
The Regulatory Role of LncRNAs in Modulating Autophagy and Drug Resistance in Non-Small-Cell Lung Cancer: Focus on Targeted Therapeutic Approaches

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
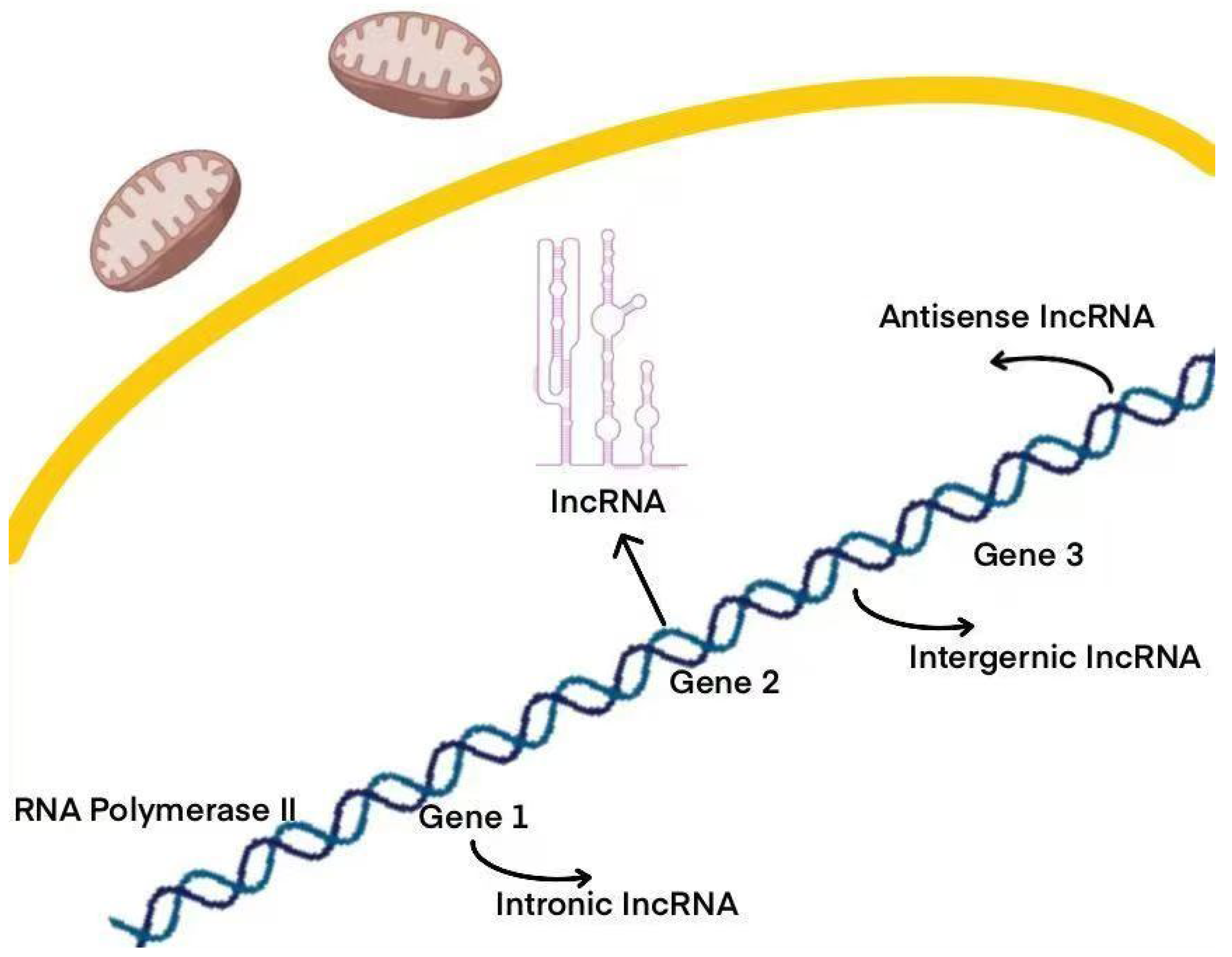
2. Overview of LncRNA
3. LncRNAs in Non-Small-Cell Lung Cancer
3.1. LINC01140
3.2. MALAT1
3.3. HOTAIR
3.4. H19
3.5. NEAT1
3.6. PVT1
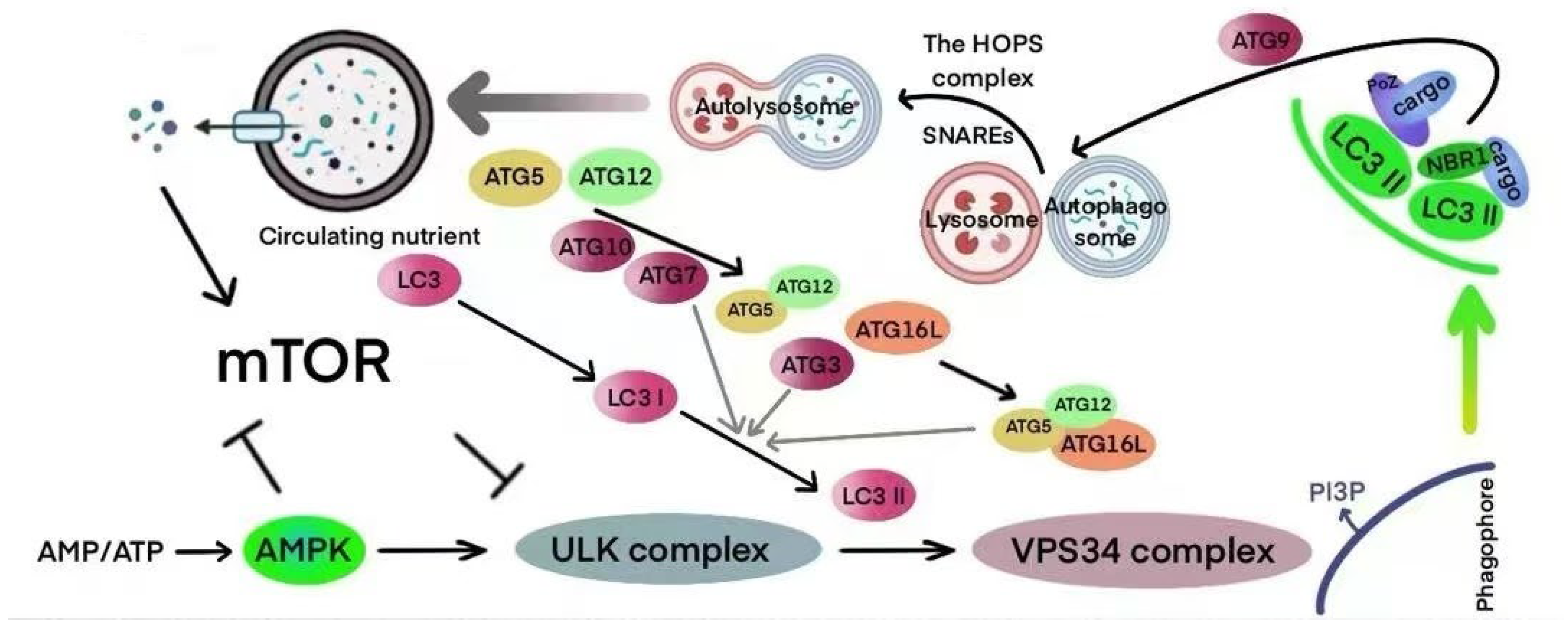
4. Autophagy
4.1. Types of Autophagy
4.2. Mechanisms of Autophagy
5. The Dual Regulatory Effect of Autophagy in Non-Small-Cell Lung Cancer
5.1. The Tumor-Inhibiting Role of Autophagy in Non-Small-Cell Lung Cancer
5.2. The Tumor-Promoting Effects of Autophagy in Non-Small-Cell Lung Cancer
6. lncRNAs and Autophagy in Non-Small-Cell Lung Cancer
6.1. The Suppressive Role of Autophagy-Related lncRNAs in Autophagy in Non-Small-Cell Lung Cancer
6.1.1. Autophagy-Related lncRNAs Inhibit Autophagy and the Pathogenesis of Non-Small-Cell Lung Cancer
6.1.2. Autophagy-Related lncRNAs Inhibit Autophagy and Promote the Pathogenesis of Non-Small-Cell Lung Cancer
6.2. The Promoting Effect of Autophagy-Related lncRNAs on Autophagy in Non-Small-Cell Lung Cancer
6.2.1. Autophagy-Related lncRNAs Activate Autophagy and Suppress the Pathogenesis of Non-Small-Cell Lung Cancer
6.2.2. Autophagy-Related lncRNAs Activate Autophagy and Promote the Pathogenesis of Non-Small-Cell Lung Cancer
7. Clinical Significance of lncRNAs in Autophagy and Resistance in Non-Small-Cell Lung Cancer
8. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Abbreviation | Full Term |
| NSCLC | Non-Small-Cell Lung Cancer |
| lncRNA | Long Non-Coding RNA |
| ceRNA | Competing Endogenous RNA |
| EMT | Epithelial–Mesenchymal Transition |
| FDA | Food and Drug Administration |
| EGFR-TKI | Epidermal Growth Factor Receptor-Tyrosine Kinase Inhibitor |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| mTOR | Mechanistic Target of Rapamycin |
| AMPK | AMP-Activated Protein Kinase |
| ULK1 | Unc-51 Like Autophagy Activating Kinase 1 |
| LC3 | Microtubule-Associated Protein 1 Light Chain 3 |
| ATG | Autophagy-Related Gene |
| CMA | Chaperone-Mediated Autophagy |
| ROS | Reactive Oxygen Species |
| PSA | Prostate-Specific Antigen |
| HOTAIR | HOX Transcript Antisense RNA |
| MALAT1 | Metastasis-Associated Lung Adenocarcinoma Transcript 1 |
| PVT1 | Plasmacytoma Variant Translocation 1 |
| XIST | X-Inactive Specific Transcript |
| BLACAT1 | Bladder Cancer-Associated Transcript 1 |
| LUCAT1 | Lung Cancer-Associated Transcript 1 |
| SNHG7 | Small Nucleolar RNA Host Gene 7 |
| TCGA | The Cancer Genome Atlas |
