Apoptosis Induction of Fibroblast-Like Synoviocytes Is an Important Molecular-Mechanism for Herbal Medicine along with its Active Components in Treating Rheumatoid Arthritis

 , and
, and
Abstract
1. Introduction
2. Cell Apoptosis
3. Apoptosis of FLS in RA Patients and RA Pathology
4. Effects and Mechanisms for Herbal Medicine and its Components on FLS Apoptosis in RA
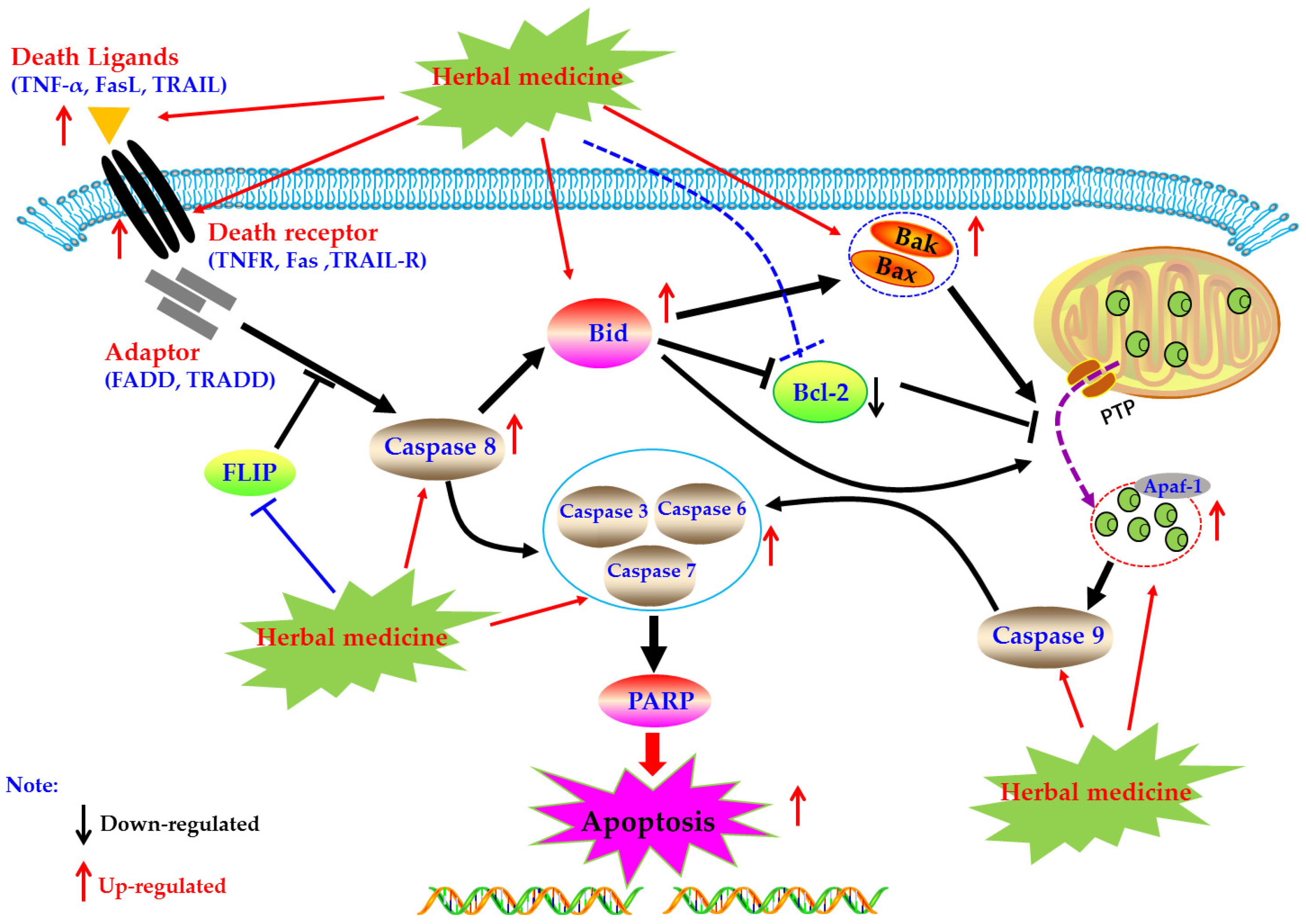
4.1. Death Receptors Mediated Apoptotic Pathway
4.1.1. Herbal Medicine Extracts
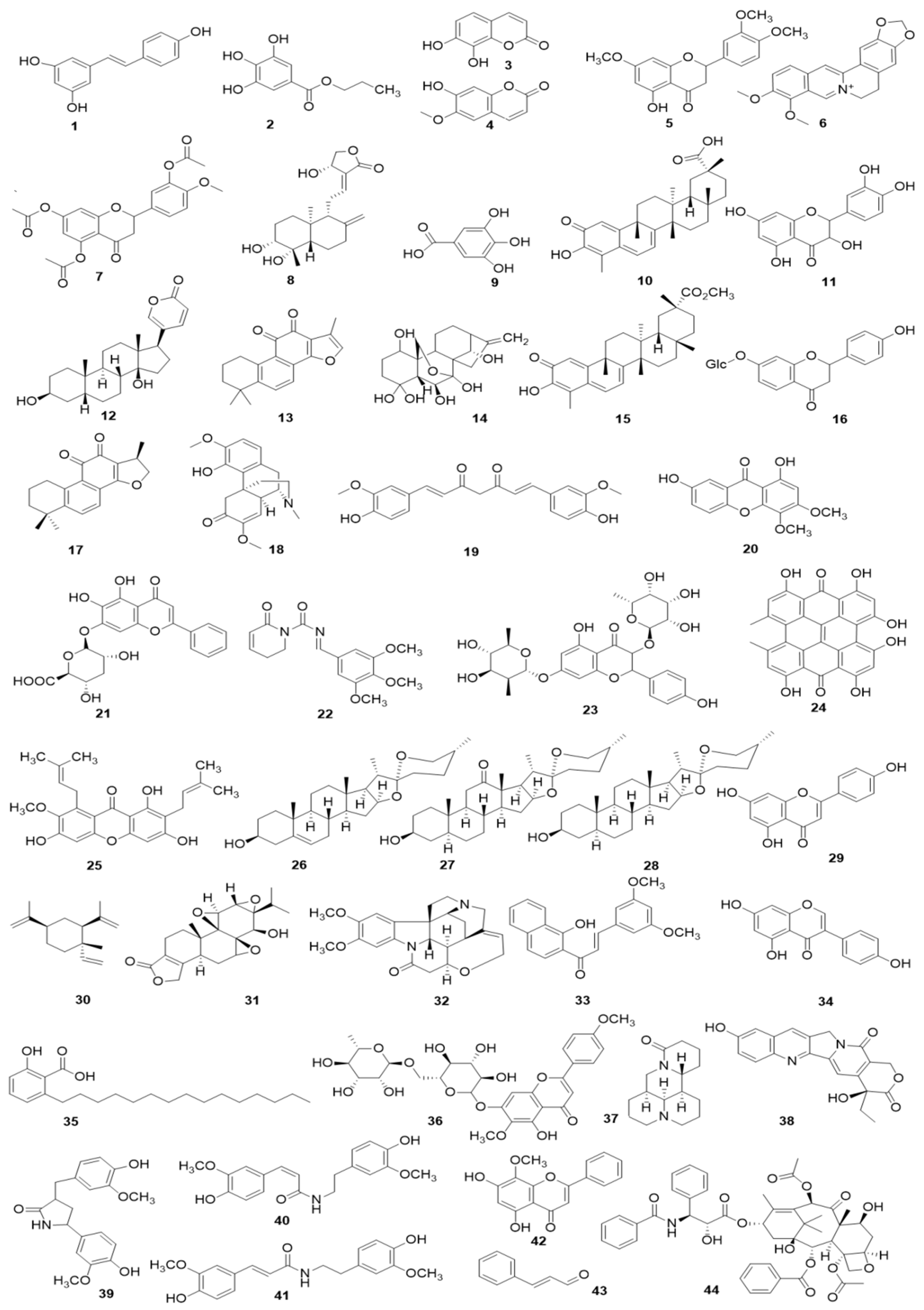
4.1.2. Monomers from Herbal Medicine
4.2. Mitochondrial Dependent Apoptotic Pathway
4.2.1. Herbal Medicine Extracts
4.2.2. Monomers from Herbal Medicine
4.3. NF-κB Mediated Apoptotic Pathways
4.4. MAPK Mediated Apoptotic Pathway
4.5. ERS Mediated Apoptotic Pathway
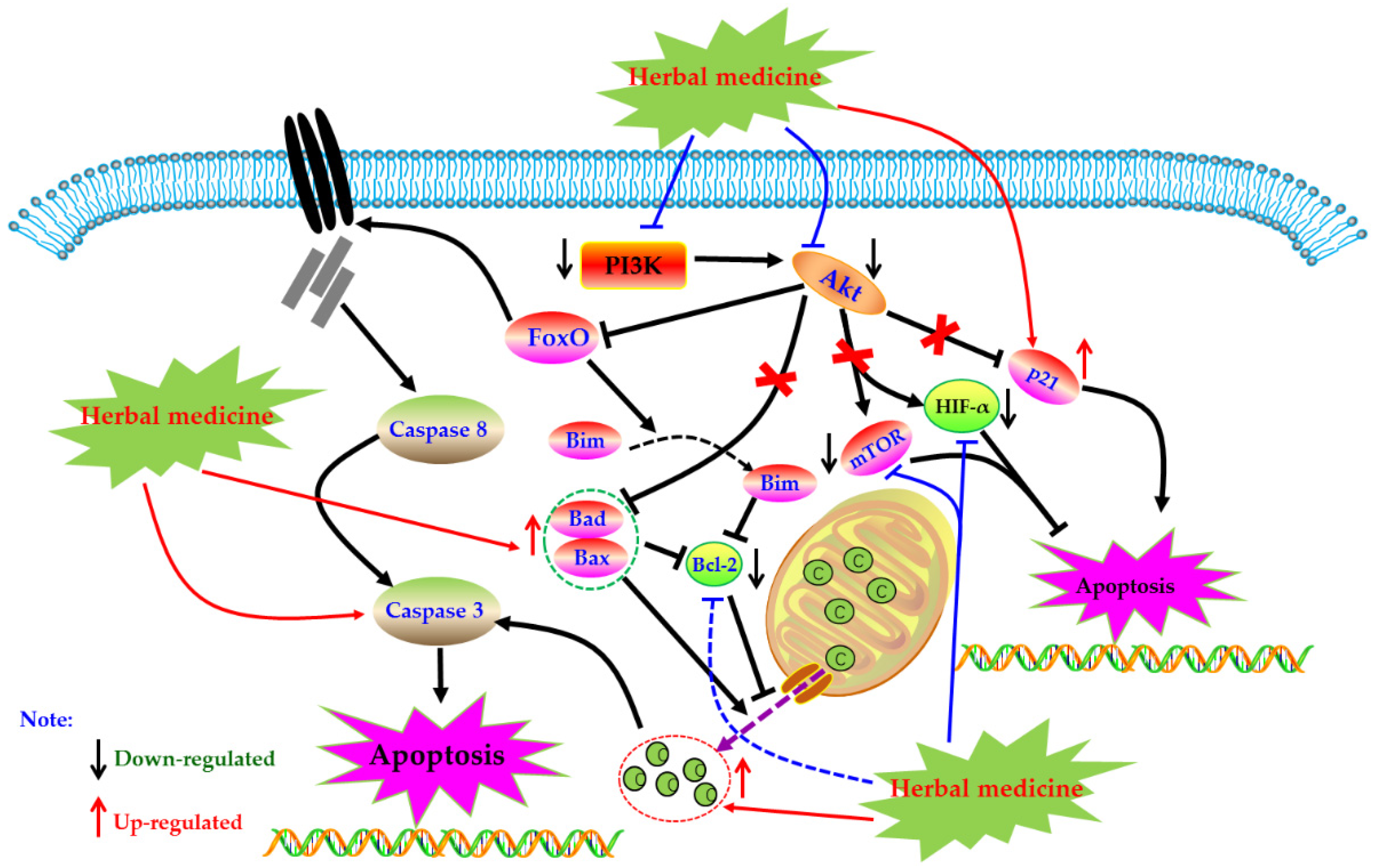
4.6. PI3K-Akt Mediated Apoptotic Pathway
4.6.1. Herbal Medicine Extracts
4.6.2. Monomers from Herbal Medicine
4.7. Other Reported Pathways
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Aletaha, D.; Smolen, J.S. Diagnosis and management of rheumatoid arthritis: A review. JAMA 2018, 320, 1360–1372. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Han, F.; Cui, H.; Huang, J.; Wang, T.; Zhou, Y.; Zhou, J. Matrine suppresses proliferation and induces apoptosis in human cholangiocarcinoma cells through suppression of JAK2/STAT3 signaling. Pharmacol. Rep. 2015, 67, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, Y.; Liu, Y.; Shi, G. The use of biologic therapies in the treatment of rheumatoid arthritis. Curr. Pharm. Biotechnol. 2014, 15, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Cutolo, M.; Sulli, A.; Capellino, S.; Villaggio, B.; Montagna, P.; Seriolo, B.; Straub, R.H. Sex hormones influence on the immune system: Basic and clinical aspects in autoimmunity. Lupus 2004, 13, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.F.; Zhu, S.L.; Tian, A.C.; Xie, X.P. Systematic review of research on disease burden and quality of life in rheumatoid arthritis in China. Chin. J. Evid. Based Med. 2013, 13, 300–307. [Google Scholar]
- McInnes, I.B.; Schett, G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet 2017, 389, 2328–2337. [Google Scholar] [CrossRef]
- Chinese Rheumatology Association. 2018 Chinese guideline for the diagnosis and treatment of rheumatoid arthritis. Chin. J. Int. Med. 2018, 57, 242–250. [Google Scholar]
- Nissen, S.E.; Yeomans, N.D.; Solomon, D.H.; Luscher, T.F.; Libby, P.; Husni, M.E.; Graham, D.Y.; Borer, J.S.; Wisniewski, L.M.; Wolski, K.E.; et al. Cardiovascular safety of Celecoxib, Naproxen, or Ibuprofen for arthritis. N. Engl. J. Med. 2017, 375, 2519–2529. [Google Scholar] [CrossRef]
- Chen, Y.R.; Hsieh, F.I.; Chang, C.Cl.; Chi, N.F.; Wu, H.C.; Chiou, H.Y. Effect on risk of stroke and acute myocardial infarction of nonselective nonsteroidal anti-inflammatory drugs in patients with rheumatoid arthritis. Am. J. Cardiol. 2018, 10, 1271–1277. [Google Scholar] [CrossRef]
- Urushibara, M.; Takayanagi, H.; Koga, T.; Kim, S.; Isobe, M.; Morishita, Y.; Nakagawa, T.; Löeffler, M.; Kodama, T.; Kurosawa, H.; et al. The antirheumatic drug leflunomide inhibits osteoclastogenesis by interfering with receptor activator of NF-kappa B ligand-stimulated induction of nuclear factor of activated T cells c1. Arthritis Rheum. 2004, 50, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.K.; Li, C.; Brennan, F.M. Recent developments in the immunobiology of rheumatoid arthritis. Arthritis Res. Ther. 2008, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.H.; Xu, X.R.; Wei, X.C.; Feng, W.W.; Huang, H.Z.; Liu, H.Y.; Xu, R.C.; Lin, J.Z.; Han, L.; Zhang, D.K. Natural medicines for the treatment of fatigue: Bioactive components, pharmacology, and mechanisms. Pharmacol. Res. 2019, 148, 104409. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Gragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Fang, F.F.; Li, X.Q.; Shu, Z.H.; Jiang, Y.P.; Han, T.; Peng, W.; Zheng, C.J. Matrine Exerts a Strong Anti-Arthritic Effect on Type II Collagen-Induced Arthritis in Rats by Inhibiting Inflammatory Responses. Int. J. Mol. Sci. 2016, 17, 1410. [Google Scholar] [CrossRef]
- Zhang, Q.; Peng, W.; Wei, S.J.; Wei, D.N.; Li, R.L.; Liu, J.; Peng, L.Y.; Yang, S.; Gao, Y.; Wu, C.; et al. Guizhi-Shaoyao-Zhimu decoction possesses anti-arthritic effects on type II collagen-induced arthritis in rats via suppression of inflammatory reactions, inhibition of invasion and migration and induction of apoptosis in synovial fibroblasts. Biomed. Pharmacother. 2019, 118, 109367. [Google Scholar] [CrossRef]
- Soeken, K.L.; Miller, S.A.; Ernst, E. Herbal medicines for the treatment of rheumatoid arthritis: A systematic review. Rheumatology 2003, 42, 652–659. [Google Scholar] [CrossRef]
- Bartox, B.; Firestein, G.S. Fibroblat-like synoviocytes: Key effector cells in rheumatoid arthritis. Immunol. Rev. 2010, 233, 233–255. [Google Scholar] [CrossRef]
- Kaczanowski, S. Apoptosis: Its origin, history, maintenance and the medical implications for cancer and aging. Phys. Biol. 2016, 13, 031001. [Google Scholar] [CrossRef]
- Ntuli, T.M. Apoptosis and Medicine; InTech Prepress: Novi Sad, Croatia, 2012; pp. 71–87. [Google Scholar]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef]
- Horvitz, H.R. Genetic control of programmed cell death in the nematode Caenorhabditis elegans. Cancer Res. 1999, 59, 1701s–1706s. [Google Scholar] [PubMed]
- Fuchs, Y.; Steller, H. Live to die another way: Modes of programmed cell death and the signals emanating from dying cells. Nat. Rev. Mol. Cell Biol. 2015, 16, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Norbury, C.J.; Hickson, I.D. Cellular responses to DNA damage. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 367–401. [Google Scholar] [CrossRef] [PubMed]
- Favaloro, B.; Allocati, N.; Graziano, V.; Di Ilio, C.; De Laurenzi, V. Role of apoptosis in disease. Aging 2012, 4, 330–349. [Google Scholar] [CrossRef] [PubMed]
- Pap, T.; Gay, S. Fibroblasts and fibroblast-like synoviocytes. In Kelly’s Textbook of Rheumatology, 8th ed.; Firestein, G.S., Budd, R.C., Harris, T., McInnes, I.B., Ruddy, S., Sergent, J.S., Eds.; Saunders Elsevier: Philadelphia, PA, USA, 2009; pp. 201–214. [Google Scholar]
