PubMed]
- Kühn, S.; Gallinat, J. Gray matter correlates of posttraumatic stress disorder: A quantitative meta-analysis. Biol. Psychiatry 2013, 73, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Pitman, R.K.; Rasmusson, A.M.; Koenen, K.C.; Shin, L.M.; Orr, S.P.; Gilbertson, M.W.; Milad, M.R.; Liberzon, I. Biological studies of post-traumatic stress disorder. Nat. Rev. Neurosci. 2012, 13, 769–787. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.B.; Zayfert, C.; Anderson, E. A survey of psychologists’ attitudes towards and utilization of exposure therapy for PTSD. Behav. Res. Ther. 2004, 42, 277–292. [Google Scholar] [CrossRef]
- Ghaffarzadegan, N.; Larson, R.C. Postraumatic Stress Disorder: Five Vicious Cycles that Inhibit Effective Treatment. US Army Med. Dep. J. 2015, 1, 8–13. [Google Scholar]
- Bourre, J.-M. Dietary omega-3 fatty acids and psychiatry: Mood, behaviour, stress, depression, dementia and aging. J. Nutr. Health Aging 2005, 9, 31–38. [Google Scholar]
- Glade, M.J.; Smith, K. Phosphatidylserine and the human brain. Nutrition 2015, 31, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Belkouch, M.; Hachem, M.; Elgot, A.; Van, A.L.; Picq, M.; Guichardant, M.; Lagarde, M.; Bernoud-Hubac, N. The pleiotropic effects of omega-3 docosahexaenoic acid on the hallmarks of Alzheimer’s disease. J. Nutr. Biochem. 2016, 38, 1–11. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; al Subeh, Z.Y.; Khabour, O.F. Evaluating the protective effect of etazolate on memory impairment, anxiety- and depression-like behaviors induced by post traumatic stress disorder. Brain Res. Bull. 2017, 135, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Al-Ibbini, A.M.; Nuseir, K.Q. Prevention of memory impairment induced by post-traumatic stress disorder by cerebrolysin. Psychiatry Res. 2018, 270, 430–437. [Google Scholar] [CrossRef] [PubMed]
- El-Elimat, T.; Alzoubi, K.H.; AbuAlSamen, M.M.; al Subeh, Z.Y.; Graf, T.N.; Oberlies, N.H. Silymarin Prevents Memory Impairments, Anxiety, and Depressive-Like Symptoms in a Rat Model of Post-Traumatic Stress Disorder. Planta Med. 2019, 85, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.V.; Chaby, L.E.; Nazeer, S.; Liberzon, I. Effects of Trauma in Adulthood and Adolescence on Fear Extinction and Extinction Retention: Advancing Animal Models of Posttraumatic Stress Disorder. Front. Behav. Neurosci. 2018, 12, 247. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chang, H.A.; Tai, Y.M.; Chen, T.Y.; Wan, F.J.; Chang, C.C.; Tung, C.S.; Liu, Y.P. Subchronic administration of aripiprazole improves fear extinction retrieval of Pavlovian conditioning paradigm in rats experiencing psychological trauma. Behav. Brain Res. 2019, 362, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Atrooz, F.; Salvi, A.; Salim, S. Behavioral and cognitive impact of early life stress: Insights from an animal model. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 78, 88–95. [Google Scholar] [CrossRef]
- Solanki, N.; Alkadhi, I.; Atrooz, F.; Patki, G.; Salim, S. Grape powder prevents cognitive, behavioral, and biochemical impairments in a rat model of posttraumatic stress disorder. Nutr. Res. 2015, 35, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Lin, C.C.; Chen, C.C.; Tzeng, N.S.; Liu, Y.P. Effects of Oxytocin on Fear Memory and Neuroinflammation in a Rodent Model of Posttraumatic Stress Disorder. Int. J. Mol. Sci. 2018, 19, 3848. [Google Scholar] [CrossRef]