| ORR | Objective Response Rate |
| IC50 | Half Maximal Inhibitory Concentration |
References
- Suster, D.I.; Mino-Kenudson, M. Molecular Pathology of Primary Non-small Cell Lung Cancer. Arch. Med. Res. 2020, 51, 784–798. [Google Scholar] [CrossRef]
- Pelosi, G.; Barbareschi, M.; Cavazza, A.; Graziano, P.; Rossi, G.; Papotti, M. Large cell carcinoma of the lung: A tumor in search of an author. A clinically oriented critical reappraisal. Lung Cancer 2015, 87, 226–231. [Google Scholar] [CrossRef]
- Garg, P.; Singhal, S.; Kulkarni, P.; Horne, D.; Malhotra, J.; Salgia, R.; Singhal, S.S. Advances in Non-Small Cell Lung Cancer: Current Insights and Future Directions. J. Clin. Med. 2024, 13, 4189. [Google Scholar] [CrossRef]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Leiter, A.; Veluswamy, R.R.; Wisnivesky, J.P. The global burden of lung cancer: Current status and future trends. Nat. Rev. Clin. Oncol. 2023, 20, 624–639. [Google Scholar] [CrossRef]
- Sosa Iglesias, V.; Giuranno, L.; Dubois, L.J.; Theys, J.; Vooijs, M. Drug Resistance in Non-Small Cell Lung Cancer: A Potential for NOTCH Targeting? Front. Oncol. 2018, 8, 267. [Google Scholar] [CrossRef]
- Min, H.Y.; Lee, H.Y. Mechanisms of resistance to chemotherapy in non-small cell lung cancer. Arch. Pharmacal Res. 2021, 44, 146–164. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, S.; Han, Y.; Shi, H.; Yuan, J.; Cui, W. LncRNA SH3PXD2A-AS1 facilitates cisplatin resistance in non-small cell lung cancer by regulating FOXM1 succinylation. BMC Cancer 2024, 24, 848. [Google Scholar] [CrossRef]
- Vitto, V.A.M.; Bianchin, S.; Zolondick, A.A.; Pellielo, G.; Rimessi, A.; Chianese, D.; Yang, H.; Carbone, M.; Pinton, P.; Giorgi, C.; et al. Molecular Mechanisms of Autophagy in Cancer Development, Progression, and Therapy. Biomedicines 2022, 10, 1596. [Google Scholar] [CrossRef]
- Debnath, J.; Gammoh, N.; Ryan, K.M. Autophagy and autophagy-related pathways in cancer. Nat. Rev. Mol. Cell Biol. 2023, 24, 560–575. [Google Scholar] [CrossRef]
- Li, X.; He, S.; Ma, B. Autophagy and autophagy-related proteins in cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef]
- Han, J.; Goldstein, L.A.; Hou, W.; Chatterjee, S.; Burns, T.F.; Rabinowich, H. HSP90 inhibition targets autophagy and induces a CASP9-dependent resistance mechanism in NSCLC. Autophagy 2018, 14, 958–971. [Google Scholar] [CrossRef]
- Guo, W.; Du, K.; Luo, S.; Hu, D. Recent Advances of Autophagy in Non-Small Cell Lung Cancer: From Basic Mechanisms to Clinical Application. Front. Oncol. 2022, 12, 861959. [Google Scholar] [CrossRef]