- Wang, H.; Jia, X.Z.; Sui, C.J. Effects of thapsigargin on the proliferation and survival of human rheumatoid arthritis synovial cells. Sci. World J. 2014, 2014, 605416. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Abramson, S.B.; Pillinger, M.H. The fibroblast-like synovial cell in rheumatoid arthritis: A key player in inflammation and joint destruction. Clin. Immunol. 2005, 115, 118–128. [Google Scholar] [CrossRef]
- Matsumoto, S.; Müller-Ladner, U.; Gay, R.E.; Nishioka, K.; Gay, S. Ultrastructural demonstration of apoptosis, Fas and Bcl-2 expression of rheumatoid synovial fibroblasts. J. Rheumatol. 1996, 23, 1345–1352. [Google Scholar]
- Perlman, H.; Georganas, C.; Pagliari, L.J.; Koch, A.E.; Haines, K., 3rd; Pope, R.M. Bcl-2 expression in synovial fibroblasts is essential for maintaining mitochondrial homeostasis and cell viability. J. Immunol. 2000, 164, 5227–5235. [Google Scholar] [CrossRef]
- Cha, H.S.; Rosengren, S.; Doyle, D.L.; Firestein, S. PUMA regulation and proapoptotic effects in fibroblast-like synoviocytes. Arthritis Rheum. 2006, 54, 587–592. [Google Scholar] [CrossRef]
- Lamhamedi-Cherradi, S.E.; Zheng, S.J.; Maguschak, K.A.; Peschon, J.; Chen, Y.H. Defective thymocyte apoptosis and accelerated autoimmune diseases in TRAIL-/- mice. Nat. Immunol. 2003, 4, 255–260. [Google Scholar] [CrossRef]
- Korb, A.; Pavenstädt, H.; Pap, T. Cell death in rheumatoid arthritis. Apoptosis 2009, 14, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.W.; Kim, J.H.; Ahn, Y.H.; Seo, J.; Ko, A.; Jeong, M.; Kim, S.J.; Ro, J.Y.; Park, K.M.; Lee, H.W.; et al. Ubiquitination and degradation of the FADD adaptor protein regulate death receptor-mediated apoptosis and necroptosis. Nat. Commun. 2012, 3, 978. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.E.; Budd, R.C.; Desbarats, J.; Hedrick, S.M.; Hueber, A.O.; Newell, M.K. The CD95 receptor: Apoptosis revisited. Cell 2007, 129, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Audo, R.; Combe, B.; Coulet, B.; Morel, J.; Hahne, M. The pleiotropic effect of TRAIL on tumor-like synovial fibroblasts from rheumatoid arthritis patients is mediated by caspases. Cell Death Dis. 2009, 16, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhu, H.; Xu, C.J.; Yuan, J. Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell 1998, 94, 491–501. [Google Scholar] [CrossRef]
- Scatizzi, J.C.; Hutcheson, J.; Bickel, E.; Haines, G.K., III; Perlman, H. Pro-apoptotic Bid is required for the resolution of the effector phase of inflammatory arthritis. Arthritis Res. Ther. 2007, 9, R49. [Google Scholar] [CrossRef]
- Garcia, S.; Conde, C. The role of poly-(ADP-ribose) polymerase-1 in rheumatoid arthritis. Med. Inflamm. 2015, 2015, 837250. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, W.D.; Chen, G.L.; Hu, W.; Han, M.X. Therapeutic effect of Xinfeng Capsule in treating adjuvant arthritis in rats and its effect on fas, fasL and bcl-2 expression in synovial membrane. Chin. J. Integr. Trad. West. Med. 2002, 22, 599–602. [Google Scholar]
- Liu, Q.; Zhu, X.; Xing, Y.; Jiang, H.; Cai, Y.; Lei, J.; Zhao, M.; Yang, M.; Tang, Z.; Yuang, G. Salvia miltiorrhiza induces expressions of Fas/FasL in fibroblat-like synoviocytes from patients with rheumatoid arthritis. Acta Acad. Med. Milt. Tert. 2010, 32, 2452–2454. [Google Scholar]
- Liang, W.; Liang, T.; Zhang, L.L.; Shen, J.M.; Sun, C.C.; Chen, Y.; Ji, Y.Z. Study on Fengshi Bitong Prescription in the treatment of rats with collagen-induced arthritis. J. Emerg. Tradit. Chin. Med. 2017, 26, 83–85. [Google Scholar]
- Peng, W.; Qin, R.X.; Li, X.L.; Zhou, H. Botany, phytochemistry, pharmacology, and potential application of Polygonum cuspidatum Sieb.et Zucc.: A review. J. Ethnopharmacol. 2013, 148, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.J. Effects of Resveratrol on the Expression of Caspase-8 and FLIP of Fibroblast-Like Synoviocytes in Rats Induced by Type II Collagen. Master’s Thesis, North China Coal Medical College, Tangshan, China, 2008. [Google Scholar]
- Palao, G.; Santiago, B.; Galindo, M.; Paya, M.; Ramirez, J.C.; Pablos, J.L. Down-regulation of FLIP sensitizes rheumatoid synovial fibroblasts to Fas-mediated apoptosis. Arthritis Rheum. 2004, 50, 2803–2810. [Google Scholar] [CrossRef] [PubMed]
- Schedel, J.; Gay, R.E.; Kuenzler, P.; Seemaye, C.; Simmen, B.; Michel, B.A.; Gay, S. FLICE-inhibitory protein expression in synovial fibroblasts and at sites of cartilage and bone erosion in rheumatoid arthritis. Arthritis Rheum. 2002, 46, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Palao, G.; Stantiago, B.; Galindo, M.; Rullas, J.; Alcami, J.; Ramirez, J.C.; Pablos, J.L. Fas activation of a proinflammatory program in rheumatoid synoviocytes and its regulation by FLIP and Caspase 8 signaling. Arthritis Rheum. 2006, 54, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Peng, A. The changes of expression of fas/fasl mRNA in rheumatoid arthritis fibroblast-like synoviocytes induced by Radix paeoniae rubra 801. J. Hunan Normal Univ. (Med. Sci.) 2013, 10, 69–72. [Google Scholar]
- Peng, A.; Cai, Y.; Jiang, H. Apoptosis of rheumatoid arthritis fibroblast- like synoviocytes induced by Radix Paeoniae rubra 801. Prac. Prev. Med. 2013, 20, 1138–1140. [Google Scholar]
- Liu, W.; Yang, H.J.; Wu, Y.H.; Zhang, L.; Xue, B. Research progress on effects of Chinese materia medica and active components on proliferation and apoptosis of fibroblast-like synoviocytes in rheumatoid arthritis. Chin. Trad. Herb. Drugs. 2016, 47, 844–850. [Google Scholar]
- Zheng, M. Daphnetin Induces Apoptosis in Fibroblast-like Synoviocytes from Collagen-Induced Arthritic Rat and its Mechanism. Ph.D. Thesis, Nanchang University, Nanchang, China, 2016. [Google Scholar]
- Newmeyer, D.D.; Farschon, D.M.; Reed, J.C. Cell-free apoptosis in Xenopus egg extracts: Inhibition by Bcl-2 and requirement for an organelle fraction enriched in mitochondria. Cell 1994, 79, 353–364. [Google Scholar] [CrossRef]
- Dai, L.; Li, T.; Zhu, L.J. Effect and related mechanism of seaweed polysaccharide on apoptosis of fibroblast like synovlocytes in patients with rheumatoid arthritis. Chin. J. Integr. Trad. West. Med. 2011, 31, 961–966. [Google Scholar]
- Ma, Y.M.; Li, Y.Y.; Wang, Y.H.; Gao, L.; Zhang, L.; Zhou, R. Effects of Fengshining Capsule on synoviocyte apoptosis of CIA rats in mitochondria pathway. Chin. J. Trad. Chin. Med. Pharm. 2013, 28, 1985–1988. [Google Scholar]