- Chen, I.S.; Subramaniam, S.; Cassidy, M.M.; Sheppard, A.J.; Vahouny, G.V. Intestinal absorption and lipoprotein transport of (omega-3) eicosapentaenoic acid. J. Nutr. 1985, 115, 219–225. [Google Scholar] [CrossRef]
- Filloux, F.; Karras, J.; Imperial, J.S.; Gray, W.R.; Olivera, B.M. The distribution of omega-conotoxin MVIICnle-binding sites in rat brain measured by autoradiography. Neurosci. Lett. 1994, 178, 263–266. [Google Scholar] [CrossRef]
- Salem, N.M.; Lin, Y.H.; Moriguchi, T.; Lim, S.Y.; Salem, N., Jr.; Hibbeln, J.R. Distribution of omega-6 and omega-3 polyunsaturated fatty acids in the whole rat body and 25 compartments. Prostaglandins Leukot. Essent. Fatty Acids 2015, 100, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Takemura, M.; Kiyama, H.; Fukui, H.; Tohyama, M.; Wada, H. Distribution of the omega-conotoxin receptor in rat brain. An autoradiographic mapping. Neuroscience 1989, 32, 405–416. [Google Scholar] [PubMed]
- Alzoubi, K.H.; Rababa’h, A.M.; Al Yacoub, O.N. Tempol prevents post-traumatic stress disorder induced memory impairment. Physiol. Behav. 2018, 184, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Han, F.; Liu, D.J.; Shi, Y.X. Single-prolonged stress induced mitochondrial-dependent apoptosis in hippocampus in the rat model of post-traumatic stress disorder. J. Chem. Neuroanat. 2010, 40, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Patki, G.; Li, L.; Allam, F.; Solanki, N.; Dao, A.T.; Alkadhi, K.; Salim, S. Moderate treadmill exercise rescues anxiety and depression-like behavior as well as memory impairment in a rat model of posttraumatic stress disorder. Physiol. Behav. 2014, 130, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Morinobu, S.; Takei, S.; Fuchikami, M.; Matsuki, A.; Yamawaki, S.; Liberzon, I. Single prolonged stress: Toward an animal model of posttraumatic stress disorder. Depress. Anxiety 2009, 26, 1110–1117. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Khabour, O.F.; Salah, H.A.; Hasan, Z. Vitamin E prevents high-fat high-carbohydrates diet-induced memory impairment: The role of oxidative stress. Physiol. Behav. 2013, 119, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Mayyas, F.A.; Mahafzah, R.; Khabour, O.F. Melatonin prevents memory impairment induced by high-fat diet: Role of oxidative stress. Behav. Brain Res. 2018, 336, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Rawashdeh, N.Q.; Khabour, O.F.; El-Elimat, T.; Albataineh, H.; Al-Zghool, H.M.; Alali, F.Q. Evaluation of the effect of Moringa peregrina extract on learning and memory: Role of oxidative stress. J. Mol. Neurosci. 2017, 63, 355–363. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Khabour, O.F.; Alharahshah, E.A.; Alhashimi, F.H.; Shihadeh, A.; Eissenberg, T. The Effect of Waterpipe Tobacco Smoke Exposure on Learning and Memory Functions in the Rat Model. J. Mol. Neurosci. 2015, 57, 249–256. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Malkawi, B.S.; Khabour, O.F.; El-Elimat, T.; Alali, F.Q. Arbutus andrachne L. Reverses Sleep Deprivation-Induced Memory Impairments in Rats. Mol. Neurobiol. 2018, 55, 1150–1156. [Google Scholar] [PubMed]
- Alzoubi, K.H.; Rababa’h, A.M.; Owaisi, A.; Khabour, O.F. l-Carnitine prevents memory impairment induced by chronic REM-sleep deprivation. Brain Res. Bull. 2017, 131, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Rababa’h, A.M.; Alzoubi, K.H.; Atmeh, A. Levosimendan enhances memory through antioxidant effect in rat model: Behavioral and molecular study. Behav. Pharmacol. 2018, 29, 344–350. [Google Scholar] [CrossRef]