- Yu, M.; Li, H.; Wu, Y.; Liu, P.; Xu, Q.; Zhang, Y. Autophagy-associated ncRNAs in lung cancer: From drug resistance to therapeutic targets. Int. J. Biol. Macromol. 2025, 319, 145477. [Google Scholar] [CrossRef]
- Ratti, M.; Lampis, A.; Ghidini, M.; Salati, M.; Mirchev, M.B.; Valeri, N.; Hahne, J.C. MicroRNAs (miRNAs) and Long Non-Coding RNAs (lncRNAs) as New Tools for Cancer Therapy: First Steps from Bench to Bedside. Target. Oncol. 2020, 15, 261–278. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Lin, W.; Zhou, Q.; Wang, C.Q.; Zhu, L.; Bi, C.; Zhang, S.; Wang, X.; Jin, H. LncRNAs regulate metabolism in cancer. Int. J. Biol. Sci. 2020, 16, 1194–1206. [Google Scholar] [CrossRef]
- Fu, J.; Yu, L.; Yan, H.; Tang, S.; Wang, Z.; Dai, T.; Chen, H.; Zhang, S.; Hu, H.; Liu, T.; et al. LncRNAs in non-small cell lung cancer: Novel diagnostic and prognostic biomarkers. Front. Mol. Biosci. 2023, 10, 1297198. [Google Scholar] [CrossRef]
- Al-Tobasei, R.; Paneru, B.; Salem, M. Genome-Wide Discovery of Long Non-Coding RNAs in Rainbow Trout. PLoS ONE 2016, 11, e0148940. [Google Scholar] [CrossRef]
- Yi, Q.; Feng, J.; Lan, W.; Shi, H.; Sun, W.; Sun, W. CircRNA and lncRNA-encoded peptide in diseases, an update review. Mol. Cancer 2024, 23, 214. [Google Scholar] [CrossRef]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef]
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long non-coding RNAs: Mechanism of action and functional utility. Non-Coding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- St Laurent, G.; Wahlestedt, C.; Kapranov, P. The Landscape of long noncoding RNA classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef]
- Lin, Y.H. Crosstalk of lncRNA and Cellular Metabolism and Their Regulatory Mechanism in Cancer. Int. J. Mol. Sci. 2020, 21, 2947. [Google Scholar] [CrossRef]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef]
- Herman, A.B.; Tsitsipatis, D.; Gorospe, M. Integrated lncRNA function upon genomic and epigenomic regulation. Mol. Cell 2022, 82, 2252–2266. [Google Scholar] [CrossRef]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Lennox, K.A.; Behlke, M.A. Cellular localization of long non-coding RNAs affects silencing by RNAi more than by antisense oligonucleotides. Nucleic Acids Res. 2016, 44, 863–877. [Google Scholar] [CrossRef]
- Carlevaro-Fita, J.; Polidori, T.; Das, M.; Navarro, C.; Zoller, T.I.; Johnson, R. Ancient exapted transposable elements promote nuclear enrichment of human long noncoding RNAs. Genome Res. 2019, 29, 208–222. [Google Scholar] [CrossRef]
