The Role of Calcium in Inflammation-Associated Bone Resorption

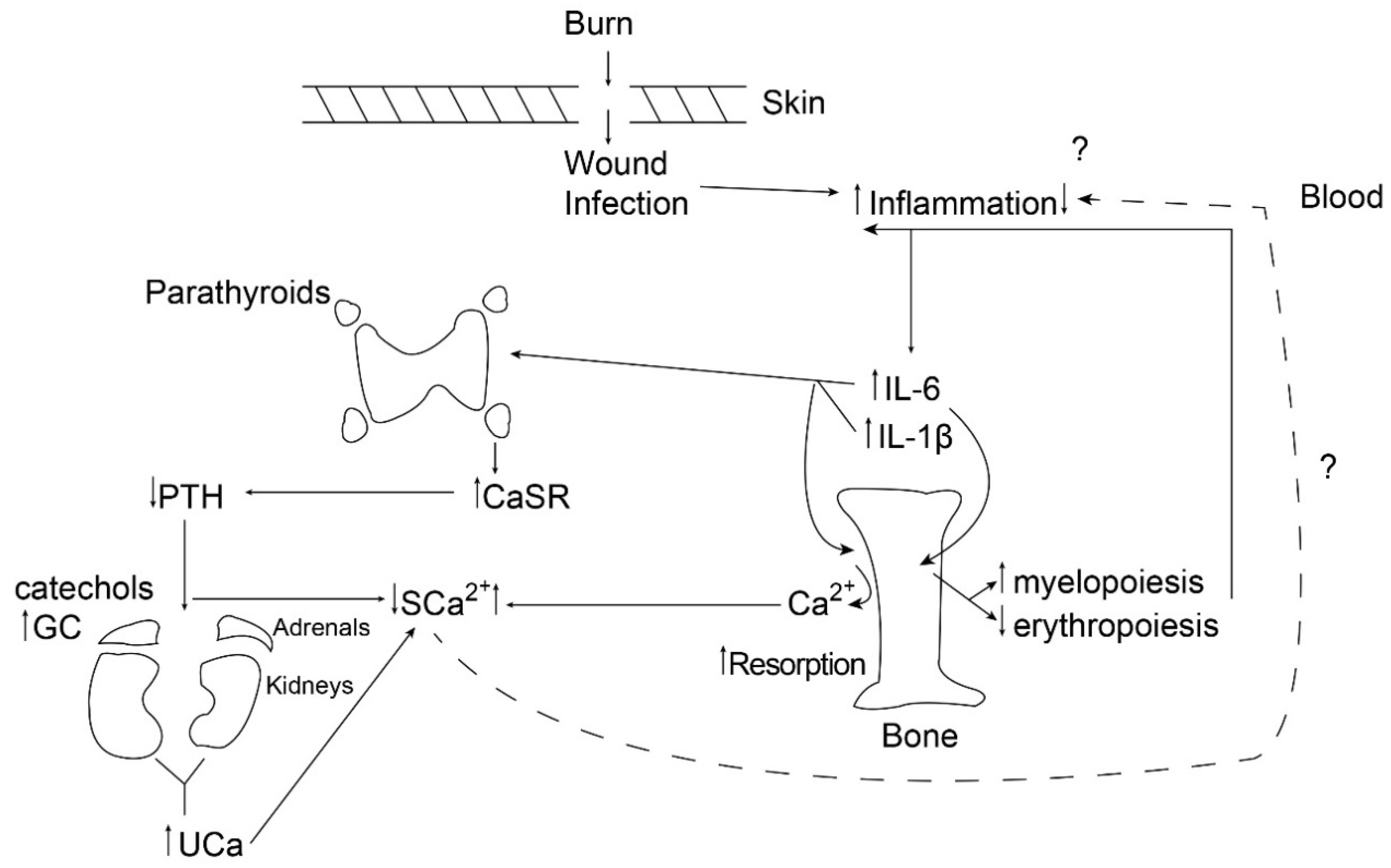
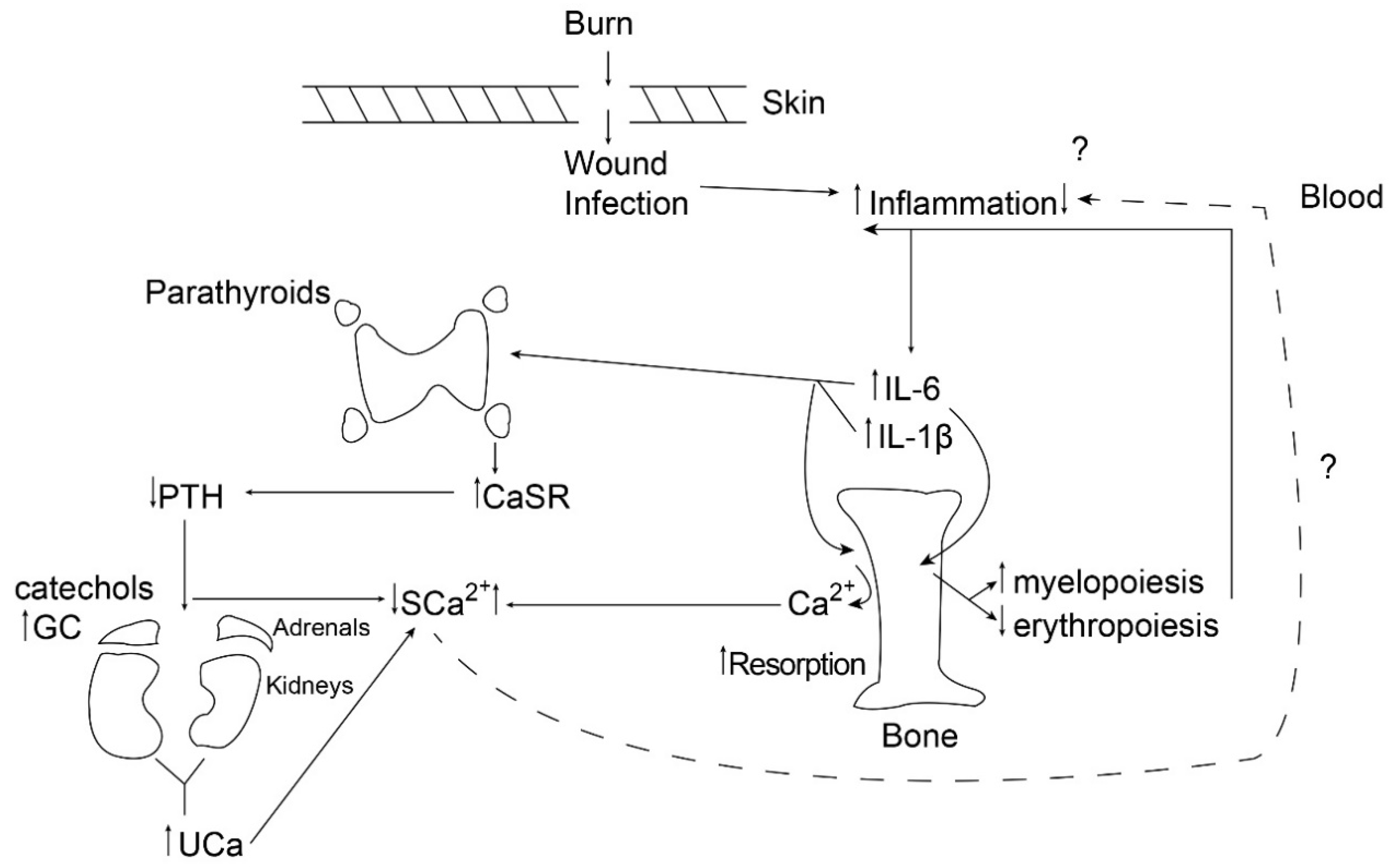{kind=link}
Abstract
:1. Introduction
2. Bone Resorption in the Setting of Burn Injury
3. Bisphosphonates and Calcium
4. Applicability to Other Inflammatory Conditions
5. Conclusions
Funding
Conflicts of Interest
References
- Griffin, L.M.; Thayu, M.; Baldassano, R.N.; DeBoer, M.D.; Zemel, B.S.; Denburg, M.R.; Denson, L.A.; Shults, J.; Herskovitz, R.; Long, J.; et al. Improvements in bone density and structure during anti-TNF therapy in pediatric Crohn’s disease. J. Clin. Endocrinol. Metab. 2015, 100, 2630–2639. [Google Scholar] [CrossRef] [PubMed]
- Ross, F.P. Osteoclast biology and bone resorption. In Primer on the Metabolic Bone Diseases and Disorders of Mineral Metabolism, 8th ed.; Rosen, C.J., Ed.; American Society for Bone and Mineral Research: New York, NY, USA; Wiley: New York, NY, USA, 2013; pp. 25–33. [Google Scholar]
- Van Niekerk, G.; Mitchell, M.; Engelbrecht, A.M. Bone resorption: Supporting immunometabolism. Biol. Lett. 2018, 14. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.; Sousse, L.E.; Irick, R.; Schryver, E.; Klein, G.L. Interactions of phosphate metabolism with severe injury, including burns. JBMR Plus 2017, 1, 59–65. [Google Scholar] [CrossRef]
- Klein, G.L.; Herndon, D.N.; Le, P.T.; Andersen, C.R.; Benjamin, D.; Rosen, C.J. The effect of burn injury on serum concentrations of sclerostin and FGF23. Burns 2015, 41, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H. Interaction of the endocrine system with inflammation: A function of energy and volume regulation. Arthritis Res. Ther. 2014, 16, 203. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.L.; Herndon, D.N.; Goodman, W.G.; Langman, C.B.; Phillips, W.A.; Dickson, I.R.; Eastell, R.; Naylor, K.E.; Maloney, N.A.; Desai, M.; et al. Histomorphometric and biochemical characterization of bone following acute severe burns in children. Bone 1995, 17, 455–460. [Google Scholar] [CrossRef]