- Aleisa, A.M.; Alzoubi, K.H.; Gerges, N.Z.; Alkadhi, K.A. Nicotine blocks stress-induced impairment of spatial memory and long-term potentiation of the hippocampal CA1 region. Int. J. Neuropsychopharmacol. 2006, 9, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Gerges, N.Z.; Alzoubi, K.H.; Park, C.R.; Diamond, D.M.; Alkadhi, K.A. Adverse effect of the combination of hypothyroidism and chronic psychosocial stress on hippocampus-dependent memory in rats. Behav. Brain Res. 2004, 155, 77–84. [Google Scholar] [CrossRef]
- Mhaidat, N.M.; Alzoubi, K.H.; Khabour, O.F.; Tashtoush, N.H.; Banihani, S.A.; Abdul-Razzak, K.K. Exploring the effect of vitamin C on sleep deprivation induced memory impairment. Brain Res. Bull. 2015, 113, 41–47. [Google Scholar] [CrossRef]
- Eagle, A.L.; Fitzpatrick, C.J.; Perrine, S.A. Single-prolonged stress impairs social and object novelty recognition in rats. Behav. Brain Res. 2013, 256, 591–597. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Khabour, O.F.; Ahmed, M. Pentoxifylline prevents post-traumatic stress disorder induced memory impairment. Brain Res. Bull. 2018, 139, 263–268. [Google Scholar] [CrossRef]
- Alzoubi, K.H.; Mokhemer, E.; Abuirmeileh, A.N. Beneficial effect of etazolate on depression-like behavior and, learning, and memory impairment in a model of Parkinson’s disease. Behav. Brain Res. 2018, 350, 109–115. [Google Scholar] [CrossRef]
- Bremner, J.D.; Scott, T.M.; Delaney, R.C.; Southwick, S.M.; Mason, J.W.; Johnson, D.R.; Innis, R.B.; McCarthy, G.; Charney, D.S. Deficits in short-term memory in posttraumatic stress disorder. Am. J. Psychiatry 1993, 150, 1015–1019. [Google Scholar]
- Whitaker, A.M.; Gilpin, N.W.; Edwards, S. Animal models of post-traumatic stress disorder and recent neurobiological insights. Behavioural. Pharmacol. 2014, 25, 398–409. [Google Scholar]
- Fernandes, J.S.; Mori, M.A.; Ekuni, R.; Oliveira, R.M.; Milani, H. Long-term treatment with fish oil prevents memory impairments but not hippocampal damage in rats subjected to transient, global cerebral ischemia. Nutr. Res. 2008, 28, 798–808. [Google Scholar] [CrossRef] [PubMed]
- D’Ascoli, T.A.; Mursu, J.; Voutilainen, S.; Kauhanen, J.; Tuomainen, T.P.; Virtanen, J.K. Association between serum long-chain omega-3 polyunsaturated fatty acids and cognitive performance in elderly men and women: The Kuopio Ischaemic Heart Disease Risk Factor Study. Eur. J. Clin. Nutr. 2016, 70, 970–975. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Ying, Z.; Gomez-Pinilla, F. Dietary omega-3 fatty acids normalize BDNF levels, reduce oxidative damage, and counteract learning disability after traumatic brain injury in rats. J. Neurotrauma 2004, 21, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Patki, G.; Solanki, N.; Atrooz, F.; Allam, F.; Salim, S. Depression, anxiety-like behavior and memory impairment are associated with increased oxidative stress and inflammation in a rat model of social stress. Brain Res. 2013, 1539, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Javed, H.; Khan, M.M.; Ahmad, A.; Vaibhav, K.; Ahmad, M.E.; Khan, A.; Ashafaq, M.; Islam, F.; Siddiqui, M.S.; Safhi, M.M.; et al. Rutin prevents cognitive impairments by ameliorating oxidative stress and neuroinflammation in rat model of sporadic dementia of Alzheimer type. Neuroscience 2012, 210, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Khabour, O.F.; Salah, H.A.; Rashid, B.E.A. The combined effect of sleep deprivation and western diet on spatial learning and memory: Role of BDNF and oxidative stress. J. Mol. Neurosci. 