- Quinodoz, S.; Guttman, M. Long noncoding RNAs: An emerging link between gene regulation and nuclear organization. Trends Cell Biol. 2014, 24, 651–663. [Google Scholar] [CrossRef]
- Naganuma, T.; Hirose, T. Paraspeckle formation during the biogenesis of long non-coding RNAs. RNA Biol. 2013, 10, 456–461. [Google Scholar] [CrossRef]
- Song, Z.; Lin, J.; Li, Z.; Huang, C. The nuclear functions of long noncoding RNAs come into focus. Non-Coding RNA Res. 2021, 6, 70–79. [Google Scholar] [CrossRef]
- Hussein, M.A.; Valinezhad, K.; Adel, E.; Munirathinam, G. MALAT-1 Is a Key Regulator of Epithelial-Mesenchymal Transition in Cancer: A Potential Therapeutic Target for Metastasis. Cancers 2024, 16, 234. [Google Scholar] [CrossRef]
- Naseer, Q.A.; Malik, A.; Zhang, F.; Chen, S. Exploring the enigma: History, present, and future of long non-coding RNAs in cancer. Discov. Oncol. 2024, 15, 214. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Al-Imam, M.J.; Hussein, U.A.; Sead, F.F.; Faqri, A.M.A.; Mekkey, S.M.; Khazel, A.J.; Almashhadani, H.A. The interactions between DNA methylation machinery and long non-coding RNAs in tumor progression and drug resistance. DNA Repair 2023, 128, 103526. [Google Scholar] [CrossRef]
- Wang, C.J.; Shi, S.B.; Tian, J.; Xu, J.; Niu, Z.X. lncRNA MALAT1, HOTTIP and PVT1 as predictors for predicting the efficacy of GEM based chemotherapy in first-line treatment of pancreatic cancer patients. Oncotarget 2017, 8, 95108–95115. [Google Scholar] [CrossRef]
- Badowski, C.; He, B.; Garmire, L.X. Blood-derived lncRNAs as biomarkers for cancer diagnosis: The Good, the Bad and the Beauty. NPJ Precis. Oncol. 2022, 6, 40. [Google Scholar] [CrossRef]
- Nagarajah, S.; Saleh, Q.W.; Rasmussen, M.; Tepel, M. Long non-coding RNA MGAT3 in kidney transplant recipients with immunoglobulin A nephropathy. J. Nephrol. 2024, 37, 1133–1135. [Google Scholar] [CrossRef]
- Galbiati, S.; Bettiga, A.; Colciago, G.; Senti, C.; Trevisani, F.; Villa, G.; Marzinotto, I.; Ghidini, M.; Passalacqua, R.; Montorsi, F.; et al. The long noncoding RNA SUMO1P3 as urinary biomarker for monitoring bladder cancer progression. Front. Oncol. 2024, 14, 1325157. [Google Scholar] [CrossRef]
- Beylerli, O.; Gareev, I.; Sufianov, A.; Ilyasova, T.; Guang, Y. Long noncoding RNAs as promising biomarkers in cancer. Non-Coding RNA Res. 2022, 7, 66–70. [Google Scholar] [CrossRef]
- Qiao, Y.; Tian, X.; Li, S.; Niu, H. Identification and experimental validation of a sialylation-related long noncoding RNA signature for prognosis of bladder cancer. BMC Urol. 2024, 24, 222. [Google Scholar] [CrossRef]