- Gao, X.P.; Lu, G.S. Medicated serum of Duhuo Jisheng Tang has effect on proliferation and apoptosis of fibroblast-like synoviocytes in rats with adjuvant-incluced arthritis. J. New Chin. Med. 2018, 50, 1–5. [Google Scholar]
- Ci, B.; Wang, W.; Ni, Y. Inhibitory effect of Saposhnikovia divaricate polysaccharide on fibroblast-like synoviocytes from rheumatoid arthritis rat in vitro. Pak. J. Pharm. Sci. 2018, 31, 2791–2798. [Google Scholar] [PubMed]
- Wu, L.L.; Wang, X.P.; Zhang, Q.S.; Zhu, J.; Yuan, L.; Xiang, Y. Effects and mechanism of Pterocarya Hupehensis Skan extracts on proliferation and apoptosis of MH7A cells. Chin. Immunol. J. 2019, 35, 653–658. [Google Scholar]
- Li, Y.; Dai, Y.; Liu, M. Scopoletin induces apoptosis of fibroblast-like synoviocytes from adjuvant arthritis rats by a mitochondrial-dependent pathway. Drug Dev. Res. 2009, 70, 378–385. [Google Scholar] [CrossRef]
- Li, R.; Cai, L.; Xie, X.F.; Peng, L.; Li, J. 7,3′-dimethoxy hesperetin induces apoptosis of fibroblast-like synoviocytes in rats with adjuvant arthritis through Caspase 3 activation. Phytother. Res. 2010, 24, 1850–1856. [Google Scholar] [CrossRef]
- Wang, X.H. Effect of Berberine on Human Rheumatoid Arthritis Fibroblast-Like Synoviocytes and Its Underlying Molecular Mechanim. Ph.D. Thesis, Shandong University, Jinan, China, 2011. [Google Scholar]
- Ren, D.Y.; Xu, T.; Huang, C.; Li, R.; Wang, Y.L.; Yu, M.Z.; Huang, Y.; Li, J. The influence of 5,7,3′-triacetyl hesperetin on Jak2/Stat3 signalling pathway and related apoptosis proteins of fibroblast-like synoviocytes in rats with adjuvant arthritis. Chin. Pharmacol. Bull. 2012, 28, 1068–1073. [Google Scholar]
- Yan, J.; Chen, Y.; He, C. Andrographolide induces cell cycle arrest and apoptosis in human rheumatoid arthritis fibroblast-like synoviocytes. Cell Biol. Toxicol. 2012, 28, 47–56. [Google Scholar] [CrossRef]
- Yoon, C.H.; Chung, S.J.; Lee, S.W.; Park, Y.B.; Lee, S.K.; Park, M.C. Gallic acid, a natural polyphenolic acid, induces apoptosis and inhibits proinflammatory gene expressions in rheumatoid arthritis fibroblast-like synoviocytes. Joint Bone Spine 2013, 80, 274–279. [Google Scholar] [CrossRef]
- Xu, Z.T. Celastrol-Introduced Apoptosis, Cell Cycle Arrest and DNA Damage in Rheumatoid Arthritis Fibroblast-Like Synovial. Ph.D. Thesis, Fujian University of Traditional Chinese Medicine, Fuzhou, China, 2013. [Google Scholar]
- Peng, X. The Effects of Quercetin on the Apoptosis of Human Rheumatoid Arthritis Fibroblast-Like Synoviocytes and Associated Molecular Mechanisms. Ph.D. Thesis, Zhengzhou University, Zhengzhou, China, 2013. [Google Scholar]
- Chang, Y.W.; Zhao, Y.F.; Cao, Y.L. Bufalin exerts inhibitory effects on IL-1β-mediated proliferation and induces apoptosis in human rheumatoid arthritis fibroblast-like synoviocytes. Inflammation 2014, 37, 1552–1559. [Google Scholar] [CrossRef]
- Jie, L.; Du, H.; Huang, Q.; Wei, S.; Huang, R.; Sun, W. Tanshinone IIA Induces Apoptosis in Fibroblast-Like Synoviocytes in Rheumatoid Arthritis via Blockade of the Cell Cycle in the G2/M Phase and a Mitochondrial Pathway. Biol. Pharm. Bull. 2014, 37, 1366–1372. [Google Scholar] [CrossRef] [PubMed]
- Shang, C.H.; Zhang, Q.Q.; Zhou, J.H. Oridonin inhibits cell proliferation and induces apoptosis in rheumatoid arthritis fibroblast-like synoviocytes. Inflammation 2016, 39, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Q.; Song, X.B.; Cao, W. Autophagy and mitochondrial dysfunction in adjuvant-arthritis rats treatment with resveratrol. Sci. Rep. 2016, 6, 32928. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.Y.; Jin, X. Effect of Resveratrol on proliferation and apoptosis of rheumatoid arthritis fibroblast-like synoviocytes. Jiangsu Med. J. 2015, 41, 2182–2184. [Google Scholar]
- Athar, M.; Back, J.H.; Kopelovich, L.; Bickers, D.R.; Kim, A.L. Multiple molecular targets of resveratrol: Anti-carcinogenic mechanisms. Arch. Biochem. Biophys. 2009, 486, 95–102. [Google Scholar] [CrossRef]
- Chuang, C.Y.; Liu, H.C.; Wu, L.C.; Chen, C.Y.; Chang, J.T.; Hsu, S.L. Gallic acid induces apoptosis of lung fibroblasts via a reactive oxygen species-dependent ataxia telangiectasia mutated-p53 activation pathway. J. Agric. Food Chem. 2010, 58, 2943–2951. [Google Scholar] [CrossRef]
- Gao, W.J. Pristimerin Inhibits Rheumatoid Arthritis Fibroblast-like Synoviocytes Proliferation and Underlying Mechanisms. Master’s Thesis, Southern Medical University, Guangzhou, China, 2016. [Google Scholar]
- Zhai, K.F.; Duan, H.; Cui, C.Y.; Cao, Y.Y.; Si, J.L.; Yang, H.J.; Wang, Y.C.; Cao, W.G.; Gao, G.Z.; Wei, Z.J. Liquiritin from Glycyrrhiza uralensis Attenuating Rheumatoid Arthritis via Reducing Inflammation, Suppressing Angiogenesis, and Inhibiting MAPK Signaling Pathway. J. Agric. Food Chem. 2019, 67, 2856–2864. [Google Scholar] [CrossRef]
- Sun, H.N.; Luo, Y.H.; Meng, L.Q.; Piao, X.J.; Wang, Y.; Wang, J.R.; Wang, H.; Zhang, Y.; Li, J.Q.; Xu, W.T.; et al. Cryptotanshinone induces reactive oxygen species mediated apoptosis in human rheumatoid arthritis fibroblast like synoviocytes. Int. J. Mol. Med. 2019, 43, 1067–1075. [Google Scholar] [CrossRef]
- Li, Q.; Yang, S.; Yang, S.; Xin, F.; Wang, M. Anti-inflammatory activity of phlomisoside F isolated from Phlomis younghusbandii Mukerjee. Int. Immunopharmacol. 2015, 28, 724–730. [Google Scholar] [CrossRef]
- Beg, A.A.; Sha, W.C.; Bronson, R.T. Embryonic lehality and liver degeneration in mice lacking the ReLa component of NF-κB. Nature 1995, 376, 167. [Google Scholar] [CrossRef]
- Swierkot, J.; Nowak, B.; Czarny, A.; Zaczynska, E.; Sokolik, R.; Madej, M.; Korman, L. The activity of JAK/STAT and NF-kappaB in patients with rheumatoid arthritis. Adv. Clin. Exp. Med. 2016, 25, 709–717. [Google Scholar] [CrossRef]
- Yin, G.; Wang, Y.; Cen, X.M. Lipid peroxidation-mediated inflammation promotes cell apoptosis through activation of NF-kappaB pathway in rheumatoid arthritis synovial cells. Med. Inflamm. 2015, 2015, 460310. [Google Scholar] [CrossRef] [PubMed]
- Xai, Z.B.; Meng, F.R.; Fang, Y.X. Inhibition of NF-κB signaling pathway induces apoptosis and suppresses proliferation and angiogenesis of human fibroblast-like synovial cells in rheumatoid arthritis. Medicine 2018, 97, e10920. [Google Scholar]
- Zhang, Y.; Fang, Y.F.; Wang, Y.; Li, J.Y.; Huang, X.L. Effects of sinomenine on in vitro proliferation and apoptosis of fibroblast like synoviocyte from patients with rheumatoid arthritis. Acta Acad. Med. Milt. Tert. 2007, 29, 1179–1182. [Google Scholar]