- Song, J.; Saeman, M.R.; De Libero, J.; Wolf, S.E. Skeletal muscle loss is associated with TNF mediated insufficient skeletal myogenic activation after burn. Shock 2015, 44, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.K.; Rasmussen, A.K.; Butters, R.; Feldt-Rasmussen, U.; Bendtzen, K.; Diaz, R.; Brown, E.M.; Olgaard, K. Inhibition of PTH secretion by interleukin-1 beta in bovine parathyroid glands in vitro is associated with up-regulation of the calcium-sensing receptor mRNA. Biochem. Biophys. Res. Commun. 1997, 238, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Toribio, R.E.; Kohn, C.W.; Capen, C.C.; Rosol, T.J. Parathyroid hormone (PTH) secretion, PTH mRNA, and calcium-sensing receptor mRNA expression in equine parathyroid cells and effects of interleukin (IL)-1, IL-6 and tumor necrosis factor-alpha on equine parathyroid cell function. J. Mol. Endocrinol. 2003, 31, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Canaff, L.; Zhou, X.; Hendy, G.N. The pro-inflammatory cytokine, interleukin-6, up-regulates calcium sensing receptor gene transcription vis Stat1/3 and Sp 1/3. J. Biol. Chem. 2008, 283, 13586–13600. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M. Ca2+ sensing receptor. In Primer on the Metabolic Bone Diseases and Disorders of Mineral Metabolism, 8th ed.; Rosen, C.J., Ed.; American Society for Bone and Mineral Research: New York, NY, USA; Wiley: New York, NY, USA, 2013; pp. 224–234. [Google Scholar]
- Klein, G.L.; Nicolai, M.; Langman, C.B.; Curneo, B.F.; Sailer, D.E.; Herndon, D.N. Dysregulation of calcium homeostasis after severe burn injury in children: Possible role of magnesium depletion. J. Pediatr. 1997, 131, 246–251. [Google Scholar] [CrossRef]
- Murphey, E.D.; Chattopadhyay, N.; Bai, M.; Kifor, O.; Harper, D.; Traber, D.L.; Hawkins, H.K.; Brown, E.M.; Klein, G.L. Up-regulation of the parathyroid calcium-sensing receptor after burn injury in sheep: A potential contributory factor to post-burn hypocalcemia. Crit. Care Med. 2000, 28, 3885–3890. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.L.; Herndon, D.N.; Rutan, T.C.; Sherrard, D.J.; Coburn, J.W.; Langman, C.B.; Thomas, M.L.; Haddad, J.G., Jr.; Cooper, C.W.; Miller, N.L.; et al. Bone disease in burn patients. J. Bone Miner. Res. 1993, 8, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.L.; Benjamin, D.; Herndon, D.N. Calcemic response to burns differs between adults and children: A review of the literature. Osteoporos. Sarcopenia 2017, 3, 170–173. [Google Scholar] [CrossRef]