- Zhang, Y. LncRNA-encoded peptides in cancer. J. Hematol. Oncol. 2024, 17, 66. [Google Scholar] [CrossRef]
- Nemeth, K.; Bayraktar, R.; Ferracin, M.; Calin, G.A. Non-coding RNAs in disease: From mechanisms to therapeutics. Nat. Rev. Genet. 2024, 25, 211–232. [Google Scholar] [CrossRef]
- Rodríguez-Malavé, N.I.; Rao, D.S. Long noncoding RNAs in hematopoietic malignancies. Brief. Funct. Genom. 2016, 15, 227–238. [Google Scholar] [CrossRef]
- Tao, Y.; Liu, J.; Qiu, W.; Li, Y. LncRNA MANCR is downregulated in non-small cell lung cancer and predicts poor survival. Discov. Oncol. 2025, 16, 40. [Google Scholar] [CrossRef]
- Miao, X.; Xi, W.; Bao, Y. LncRNA RP11-58O9.2 predicts poor prognosis and promotes progression of non-small cell lung cancer. J. Int. Med. Res. 2023, 51, 3000605231206295. [Google Scholar] [CrossRef]
- Liang, H.; Peng, J. LncRNA HOTAIR promotes proliferation, invasion and migration in NSCLC cells via the CCL22 signaling pathway. PLoS ONE 2022, 17, e0263997. [Google Scholar] [CrossRef]
- Chi, X.; Feng, L.; Wang, L.; Yu, S.; Wei, M.; Zhang, Q.; Liu, X.; Shao, S. Downregulation of lncRNA MNX1-AS1 promotes the ferroptosis and apoptosis of non-small cell lung cancer. Int. J. Med. Sci. 2025, 22, 1052–1063. [Google Scholar] [CrossRef]
- Fang, F.; Zhao, M.; Meng, J.; He, J.; Yang, C.; Wang, C.; Wang, J.; Xie, S.; Jin, X.; Shi, W. Upregulation of TTYH3 by lncRNA LUCAT1 through interacting with ALYREF facilitates the metastasis in non-small cell lung cancer. Cancer Biol. Ther. 2025, 26, 2464966. [Google Scholar] [CrossRef]
- Pan, H.; Yu, T.; Sun, L.; Chai, W.; Liu, X.; Yan, M. LncRNA FENDRR-mediated tumor suppression and tumor-immune microenvironment changes in non-small cell lung cancer. Transl. Cancer Res. 2020, 9, 3946–3959. [Google Scholar] [CrossRef]
- Xiong, Z.; Han, Z.; Pan, W.; Zhu, X.; Liu, C. Correlation between chromatin epigenetic-related lncRNA signature (CELncSig) and prognosis, immune microenvironment, and immunotherapy in non-small cell lung cancer. PLoS ONE 2023, 18, e0286122. [Google Scholar] [CrossRef]
- Wang, M.; Fu, Y.; Zhong, C.; Gacche, R.N.; Wu, P.J.H. Long non-coding RNA and Evolving drug resistance in lung cancer. Heliyon 2023, 9, e22591. [Google Scholar] [CrossRef]
- Xia, R.; Geng, G.; Yu, X.; Xu, Z.; Guo, J.; Liu, H.; Li, N.; Li, Z.; Li, Y.; Dai, X.; et al. LINC01140 promotes the progression and tumor immune escape in lung cancer by sponging multiple microRNAs. J. Immunother. Cancer 2021, 9, e002746. [Google Scholar] [CrossRef]