- Fang, Y.F.; Zhang, Y.; Wang, Y. Effects of sinomenine on synovioctye apoptosis, cell cycle and change of bcl-2 protein in patients with rheumatoid arthritis. Chin. Remed. Clin. 2008, 8, 261–264. [Google Scholar]
- Zhang, H.C.; Liu, M.X.; Wang, E.P.; Lin, Z.; Lv, G.F.; Chen, X. Effect of sinomenine on the expression of rheumatoid arthritis fibroblast-like synoviocytes MyD88 and TRAF6. Genet. Mol. Res. 2015, 14, 18928–18935. [Google Scholar] [CrossRef] [PubMed]
- Shang, W.; Zhao, Z.; Zheng, Z.; Guo, J.; Cai, H. Effect of curcumin on rats with adjuvant arthritis fibroblast-like synovial cell proliferation. Guid. J. Trad. Chin. Med. Pharm. 2010, 16, 92–94. [Google Scholar]
- Kloesch, B.; Becker, T.; Dietersdorfer, E. Anti-inflammatory and apoptotic effects of the polyphenol curcumin on human fibroblast-like synoviocytes. Int. Immunopharmacol. 2013, 15, 400–405. [Google Scholar] [CrossRef]
- Fang, Z.Y.; He, D.Y.; Yu, B.; Liu, B.; Liu, F.; Zuo, J.P.; Li, Y.X.; Lin, Q.; Zhou, X.D.; Wang, Q.W. High-throughput study of the effects of Celastrol on activated fibroblast-like synoviocytes from patients with rheumatoid arthritis. Genes 2017, 8, 221. [Google Scholar] [CrossRef]
- Ji, C.L.; Jiang, H.; Tao, M.Q.; Wu, W.T.; Jiang, J.; Zuo, J. Selective regulation of IKKb/NF-kB pathway involved in proliferation inhibition of HFLS-RA cells induced by 1,7-dihydroxyl-3,4-dimethoxylxanthone. Kaohsiung J. Med. Sci. 2017, 33, 486–495. [Google Scholar] [CrossRef]
- Zuo, J.; Dou, D.Y.; Wang, H.F.; Zhu, Y.H.; Li, Y.; Luan, J.J. Reactive oxygen species mediated NF-κB/p38 feedback loop implicated in proliferation inhibition of HFLS-RA cells induced by 1,7-dihydroxy-3,4-dimethoxyxanthone. Biomed. Pharmacother. 2017, 94, 1002–1009. [Google Scholar] [CrossRef]
- Ma, W.K.; Ning, Q.Y.; Li, D.X.; Yao, X.M.; Zhou, J.; Hou, L.; Tang, F.; Liang, J.; Huang, Y. Effects of medicated serum frozen powder of Jinwu Jiangu Formula on synovial cell proliferation in rheumatoid arthritis. Chin. J. Trad. Chin. Med. Pharm. 2017, 32, 1739–1742. [Google Scholar]
- Ning, Q.Y.; Zhou, J.; Li, D.X.; Yao, X.M.; Ma, W.K.; Hou, L. Effect of Miao Medicine Jinwu Jiangu decoction on expression of NF-κB p65, IKK-α and IKK-β in FLS of RA. Chin. J. Exp. Trad. Med. Formul. 2017, 23, 154–159. [Google Scholar]
- Zhang, K.; Gao, S.; Guo, J.Y.; Ni, G.H.; Chen, Z.; Li, F.; Zhu, X.L.; Wen, Y.B.; Guo, Y.X. Hypericin-photodynamic therapy inhibits proliferation and induces apoptosis in human rheumatoid arthritis fibroblast-like synoviocytes cell line MH7A. Iran. J. Basic Med. Sci. 2018, 21, 130–137. [Google Scholar] [PubMed]
- Xu, S.; Xiao, Y.; Zeng, S.; Zou, Y.; Qiu, Q.; Huang, M.; Zhan, Z.; Liang, L.; Yang, Y.; Xu, H. Piperlongumine inhibits the proliferation, migration and invasion of fibroblast-like synoviocytes from patients with rheumatoid arthritis. Inflamm. Res. 2018, 67, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, Q. Kaempferitrin inhibits proliferation, induces apoptosis, and ameliorates inflammation in human rheumatoid arthritis fibroblast-like synoviocytes. Phytother. Res. 2019, 33, 1726–1735. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Z.; Ge, C.X.; Wang, G.Z.; Zhao, J.; Jiang, N. Effects of Baicalin on human fibroblast like synoviocytes (rheumatoid arthritis). J. Qiqihar Univ. Med. 2017, 38, 129–131. [Google Scholar]
- Zuo, J.; Yin, Q.; Wang, Y.W.; Li, Y.; Lu, M.; Xiao, Z.G.; Wang, G.D.; Luan, J.J. Inhibition of NF-κB pathway in fibroblast-like synoviocytes by α-mangostin implicated in protective effects on joints in rats suffering from adjuvant-induced arthritis. Int. Immunopharmacol. 2018, 56, 78–89. [Google Scholar] [CrossRef]
- Arthur, J.S.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef]
- Zhu, C.; Qi, X.; Chen, Y.; Sun, B.; Dai, Y.; Gu, Y. PI3K/Akt and MAPK/ERK1/2 signaling pathways are involved in IGF-1-induced VEGF-C upregulation in breast cancer. J. Cancer Res. Clin. Oncol. 2011, 137, 1587–1594. [Google Scholar] [CrossRef]
- Li, Z.; Li, C.; Du, L.; Zhou, Y.; Wu, W. Human chorionic gonadotropin beta induces migration and invasion via activating ERK1/2 and MMP-2 in human prostate cancer DU145 cells. PLoS ONE 2013, 8, e54592. [Google Scholar]
- Stanciu, Y.; Wang, R.; Kentor, N.; Burke, S.; Watkins, G.; Kress, I.; Reynolds, E.; Klann, M.R.; Angiolieri, J.W.; Johnson, D.B. Persistent activation of ERK contributes to glutamate-induced oxidative toxicity in a neuronal cell line and primary cortical neuron cultures. J. Biol. Chem. 2000, 275, 12200–12206. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Z.; Lian, J.R.; Kong, X.Y.; Lin, N. Effects of triptolide on cell proliferation and regulation of RAS MAPKs pathway in synoviocytes induces by tumor necrosis factor. Chin. J. Chin. Mater. Med. 2010, 35, 888–891. [Google Scholar]
- Jiang, Y.; Wang, X.; Hu, D. Furanodienone induces G0/G1 arrest and causes apoptosis via the ROS/MAPKs-mediated caspase-dependent pathway in human colorectal cancer cells: A study in vitro and in vivo. Cell Death Dis. 2017, 25, e2815. [Google Scholar] [CrossRef]
- Fang, J.; Nakamura, H.; Iyer, A.K. Tumor-targeted induction of oxystress for cancer therapy. J. Drug Target. 2007, 15, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Ding, C. Drug evaluation: VX-702, a MAP kinase inhibitor for rheumatoid arthritis and acute coronary syndrome. Curr. Opin. Investig. Drugs. 2006, 7, 1020–1025. [Google Scholar] [PubMed]
- Ralph, J.A.; Morand, E.F. MAPK phosphatases as novel targets for rheumatoid arthritis. Exp. Opin. Ther. Targets 2008, 12, 795–808. [Google Scholar] [CrossRef]
- Dong, C.; Davis, R.J.; Flavell, R.A. MAP kinases in the immune response. Annu. Rev. Immunol. 2002, 20, 55–72. [Google Scholar] [CrossRef]
- Thalhamer, T.; McGrath, M.A.; Harnett, M.M. MAPKs and their relevance to arthritis and inflammation. Rheumatology 2008, 47, 409–414. [Google Scholar] [CrossRef]
- Liagre, B.; Leger, D.; Vergne-Salle, P.; Beneytout, J.L. MAP kinase subtypes and Akt regulate diosgenin-induced apoptosis of rheumatoid synovial cells in association with COX-2 expression and prostanoid production. Int. J. Mol. Med. 2007, 19, 113–122. [Google Scholar] [CrossRef]