- Klein, G.L.; Castro, S.M.; Garofalo, R.P. The calcium-sensing receptor as a mediator of inflammation. Semin. Cell Dev. Biol. 2016, 49, 52–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossol, M.; Pierer, M.; Raulien, N.; Quandt, D.; Meusch, U.; Rothe, K.; Schubert, K.; Schöneberg, T.; Schaefer, M.; Krügel, U.; et al. Extracellular Ca2+ is a danger signal activating the NLRP3 inflammasome through G protein-coupled calcium sensing receptors. Nat. Commun. 2012, 3, 1329. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, M.G.; Pinto, R.; Kraft, R.; Nathens, A.B.; Finnerty, C.C.; Gamelli, R.L.; Gibran, N.S.; Klein, M.B.; Arnoldo, B.D.; Tompkins, R.G.; et al. Morbidity and survival probability in burn patients in modern burn care. Crit. Care Med. 2015, 43, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.L.; Wimalawansa, S.J.; Kulkarni, G.; Sherrard, D.J.; Sanford, A.P.; Herndon, D.N. The efficacy of acute administration of pamidronate on the conservation of bone mass following severe burn injury in children: A double blind, randomized, controlled study. Osteoporos. Int. 2005, 16, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Prkzora, R.; Herndon, D.N.; Sherrard, D.J.; Chinkes, D.L.; Klein, G.L. Pamidronate preserves bone mass for at least two years following acute administration for pediatric burn injury. Bone 2007, 41, 297–302. [Google Scholar] [CrossRef]
- Borsheim, E.; Herndon, D.N.; Hawkins, H.K.; Suman, O.E.; Cotter, M.; Klein, G.L. Pamidronate attenuates muscle loss following pediatric burn injury. J. Bone Miner. Res. 2014, 29, 1369–1372. [Google Scholar] [CrossRef] [PubMed]
- Tsoumpra, M.K.; Muniz, J.R.; Barnett, B.L.; Kwaasi, A.A.; Pilka, E.S.; Kavanagh, K.L.; Evdokimov, A.; Walter, R.L.; Von Delft, F.; Ebetino, F.H.; et al. The inhibition of human farnesyl pyrophosphate synthase by nitrogen-containing bisphosphonates. Elucidating the role of active site threonine 201 and tyrosine 204 residues using enzyme mutants. Bone 2015, 81, 