- Chatterjee, M.; Nag, S.; Gupta, S.; Mukherjee, T.; Shankar, P.; Parashar, D.; Maitra, A.; Das, K. MicroRNAs in lung cancer: Their role in tumor progression, biomarkers, diagnostic, prognostic, and therapeutic relevance. Discov. Oncol. 2025, 16, 293. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Zhang, L.; Chen, Y.; Ha, M. LINC01140 inhibits nonsmall cell lung cancer progression and cisplatin resistance through the miR-4742-5p/TACC1 axis. J. Biochem. Mol. Toxicol. 2022, 36, e23048. [Google Scholar] [CrossRef]
- He, L.; Zhao, X.; He, L. LINC01140 Alleviates the Oxidized Low-Density Lipoprotein-Induced Inflammatory Response in Macrophages via Suppressing miR-23b. Inflammation 2020, 43, 66–73. [Google Scholar] [CrossRef]
- Li, X. LINC01140 Targeting miR-452-5p/RGS2 Pathway to Attenuate Breast Cancer Tumorigenesis. Dis. Markers 2022, 2022, 2434938. [Google Scholar] [CrossRef]
- Gencel-Augusto, J.; Wu, W.; Bivona, T.G. Long Non-Coding RNAs as Emerging Targets in Lung Cancer. Cancers 2023, 15, 3135. [Google Scholar] [CrossRef]
- Li, S.; Ma, F.; Jiang, K.; Shan, H.; Shi, M.; Chen, B. Long non-coding RNA metastasis-associated lung adenocarcinoma transcript 1 promotes lung adenocarcinoma by directly interacting with specificity protein 1. Cancer Sci. 2018, 109, 1346–1356. [Google Scholar] [CrossRef]
- Tetik Vardarlı, A.; Ozgur, S.; Goksel, T.; Korba, K.; Karakus, H.S.; Asık, A.; Pelit, L.; Gunduz, C. Conversion of specific lncRNAs to biomarkers in exhaled breath condensate samples of patients with advanced stage non-small-cell lung cancer. Front. Genet. 2023, 14, 1200262. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, L.; Li, H.; Sun, T.; Wen, X.; Li, X.; Meng, Y.; Li, Y.; Liu, M.; Liu, S.; et al. Nuclear-Encoded lncRNA MALAT1 Epigenetically Controls Metabolic Reprogramming in HCC Cells through the Mitophagy Pathway. Mol. Ther. Nucleic Acids 2021, 23, 264–276. [Google Scholar] [CrossRef]
- Zhou, Q.; Liu, L.; Zhou, J.; Chen, Y.; Xie, D.; Yao, Y.; Cui, D. Novel Insights Into MALAT1 Function as a MicroRNA Sponge in NSCLC. Front. Oncol. 2021, 11, 758653. [Google Scholar] [CrossRef]
- Chen, W.; Tan, X.; Yang, Q.; Fang, Z.; Xu, Y. MALAT1 enhances gemcitabine resistance in non-small cell lung cancer cells by directly affecting miR-27a-5p/PBOV1 axis. Cell Signal 2022, 94, 110326. [Google Scholar] [CrossRef]
- Roh, J.; Kim, B.; Im, M.; Jang, W.; Chae, Y.; Kang, J.; Youn, B.; Kim, W. MALAT1-regulated gene expression profiling in lung cancer cell lines. BMC Cancer 2023, 23, 818. [Google Scholar] [CrossRef]
- Tiansheng, G.; Junming, H.; Xiaoyun, W.; Peixi, C.; Shaoshan, D.; Qianping, C. lncRNA Metastasis-Associated Lung Adenocarcinoma Transcript 1 Promotes Proliferation and Invasion of Non-Small Cell Lung Cancer Cells via Down-Regulating miR-202 Expression. Cell J. 2020, 22, 375–385. [Google Scholar] [CrossRef]