2013, 50, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Firlag, M.; Kamaszewski, M.; Gaca, K.; Adamek, D.; Balasinska, B. The neuroprotective effect of long-term n-3 polyunsaturated fatty acids supplementation in the cerebral cortex and hippocampus of aging rats. Folia Neuropathol. 2013, 51, 235–242. [Google Scholar] [CrossRef]
- Johnson, E.J.; McDonald, K.; Caldarella, S.M.; Chung, H.Y.; Troen, A.M.; Snodderly, D.M. Cognitive findings of an exploratory trial of docosahexaenoic acid and lutein supplementation in older women. Nutr. Neurosci. 2008, 11, 75–83. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Nishi, D.; Yonemoto, N.; Hamazaki, K.; Hamazaki, T.; Hashimoto, K. Potential role of brain-derived neurotrophic factor in omega-3 fatty acid supplementation to prevent posttraumatic distress after accidental injury: An open-label pilot study. Psychother. Psychosom. 2011, 80, 310–312. [Google Scholar] [CrossRef]
- Budni, J.; Zomkowski, A.D.; Engel, D.; Santos, D.B.; dos Santos, A.A.; Moretti, M.; Valvassori, S.S.; Ornell, F.; Quevedo, J.; Farina, M.; et al. Folic acid prevents depressive-like behavior and hippocampal antioxidant imbalance induced by restraint stress in mice. Exp. Neurol. 2013, 240, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Fontella, F.U.; Siqueira, I.R.; Vasconcellos, A.P.; Tabajara, A.S.; Netto, C.A.; Dalmaz, C. Repeated restraint stress induces oxidative damage in rat hippocampus. Neurochem. Res. 2005, 30, 105–111. [Google Scholar] [CrossRef]
- Erdogan, H.; Fadillioglu, E.; Ozgocmen, S.; Sogut, S.; Ozyurt, B.; Akyol, O.; Ardicoglu, O. Effect of fish oil supplementation on plasma oxidant/antioxidant status in rats. Prostaglandins Leukot. Essent. Fatty Acids 2004, 71, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Sarsilmaz, M.; Songur, A.; Ozyurt, H.; Kus, I.; Ozen, O.A.; Ozyurt, B.; Sogut, S.; Akyol, O. Potential role of dietary omega-3 essential fatty acids on some oxidant/antioxidant parameters in rats’ corpus striatum. Prostaglandins Leukot. Essent. Fatty Acids 2003, 69, 253–259. [Google Scholar] [CrossRef]
- Mayyas, F.; Jaradat, R.; Alzoubi, K.H. Cardiac effects of fish oil in a rat model of streptozotocin-induced diabetes. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 592–599. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alquraan, L.; Alzoubi, K.H.; Hammad, H.; Rababa’h, S.Y.; Mayyas, F. Omega-3 Fatty Acids Prevent Post-Traumatic Stress Disorder-Induced Memory Impairment. Biomolecules 2019, 9, 100. https://doi.org/10.3390/biom9030100
Alquraan L, Alzoubi KH, Hammad H, Rababa’h SY, Mayyas F. Omega-3 Fatty Acids Prevent Post-Traumatic Stress Disorder-Induced Memory Impairment. Biomolecules. 2019; 9(3):100. https://doi.org/10.3390/biom9030100
Chicago/Turabian StyleAlquraan, Laiali, Karem H. Alzoubi, Hana Hammad, Suzie Y. Rababa’h, and Fadia Mayyas. 2019. "Omega-3 Fatty Acids Prevent Post-Traumatic Stress Disorder-Induced Memory Impairment" Biomolecules 9, no. 3: 100. https://doi.org/10.3390/biom9030100
APA StyleAlquraan, L., Alzoubi, K. H., Hammad, H., Rababa’h, S. Y., & Mayyas, F. (2019). Omega-3 Fatty Acids Prevent Post-Traumatic Stress Disorder-Induced Memory Impairment. Biomolecules, 9(3), 100. https://doi.org/10.3390/biom9030100