- Liagre, B.; Vergne-Salle, P.; Leger, D.; Beneytout, J.L. Inhibition of human rheumatoid arthritis synovial cell survival by hecogenin and tigogenin is associated with increased apoptosis, p38 mitogen-activated protein kinase activity and upregulation of cyclooxygenase-2. Int. J. Mol. Med. 2007, 20, 451–460. [Google Scholar] [CrossRef][Green Version]
- Shin, G.C.; Kim, C.; Lee, J.M.; Cho, W.S.; Lee, S.G.; Jeong, M.; Cho, J.; Lee, K. Apigenin-induced apoptosis is mediated by reactive oxygen species and activation of ERK1/2 in rheumatoid fibroblast-like synoviocytes. Chem. Biol. Interact. 2009, 182, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Xia, Y.; Li, X.; Ou-yang, Z.; Chen, J.W. Selective modulation of MAPKs contribute to the anti-proliferative and anti-inflammatory activities of 1,7-dihydroxy-3,4-dimethoxyxanthone in rheumatoid arthritis-derived fibroblast-like synoviocyte MH7A cells. J. Ethnopharmacol. 2015, 168, 248–254. [Google Scholar] [CrossRef]
- Zou, S.; Wang, C.; Cui, Z.; Guo, P.; Meng, Q.; Shi, X.; Gao, Y.; Yang, G.; Han, Z. β-Elemene induces apoptosis of human rheumatoid arthritis fibroblast-like synoviocytes via reactive oxygen species-dependent activation of p38 mitogen-activated protein kinase. Pharmacol. Rep. 2015, 68, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Jia, S.; Pan, W.P. Study on inhibitory effects of Triptolide on the proliferation of fibroblast-like synovial cells from patients with rheumatoid arthritis in vitro. Chin. Pharm. 2015, 26, 4357–4359. [Google Scholar]
- Tang, M. Study on Brucine’s Effect on Proliferation and JNK Regulation in MAPK Signal Transduction Pathway on RA HFLS Induced by TNF-α. Master Thesis, Luzhou Medical College, Luzhou, China, 2014. [Google Scholar]
- Zhang, L.B.; Man, Z.T.; Li, W.; Zhang, W.; Wang, X.Q.; Sun, S. Calcitonin protects chondrocytes from lipopolysaccharide-induced apoptosis and inflammatory response through MAPK/Wnt/NF-κB pathways. Mol. Immunol. 2017, 87, 249–257. [Google Scholar] [CrossRef]
- Faitova, J.; Krekac, D.; Hrstka, R. Endoplasmic reticulum stress and apoptosis. Cell. Mol. Biol. Lett. 2006, 11, 488–505. [Google Scholar] [CrossRef]
- Kim, R.; Emi, M.; Tanabe, K.; Murakami, S. Role of the unfolded protein response in cell death. Apoptosis 2006, 11, 5–13. [Google Scholar] [CrossRef]
- Mihailidou, C.; Papazian, I.; Papavassiliou, A.G.; Kiaris, H. CHOP-dependent regulation of p21/waf1 during ER stress. Cell. Physiol. Biochem. 2010, 25, 761–766. [Google Scholar] [CrossRef]
- Mccullough, K.D.; Martindale, J.L.; Klotz, L.O. Gadd153 sensitizes cells to endoplasmic reticulum stress by down-regulating Bcl2 and perturbing the cellular redox state. Mol. Cell. Biol. 2001, 21, 1249–1259. [Google Scholar] [CrossRef]
- Jeong, M.; Cho, J.; Shin, J.I.; Jeon, Y.J.; Kim, J.H.; Lee, S.J.; Kim, E.S.; Lee, K. Hempseed oil induces reactive oxygen species-and C/EBP homologous protein-mediated apoptosis in MH7A human rheumatoid arthritis fibroblast-like synovial cells. J. Ethnopharmacol. 2014, 154, 745–752. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeon, Y.J.; Cho, J.; Shin, J.I.I.; Baek, C.Y.; Lim, Y.; Koh, D.; Shin, S.Y.; Lee, Y.H.; Lee, K. A novel synthetic chalcone derivative promotes caspase-dependent apoptosis through ROS generation and activation of the UPR in MH7A cells. Genes Genom. 2015, 37, 1051–1059. [Google Scholar] [CrossRef]
- Shin, J.I.; Jeon, Y.J.; Lee, S. Apoptotic and anti-inflammatory effects of Eupatorium japonicum Thunb. in rheumatoid arthritis fibroblast-like synoviocytes. BioMed Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zheng, Y.; Yang, J. Resveratrol alleviates inflammatory injury and enhances the apoptosis of fibroblast like synoviocytes via mitochondrial dysfunction and ER stress in rats with adjuvant arthritis. Mol. Med. Rep. 2019, 20, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.J.; Foster, J.G.; Ward, S.G. PI3K isoforms as drug targets in inflammatory diseases: Lessons from pharmacological and genetic strategies. Curr. Opin. Investig. Drugs. 2009, 10, 1151–1162. [Google Scholar] [PubMed]
- Smith, M.D.; Weedon, H.; Papangelis, V.; Walker, J.; Roberts-Thomson, P.J.; Ahern, M.J. Apoptosis in the rheumatoid arthritis synovial membrane: Modulation by disease-modifying anti-rheumatic drug treatment. Rheumatology 2010, 49, 862–875. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.D. Apoptosis a relevant therapeutic target in rheumatoid arthritis? Rheumatology 2004, 43, 405–407. [Google Scholar] [CrossRef][Green Version]
- Hongtao, L.; Pope, R.M. The role of apoptosis in rheumatoid arthritis. Curr. Opin. Pharmacol. 2003, 3, 317–322. [Google Scholar]
- Yu, D.H.; Liu, L.; Lu, F.; Yang, C.; Xue, H.H.; Liu, S.M. Effect of total saponin of dioscoreae nipponicae rhizoma on PI3K/Akt signal pathway by in rIL-1β induced fibroblast-like synoviocytes. Chin. J. Exp. Trad. Med. Formaul. 2012, 18, 199–202. [Google Scholar]
- Pan, D.M.; Wang, Q.; Cai, X.D.; Guo, T.F.; Wei, Z.Q.; Han, L.Y.; Jiang, Y.B.; Liu, M.Y.; Liu, X.B.; Guo, W.J.; et al. Duanteng Yimu Decoction induces apoptosis through inhibition of PI3K/Akt pathway in rheumatoid arthritis fibroblast-like synoviocytes. Chin. J. Trad. Chin. Med. Pharm. 2018, 33, 2051–2055. [Google Scholar]
- Zou, L.H.; Hou, C.F.; Chen, Z.H.; Hua, Z.; Wei, S. Effects of Shuangwuxuanbi granule on the apoptosis of rheumatoid arthritis fibroblast-like synoviocytes and PI3K/Akt signaling pathway. Med. J. Chin. PLA 2018, 43, 721–726. [Google Scholar]
- Liu, Y.; Jiang, S.S.; Chen, X.Y.; Yang, X.; Yan, F.L.; Cao, F.; Yang, C.F.; Liang, J. Effect of Hei Gu Teng Zhui Feng Huo Luo Capsule on PI3K/AKT/HIF-1α protein signaling pathway in rheumatoid arthritis rats. Chin. J. Immun. 2019, 35, 2206–2212. [Google Scholar]
- Zhang, Y.; Zhang, X.Z.; Shen, W.G.; Xu, J.X.; Ma, Y.Y. Effect of Genistein on the proliferation and apoptosis of fibroblast-like synoviocytes from rats with collagen II induced arthritis. Chin. Pharmacol. Bull. 2011, 27, 1161–1165. [Google Scholar]
- Li, G.; Liu, Y.; Meng, F.; Xia, Z.; Wu, X.; Fang, Y.; Zhang, C.; Liu, D. Tanshinone IIA promotes the apoptosis of fibroblast-like synoviocytes in rheumatoid arthritis by up-regulating lncRNA GAS5. Biosci. Rep. 2018, 38, BSR20180626. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.H. Therapeutic Effects of Anacardic Acid on Rheumatoid Arthritis and Associated Molecular Mechanisms. Ph.D. Thesis, Zhengzhou University, Zhengzhou, China, 2018. [Google Scholar]