- Nazari, M.; Babakhanzadeh, E.; Mollazadeh, A.; Ahmadzade, M.; Mohammadi Soleimani, E.; Hajimaqsoudi, E. HOTAIR in cancer: Diagnostic, prognostic, and therapeutic perspectives. Cancer Cell Int. 2024, 24, 415. [Google Scholar] [CrossRef]
- Yao, X.; Wang, T.; Sun, M.Y.; Yuming, Y.; Guixin, D.; Liu, J. Diagnostic value of lncRNA HOTAIR as a biomarker for detecting and staging of non-small cell lung cancer. Biomarkers 2022, 27, 526–533. [Google Scholar] [CrossRef]
- Li, H.S.; Xu, Y. Inhibition of EZH2 via the STAT3/HOTAIR signalling axis contributes to cell cycle arrest and apoptosis induced by polyphyllin I in human non-small cell lung cancer cells. Steroids 2020, 164, 108729. [Google Scholar] [CrossRef]
- Yu, Y.; Ren, K. Five long non-coding RNAs establish a prognostic nomogram and construct a competing endogenous RNA network in the progression of non-small cell lung cancer. BMC Cancer 2021, 21, 457. [Google Scholar] [CrossRef]
- Chen, D.; Li, Y.; Wang, Y.; Xu, J. LncRNA HOTAIRM1 knockdown inhibits cell glycolysis metabolism and tumor progression by miR-498/ABCE1 axis in non-small cell lung cancer. Genes Genom. 2021, 43, 183–194. [Google Scholar] [CrossRef]
- Zhu, C.; Wang, X.; Wang, Y.; Wang, K. Functions and underlying mechanisms of lncRNA HOTAIR in cancer chemotherapy resistance. Cell Death Discov. 2022, 8, 383. [Google Scholar] [CrossRef]
- Zhang, X.; Luo, M.; Zhang, J.; Guo, B.; Singh, S.; Lin, X.; Xiong, H.; Ju, S.; Wang, L.; Zhou, Y.; et al. The role of lncRNA H19 in tumorigenesis and drug resistance of human Cancers. Front. Genet. 2022, 13, 1005522. [Google Scholar] [CrossRef]
- Zhang, R.; Zheng, Y.; Zhu, Q.; Gu, X.; Xiang, B.; Gu, X.; Xie, T.; Sui, X. β-Elemene Reverses Gefitinib Resistance in NSCLC Cells by Inhibiting lncRNA H19-Mediated Autophagy. Pharmaceuticals 2024, 17, 626. [Google Scholar] [CrossRef]
- Lei, Y.; Guo, W.; Chen, B.; Chen, L.; Gong, J.; Li, W. Tumor-released lncRNA H19 promotes gefitinib resistance via packaging into exosomes in non-small cell lung cancer. Oncol. Rep. 2018, 40, 3438–3446. [Google Scholar] [CrossRef]
- Gu, B.; Yang, M.; Shi, L.; Yuan, G.; Xie, H.; Ni, B. Progranulin modulates the progression of non-small cell lung cancer through lncRNA H19. Am. J. Transl. Res. 2023, 15, 4887–4901. [Google Scholar]
- Xia, Y.; Pei, T.; Zhao, J.; Wang, Z.; Shen, Y.; Yang, Y.; Liang, J. Long noncoding RNA H19: Functions and mechanisms in regulating programmed cell death in cancer. Cell Death Discov. 2024, 10, 76. [Google Scholar] [CrossRef]