478–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, G.L. The role of bone secreted factors in burn-induced muscle cachexia. Curr. Osteoporos. Rep. 2018, 16, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, R.E.; Lissina, A.; Green, A.E.; Slay, E.S.; Price, D.A.; Sewell, A.K. The bisphosphonate acute phase response: Rapid and copious production of pro-inflammatory cytokines by peripheral blood γδ T cells in response to aminobisphosphonates is inhibited by statins. Clin. Exp. Immunol. 2005, 139, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Finnerty, C.C.; Jeschke, M.G.; Herndon, D.N.; Gamelli, R.; Gibran, N.; Klein, M.; Silver, G.; Arnoldo, B.; Remick, D.; Tompkins, R.G.; et al. Temporal cytokine profiles in severely burned patients: A comparison of adults and children. Mol. Med. 2008, 14, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Zheng, X.F.; Qian, B.H.; Fang, H.; Wang, J.J.; Zhang, L.L.; Pang, Y.F.; Zhang, J.; Wei, X.Q.; Xia, Z.F.; et al. Plasma IL-37 is elevated in patients with rheumatoid arthritis: Its correlation with disease activity and Th1/Th2 and Th17 related cytokines. Dis. Mark. 2015, 2015, 795043. [Google Scholar] [CrossRef]
- Gomes, R.G.; Brito, C.A.A.; Martinelli, V.F.; Santos, R.N.D.; Gomes, F.O.D.S.; Peixoto, C.A.; Crispim, J.O.; Diniz, G.T.N.; Donadi, E.A.; Lucena-Silva, N. HLA-G is expressed in intestinal samples of ulcerative colitis and Crohn’s disease patients and HLA-G5 expression is differentially correlated with TNF and IL-10 cytokine expression. Hum. Immunol. 2018, 79, 477–484. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klein, G.L. The Role of Calcium in Inflammation-Associated Bone Resorption. Biomolecules 2018, 8, 69. https://doi.org/10.3390/biom8030069
Klein GL. The Role of Calcium in Inflammation-Associated Bone Resorption. Biomolecules. 2018; 8(3):69. https://doi.org/10.3390/biom8030069
Chicago/Turabian StyleKlein, Gordon L. 2018. "The Role of Calcium in Inflammation-Associated Bone Resorption" Biomolecules 8, no. 3: 69. https://doi.org/10.3390/biom8030069
APA StyleKlein, G. L. (2018). The Role of Calcium in Inflammation-Associated Bone Resorption. Biomolecules, 8(3), 69. https://doi.org/10.3390/biom8030069