- Zuo, R.T.; Wang, X.B. Effects of juglone on cell proliferation and apoptosis in rheumatoid arthritis fibroblast-like synoviocytes. Chin. Trad. Pat. Med. 2019, 41, 2334–2339. [Google Scholar]
- Feng, Y.J.; Wang, Z.X.; Yang, J. Effects of diosgenin on cell apoptosis and cell cycle of fibroblast like synoviocytes in rheumatoid arthritis via PI3K/Akt signaling pathway. Immunol. J. 2019, 35, 752–758. [Google Scholar]
- Wang, L.; Wang, N.; Zhao, Q.; Zhang, B.; Ding, Y. Pectolinarin inhibits proliferation, induces apoptosis, and suppresses inflammation in rheumatoid arthritis fibroblast-like synoviocytes by inactivating the phosphatidylinositol 3 kinase/protein kinase B pathway. J. Cell. Biochem. 2019, 120, 15202–15210. [Google Scholar] [CrossRef]
- Yang, Y.; Dong, Q.; Li, R. Matrine induces the apoptosis of fibroblast-like synoviocytes derived from rats with collagen-induced arthritis by suppressing the activation of the JAK/STAT signaling pathway. Int. J. Mol. Med. 2017, 39, 307–316. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, G.; Wei, X. Celastrol induced DNA damage, cell cycle arrest, and apoptosis in human rheumatoid fibroblast-like synovial cells. Am. J. Chin. Med. 2013, 41, 615–628. [Google Scholar] [CrossRef]
- Ren, L.L. Effect of 10-Hydroxycamptothecine on Inhibiting Fibroblast-Like Synocytes in Patients with Rheumatoid Arthritis. Master Thesis, Zhengzhou University, Zhengzhou, China, 2016. [Google Scholar]
- Yao, Y.; Cheng-Shuai, J.; Na, S.; Li, W.Q.; Niu, Y.; Han, H.Q.; Miao, Z.H.; Zhao, X.X.; Zhao, J.; Li, J. Tamaractam, a new bioactive lactam from Tamarix ramosissima, induces apoptosis in rheumatoid arthritis fibroblast-like synoviocytes. Molecules 2017, 22, 96. [Google Scholar] [CrossRef]
- Liao, L.; Li, D.; Wu, C. Effects of Huangqi Chongteng drink on the growth and proliferation of synovial cells in rheumatoid arthritis rats. J. Hunan Univ. Chin. Med. 2018, 38, 31–35. [Google Scholar]
- Cao, W.; Zhang, J.; Wang, G.; Lu, J.; Wang, T.; Chen, X. Reducing-autophagy derived mitochondrial dysfunction during resveratrol promotes FLS cell apoptosis. Anat. Rec. 2018, 301, 1179–1188. [Google Scholar]
- Wang, D. A Preliminary Research on the Effect of Wogonin for Synovial Fibroblasts on Rheumatoid Arthritis. Master Thesis, Guangzhou University of Chinese Medicine, Guangzhou, China, 2013. [Google Scholar]
- Wang, H.; Wang, Q.P.; Tang, X.Z.; Ding, H.T.; Zhang, M.L.; Zhang, D.; Guo, Y.R.; Liu, S.T.; Fan, X.Y.; Xu, J.; et al. Experimental research on the influence of total glucosides of paeonia on the proliferation, apoptosis and cell cycle of fibroblast-like synovial cells. Tianjing J. Trad. Chin. Med. 2017, 34, 841–844. [Google Scholar]
- Wang, H.; Wang, Q.F.; Yang, L.L.; Zhang, D.; Wang, W.L.; Ding, H.B.; Guo, Y.R.; Du, W.Y.; Xu, M.K.; Gan, W. Experimental research on the influence of cinnamic aldehyde on the proliferation, apoptosis and cell cycle of fibroblast-like synovial cells. Tianjing J. Trad. Chin. Med. 2017, 34, 54–58. [Google Scholar]
- Fang, W.L.; Quan, F.; Nie, Y.K. Effect of water-soluble paclitaxel on inhibition of synovial fibroblasts cells. J. Harbin Med. Univ. 2018, 52, 314–316. [Google Scholar]
- Hou, Y.N.; Guo, L.H. Role of synovial fibroblasts in the pathogenesis of rheumatoid arthritis. Chin. J. Cell Biol. 2009, 31, 157–162. [Google Scholar]
- Liu, P.; Yang, H.; Long, F.; Hao, H.P.; Xu, X.; Liu, Y.; Shi, X.W.; Zhang, D.D.; Zheng, H.C.; Wen, Q.Y.; et al. Bioactive equivalence of combinatorial components identified in screening of an herbal medicine. Pharm. Res. 2014, 31, 1788–1800. [Google Scholar] [CrossRef]
- Peng, W.; Shen, H.; Lin, B.; Han, P.; Li, C.H.; Zhang, Q.Y.; Ye, B.Z.; Rahman, K.; Xin, H.L.; Qin, L.P.; et al. Docking study and antiosteoporosis effects of a dibenzylbutane lignan isolated from Litsea cubeba targeting Cathepsin K and MEK1. Med. Chem. Res. 2018, 27, 2062–2070. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potential Pathways | Detail Mechanisms | Extracts/Monomers (dose/concentration) | Cells/Tissues | Related Genes/Proteins | Reference |
|---|---|---|---|---|---|
| Death receptors mediated apoptotic pathway | Up-regulating Fas and FasL; Down-regulating Bcl-2 | XFC (1.8 g/kg, p.o.) | Synovia tissues in rats | Fas, FasL, Bcl-2 | [40] |
| Up-regulating Fas | ERSM (0.4 mg/mL) | RA-FLS | Fas | [41] | |
| Up-regulating Caspase-8, Fas and FasL | FSBT (9.5–38 g/kg, p.o.) | Synovia tissues in rats | Caspase-8, Fas, FasL | [42] | |
| Up-regulating Caspase-8 and FLIP | Resveratrol (50–400 μM) | rFLS (CIA) | Caspase-8, FLIP | [44] | |
| Increasing Fas mRNA | Propyl gallate (64 μg/mL) | RA-FLS | Fas | [48,49] | |
| Up-regulating Caspase -3, -8, -9, and FasL | Daphnetin (40 μg/mL) | rFLS (CIA) | FasL, TNF, Bid, Bax, Bcl-2, Cyt-C, Caspase-3, -8, and -9 | [50,51] | |
| Mitochondrial apoptotic pathway | Up-regulating Bax and Caspase-3; Down-regulating Bcl-2 | SWPD (15–25 mg/mL) | RA-FLS | Bcl-2, Bax, Caspase-3 | [53] |
| Down-regulating Bcl-2; Activating Caspase-3; Increasing Cyt-C release | FSNC (0.33–1.32 g/kg) | rFLS (CIA) | Bcl-2, Caspase-3, Cyt-C | [54] | |
| Up-regulating Bax and Caspase-3; Down-regulating Bcl-2 | DHJS medicated serum, (0.75, 1.5 and 3 g/kg) | rFLS (AIA) | Bax, Bcl-2, and Caspase-3 | [55] | |
| Up-regulating p53, Bax and Caspase-3; Downreglulating Bcl-2 | SDP (5–15 mg/mL) | rFLS (AIA) | p53, Bax, Caspase-3, Bcl-2 | [56] | |
| Up-regulating p53, Bak, Cyt-C, Bax, Caspase-3, -9; Down-regulating Bcl-2 and Bcl-xL; Activating Caspase-3, -9 | PHSE (25–100 μg/mL) | MH7A | p53, Bax, Bak, Bcl-2, Bcl-xL, Cyt-C, Caspase-3, -9 | [57] | |
| Inhibiting proinflammatory cytokines; Up-regulating Caspase-3, -9 and Bax; Down-regulating Bcl-2, | GSZD (0.4–1.6 mg/mL) | MH7A, stimulated by TNF-α (20 ng/mL) | Caspase-3, -9, Bax, Bcl-2 | [16] | |
| Up-regulating Bax; Down-regulating Bcl-2; Decreasing MCMP (Δψm); Activating Caspase -3 | Scopoletin (250–1000 μM) | rFLS (AIA), stimulated by LPS (5 μg/mL) | MCMP, Caspase-3, Bax, Bcl | [58] | |
| Up-regulating Caspase-3 and Bax; Down-regulating Bcl-2; Activating Caspase-3 | DMHP (10–250 μM) | rFLS (AIA) | Caspase-3, Bax, Bcl-2 | [59] | |
| Increasing Caspase-3, -9, Bax and PARP; Decreasing Bcl-2, Bcl-xL and MCMP (Δψm) | Berberine (5–75 μM) | RA-FLS | Caspase-3, -9, Bax, PARP, Bcl-2, Bcl-xL | [60] | |
| Up-regulating Bax and Caspase-3; Down-regulating Bcl-2 | TAHP (50, 250 μM) | rFLS (AIA) | Bcl-2, Bax, Caspase-3 | [61] | |
| Up-regulating Bax and Cyt-C; Down-regulating Bcl-2; Activating Caspase -3 | AGPD (10–30 μM) | RA-FLS | Bax, Bcl-2, Caspase-3, Cyt-C | [62] | |
| Increasing Caspase-3 activity; Up-regulating Bax and p53; Down-regulating Bcl-2 | Gallic acid (0.1, 1 μM) | RA-FLS | Caspase-3,Bax, p53, Bcl-2 | [63] | |
| Increasing Caspase-3, -9, PARP, Fas and Bax; Decreasing Bcl-2 and MCMP (Δψm) | Celastrol (1, 2, 5 μM) | RA-FLS | Caspase-3, -9, PARP, Fas, Bax, Bcl-2 | [64] | |
| Increasing Caspase-3, -9, Cyto C; Decreasing Bcl-2 | Quercetin (100–300 μM) | RA-FLS | Caspase-3, -9, Cyto C, Bcl-2 | [65] | |
| Up-regulating Bax; Down-regulating Bcl-2; Activating Caspase -3 and PARP Increasing Cyt-C release; Decreasing MCMP (Δψm) | Bufalin (10–40 nM) | RA-FLS, stimulated by IL-1β (1 ng/mL) | MCMP, Bax, Bcl-2, PARP, Caspase-3, Cyt-C | [66] | |
| Up-regulating Bax, cytosolic Cyto C, Apaf-1, Caspase-3, -9; Down-regulating Bcl-2 | Tanshinone IIA (2.5–20 μM) | RA-FLS | Bax, Cyto C, Apaf-1, Caspase-3, -9, Bcl-2 | [67] | |
| Up-regulating Caspase-3,-8, -9, Bax, Bid and Cyt-C; Down-regulating Bcl-2 | Daphnetin (40 μg/mL) | rFLS (CIA) | FasL, TNF, Cyt-C, Bid, Bax, Bcl-2, Caspase-3, -8, -9 | [68] | |
| Decreasing MCMP (Δψm); Increasing Cyt-C release; Up-regulating Caspase-3 -9 and PARP | Oridonin (5–40 μM) | RA-FLS, stimulated by IL-1β (1 μg/mL) | MCMP, Caspase-3,-9, PARP, Cyt- C | [69] | |
| Down-regulating MCMP (Δψm) | Resveratrol (50–400 μM) | rFLS (AIA), stimulated by H2O2 (5 μM) | MCMP | [70,71] | |
| Up-regulating Bax and Caspase-3; Down-regulating Bcl-2 | Pristimerin (0.75–3 μM) | rFLS (AIA) | Bax, Caspase-3, Bcl-2 | [73] | |
| Down-regulating Bcl-2/Bax | Liquirtin (0.345–34.5 μM) | RA-FLS | Bcl-2, Bax | [74] | |
| Increasing ROS; Up-regulating Bad, Caspase-3, PARP; Down-regulating Bcl-2 | Cryptotanshinone (5 μM) | MH7A cells and RA-FLS cells | Bcl-2, Bad, Caspase-3, PARP | [75] | |
| NF-κB mediated apoptotic pathway | Down-regulating Bcl-2, MyD88 and TRAF-6 | Sinomenine (0.5–3.2 mM) | RA-FLS | MyD88, TRAF-6, Bcl-2 | [81,82,83] |
| Up-regulating Bax and IκBα; Down-regulating Bcl-2, p-IKK and p-IκBα | Scopoletin (250–1000 μM) | rFLS (AIA), stimulated by LPS (5 μg/mL) | Bax, IκBα, Bcl-2, p-IKK, p-IκBα | [57] | |
| Inhibiting phosphorylation of NF-κB and IκBα; Activating Caspase-3, -7 | Curcumin (12.5–80 μM) | RA-FLS, MH7A, stimulated by IL-1β (10 ng/mL) | NF-κB, IKBα, Caspase-3, -7 | [84,85] | |
| Inhibiting phosphorylation of IKK and IκBα; Down-regulating NF-κB p65 | Celastrol (0.25–2 μM) | RA-FLS | IKK, IκBα, NF-κBp65 | [86] | |
| Inhibiting phosphorylation of NF-κBp65 and IKKβ, IκB and MSK1; Down-regulating XIAP and Cyclin D1; Up-regulating GADD45a | XAN (8.7–34.7 μM) | RA-FLS | NF-κBp65, IKKβ, IκB, MSK1, XIAP, Cyclin D1, GADD45a | [87,88] | |
| Down-regulating NF-κB p65, IKKα and IKKβ | JJD medicated serum | RA-FLS | NF-κBp65, IKKα, IKKβ | [89,90] | |
| Decreasing NF-κB p65, phospho-NF-κB p65 and acetyl-NF-κB p65, as well as pro-inflammatory cytokines | Baicalin (10, 20, 30 μM) | RA-FLS | NF-κB p65 | [91] | |
| Inhibiting phosphorylation of NF-κBp65 | PLM (5–20 μM) | RA-FLS, stimulated by TNF-α (10 ng/mL) | NF-κBp65 | [92] | |
| Down-regulating p-NF-κBp65 and p-IκB | Kaempferitrin (5–20 μM) | MH7A | NF-κBp65, p-NF-κBp65, IκB, p-IκB, | [93] | |
| Increasing ROS, cleaved Caspase-9, Cleaved PARP; Decreasing NF-κB p65 | Hypericin (0.25–4 μM) | MH7A | Caspase-9, PARP, NF-κB p65 | [94] | |
| Down-regulating XIAP; Up-regulating Caspase 3; Inhibit phosphorylation of NF-κBp65, IκB and IKK | α-Mangostin (6–14 μg/mL) | RA-FLS, stimulated by TNF-α (10 ng/mL) | XIAP, Caspase 3, p65, IκB, IKK | [95] | |
| MAPK mediated apoptotic pathway | Up-regulating JNK, p38; Down-regulating ERK; Increasing DNA fragmentation | Diosgenin (40 μM) | RA-FLS, stimulated by IL-1β (1 ng/mL) | JNK, ERK, p38 | [107] |
| Up-regulating JNK, p38α; Down-regulating ERK; Activating Caspase -3, -8, -9 | Hecogenin and Tigogenin (10, 40 μM) | RA-FLS | JNK, ERK, p38α, Caspase-3,-8,-9 | [108] | |
| Up-regulating JNK, p38α; Down-regulating ERK1/2; Activating Caspase -3, - 7 and PARP-1 | Apigenin (25–100 μM) | MH7A | JNK, ERK, p38, Caspase-3, - 7, PARP-1 | [109] | |
| Inhibiting Ras-MAPK signaling | Triptolide (0.28–200 nM) | RA-FLS, stimulated by TNF-α (10 ng/mL) | Ras, p38, ERK, JNK | [100,112] | |
| Up-regulating Bax, ERK, P38 and p21; Down-regulating Bcl-2, JNK | XAN (10, 30 μg/mL) | MH7A, stimulated by TNF-α (10 ng/mL) | Bax, ERK, p38, Bcl-2, JNK | [110] | |
| Down-regulating JNK and p-JNK | Brucine (0.125–2 mg/mL) | RA-FLS, stimulated by TNF-α (10 ng/mL) | JNK, p-JNK | [113] | |
| Up-regulating p38; Activating Caspase-3, -9 | β-Elemene (10–200 μg/mL) | RA-FLS | p38, Caspase- 3, - 9 | [114] | |
| ERS mediated apoptotic pathway | Up-regulating CHOP, GRP94 and GRP78; Activating PARP | Hempseed oil (2.5%) | MH7A | CHOP, PARP | [119] |
| Up-regulating ATF4, CHOP and XBPI; Activating Caspase-3, -7 and PARP; Increasing phosphorylation of eIF2α, IRE1α and BiP | DK-59 (10 μM) | MH7A | ATF4, CHOP, XBPI, Caspase-3, -7, PARP, eIF2α, IRE1α, BiP | [120] | |
| Up-regulating ATF4 and CHOP; Activating Caspase -7 and PARP; Increasing phosphorylation of eIF2α | EJTE (37.5 μg/mL) | MH7A, stimulated by TNF-α (10 ng/mL) | ATF4, CHOP, Caspase- 7, PARP, IeIF2α | [121] | |
| Up-regulating Bax and CHOP; Down-regulating Bcl-2 | Resveratrol (50–400 μM) | rFLS (AIA), stimulated by H2O2 (5 μM) | CHOP, Bcl-2 | [122] | |
| PI3K/AKT mediated apoptotic pathway | Up-regulating Bax; Down-regulating Bcl-2; Inhibiting phosphorylation of PI3K and Akt | DTYD (100, 200 μg/mL) | RA-FLS | Bax, Bcl-2, PI3K, Akt | [128] |
| Up-regulating Bax; Down-regulating Bcl-2, PI3K and Akt | SWXB medicated serum (4.32, 8.64, 17.28 g/kg) | RA-FLS | Bax, Bcl-2, PI3K, Akt | [129] | |
| Inhibiting phosphorylation of PI3K and Akt | TSDNR (100 μg/L) | rFLS, stimulated by IL-1β (10 μg/L) | PI3K, Akt | [127] | |
| Down-regulating HIF-α, p-PI3K, p-Akt, Bax, Bcl-2 in synovial tissue of CIA rats | HGTZFC (0.315 g/kg) | Synovial tissue of CIA rats | HIF-α, p-PI3K, p-Akt, Bcl-2, Bax | [130] | |
| Up-regulating Bax; Down-regulating Bcl-2; Inhibiting phosphorylation of Akt | Genistein (50–200 μM) | rFLS (CIA) | Bax, Bcl-2, Akt | [131] | |
| Up-regulating lncRNA GAS5; Up-regulating cleaved Caspase-3, -9; Inhibiting PI3K/Akt signaling | Tanshinone IIA (40 µM) | RA-FLS | lncRNA GAS5, Caspase-3, -9, PI3K, Akt | [132] | |
| Decreasing Akt and miR-633 | Anacardic acid (5, 30 and 60 µM) | RA-FLS, stimulated by TNF-α (10 ng/mL) | Akt | [133] | |
| Inhibiting the phosphorylation of Akt; Increasing p21 | Juglone (30 µM) | RA-FLS | Akt, p21 | [134] | |
| Up-regulating cleaved Caspase-3, Bax; Down-regulating PI3K, Akt, mTOR | Diosgenin (10, 20, 40 μg/mL) | RA-FLS | Caspase-3, Bax, PI3K, Akt, mTOR | [135] | |
| Up-regulating Bax; Down-regulating Bcl-2, PI3K and Akt | Pectolinarin (10, 20 µM) | RA-FLS | Bax, Bcl-2, PI3K, Akt | [136] | |
| Other | Down-regulating Jak2 and STAT3; Inhibiting Jak2/STAT3 signaling | TAHP (50, 250 μM) | rFLS (AIA) | Jak2, STAT3, p-STAT3 | [60] |
| Up-regulating Bax, LC3A, ATR, Chk-1 and ATR; Down-regulating Bcl-2, FasR and Cyclin-B1; Increasing phosphorylation of Cdc-2, -25; Activating Caspase-3, -9 and PARP; Increasing DNA damage G2/M phase of stagnation | Celastrol (1–5 µM) | RA-FLS | Bax, LC3A, ATR, Chk-1, γ-H2AX, Bcl-2, FasR, Cyclin-b1, Cdc-25, Cdc-2, Caspase-3, -9, PARP | [138] | |
| Inducing pro-apoptosis effects on the RA-FLS | Wogonin (111, 0–200 μM) | RA-FLS | [143] | ||
| Up-regulating Bax and Caspase-3; Down-regulating Bcl-2; Inhibiting phosphorylation of JAK2, STAT-1, -3 | Matrine (0.75 mg/mL) | rFLS (CIA) | Bax, Bcl-2, Caspase-3, JAK2, STAT1, -3 | [137] | |
| Down-regulating VEGF and MMP-3 | 10-HCPT (1, 10 μg/mL) | RA-FLS | VEGF and MMP-3 | [139] | |
| Activating Caspase-3, -7; Increasing G1 cell cycle | Tamaractam; CFM; TFM (0.1, 1 µM) | RA-FLS | Caspase-3, -7 | [140] | |
| Inducing growth stagnation of synovial cells at G0/G1 stage | HCTD (6.25–100 µg/mL) | rFLS (CIA) | [141] | ||
| Up-regulating Bax and LC3A; Down-regulating Bcl-2, Atg5 and LC3B; Increasing ROS production and Ca2+ release | Resveratrol (40–320 µM) | RA-FLS, stimulated by H2O2 (5 μM) | Bax, LC3A, Bcl-2, Atg5, LC3B | [142] | |
| Inducing pro-apoptosis effects on the RA-FLS | TGP (5–50 μg/mL) | RA-FLS | [144] | ||
| Inhibiting pro-inflammatory cytokines; Down-regulating Bcl-2, Jak2, STAT-3, -5; Inhibiting Jak2/STAT3 signaling | GSZD (0.4–1.6 mg/mL) | MH7A, stimulated by TNF-α (20 ng/mL) | SOCS1, JAK2, STAT-3, -5 | [16] | |
| Inducing pro-apoptosis effects on the RA-FLS | CINA (1,10 μg/mL) | RA-FLS | [145] | ||
| Inducing pro-apoptosis effects on the RA-FLS | Paclitaxel (2,4,8 μM) | RA-FLS | [146] |
| Classification | Monomers | Apoptotic pathways | References |
|---|---|---|---|
| Alkaloids | Berberine | Mitochondrial dependent apoptosis | [60] |
| Sinomenine | NF-κB mediated apoptosis | [81,82,83] | |
| PLM | NF-κB mediated apoptosis | [92] | |
| Brucine | MAPK mediated apoptosis | [113] | |
| Matrine | Inhibiting JAK/STAT | [137] | |
| 10-HCPT | Down-regulating VEGF and MMP-3 | [139] | |
| Tamaractam CFM TFM | Caspase activation induced apoptosis and induction of cell arrest | [140] | |
| Flavonoids | DMHP | Mitochondrial dependent apoptosis | [59] |
| TAHP | Mitochondrial dependent apoptosis Inhibiting JAK/STAT | [61] | |
| Quercetin | Mitochondrial dependent apoptosis | [65] | |
| Liquiritin | Mitochondrial dependent apoptosis | [74] | |
| XAN | NF-κB mediated apoptosis MAPK mediated apoptosis | [87,88] [110] | |
| Baicalin | NF-κB mediated apoptosis | [91] | |
| Kaempferitrin | NF-κB mediated apoptosis | [93] | |
| α-mangostin | NF-κB Mediated apoptosis | [95] | |
| Apigenin | MAPK mediated apoptosis | [109] | |
| Genistein | PI3K-Akt mediated apoptosis | [131] | |
| Pectolinarin | PI3K-Akt mediated apoptosis | [136] | |
| Wogonin | Not mentioned | [143] | |
| Steroids | Bufalin | Mitochondrial dependent apoptosis NF-κB mediated apoptosis | [66] |
| Diosgenin | MAPK mediated apoptosis | [107,108] | |
| Hecogenin | |||
| Tigogenin | |||
| Phenylpropanoids | Daphnetin | Death receptors mediated Mitochondrial dependent apoptosis | [50,51] |
| Scopoletin | Mitochondrial dependent apoptosis NF-κB mediated apoptosis | [57,58] | |
| Curcumin | NF-κB mediated apoptosis | [84,85] | |
| CINA | Not mentioned | [145] | |
| Terpenoids | AGPD | Mitochondrial dependent apoptosis | [62] |
| Oridonin | Mitochondrial dependent apoptosis; | [69] | |
| Celastrol | Mitochondrial dependent apoptosis; NF-κB mediated apoptosis; Inducing DNA damage and cell cycle arrest | [64] [86] [138] | |
| Pristimerin | Mitochondrial dependent apoptosis | [73] | |
| Triptolide | MAPK mediated apoptosis | [100,112] | |
| β-Elemene | MAPK mediated apoptosis | [114] | |
| Paclitaxel | Not mentioned | [146] | |
| Quinones | Cryptotanshinone | Mitochondrial dependent apoptosis | [75] |
| Hypericin | NF-κB mediated apoptosis | [94] | |
| Tanshinone IIA | PI3K-Akt mediated apoptosis | [132] | |
| Others | Propyl gallate | Death receptors mediated apoptosis | [48,49] |
| Gallic acid | Mitochondrial dependent apoptosis | [63] | |
| Resveratrol | Mitochondrial dependent apoptosis ERS mediated apoptosis | [70,71] [122] | |
| DK-59 | ERS mediated apoptosis | [120] | |
| Anacardic acid | PI3K-Akt mediated apoptosis | [133] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Liu, J.; Zhang, M.; Wei, S.; Li, R.; Gao, Y.; Peng, W.; Wu, C. Apoptosis Induction of Fibroblast-Like Synoviocytes Is an Important Molecular-Mechanism for Herbal Medicine along with its Active Components in Treating Rheumatoid Arthritis. Biomolecules 2019, 9, 795. https://doi.org/10.3390/biom9120795
Zhang Q, Liu J, Zhang M, Wei S, Li R, Gao Y, Peng W, Wu C. Apoptosis Induction of Fibroblast-Like Synoviocytes Is an Important Molecular-Mechanism for Herbal Medicine along with its Active Components in Treating Rheumatoid Arthritis. Biomolecules. 2019; 9(12):795. https://doi.org/10.3390/biom9120795
Chicago/Turabian StyleZhang, Qing, Jia Liu, Mengmeng Zhang, Shujun Wei, Ruolan Li, Yongxiang Gao, Wei Peng, and Chunjie Wu. 2019. "Apoptosis Induction of Fibroblast-Like Synoviocytes Is an Important Molecular-Mechanism for Herbal Medicine along with its Active Components in Treating Rheumatoid Arthritis" Biomolecules 9, no. 12: 795. https://doi.org/10.3390/biom9120795
APA StyleZhang, Q., Liu, J., Zhang, M., Wei, S., Li, R., Gao, Y., Peng, W., & Wu, C. (2019). Apoptosis Induction of Fibroblast-Like Synoviocytes Is an Important Molecular-Mechanism for Herbal Medicine along with its Active Components in Treating Rheumatoid Arthritis. Biomolecules, 9(12), 795. https://doi.org/10.3390/biom9120795