- Liu, L.; Liu, L.; Lu, S. lncRNA H19 promotes viability and epithelial-mesenchymal transition of lung adenocarcinoma cells by targeting miR-29b-3p and modifying STAT3. Int. J. Oncol. 2019, 54, 929–941. [Google Scholar] [CrossRef]
- Tang, F.; Tian, L.H.; Zhu, X.H.; Yang, S.; Zeng, H.; Yang, Y.Y. METTL3-mediated m6A modification enhances lncRNA H19 stability to promote endothelial cell inflammation and pyroptosis to aggravate atherosclerosis. Faseb J. 2024, 38, e70090. [Google Scholar] [CrossRef]
- Qi, L.; Yin, Y.; Sun, M. m6A-mediated lncRNA NEAT1 plays an oncogenic role in non-small cell lung cancer by upregulating the HMGA1 expression through binding miR-361-3p. Genes Genom. 2023, 45, 1537–1547. [Google Scholar] [CrossRef]
- Hussain, M.S.; Afzal, O.; Gupta, G.; Goyal, A.; Almalki, W.H.; Kazmi, I.; Alzarea, S.I.; Alfawaz Altamimi, A.S.; Kukreti, N.; Chakraborty, A.; et al. Unraveling NEAT1’s complex role in lung cancer biology: A comprehensive review. EXCLI J. 2024, 23, 34–52. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Dong, M.; Wu, D. Long non-coding RNA NEAT1 regulates E2F3 expression by competitively binding to miR-377 in non-small cell lung cancer. Oncol. Lett. 2017, 14, 4983–4988. [Google Scholar] [CrossRef]
- Sun, C.; Li, S.; Zhang, F.; Xi, Y.; Wang, L.; Bi, Y.; Li, D. Long non-coding RNA NEAT1 promotes non-small cell lung cancer progression through regulation of miR-377-3p-E2F3 pathway. Oncotarget 2016, 7, 51784–51814. [Google Scholar] [CrossRef]
- Zhao, L.; Bi, M.; Zhang, H.; Shi, M. Downregulation of NEAT1 Suppresses Cell Proliferation, Migration, and Invasion in NSCLC Via Sponging miR-153-3p. Cancer Biother. Radiopharm. 2020, 35, 362–370. [Google Scholar] [CrossRef]
- Zhao, M.M.; Ge, L.Y.; Yang, L.F.; Zheng, H.X.; Chen, G.; Wu, L.Z.; Shi, S.M.; Wang, N.; Hang, Y.P. LncRNA NEAT1/miR-204/NUAK1 Axis is a Potential Therapeutic Target for Non-Small Cell Lung Cancer. Cancer Manag. Res. 2020, 12, 13357–13368. [Google Scholar] [CrossRef]
- Wei, C.M.; Zhao, X.F.; Qiu, H.B.; Ming, Z.; Liu, K.; Yan, J. The long non-coding RNA PVT1/miR-145-5p/ITGB8 axis regulates cell proliferation, apoptosis, migration and invasion in non-small cell lung cancer cells. Neoplasma 2020, 67, 802–812. [Google Scholar] [CrossRef]
- Zeng, S.H.G.; Xie, J.H.; Zeng, Q.Y.; Dai, S.H.H.; Wang, Y.; Wan, X.M.; Liu, J.C.H. lncRNA PVT1 Promotes Metastasis of Non-Small Cell Lung Cancer Through EZH2-Mediated Activation of Hippo/NOTCH1 Signaling Pathways. Cell J. 2021, 23, 21–31. [Google Scholar] [CrossRef]
- Li, M.Y.; Tang, X.H.; Fu, Y.; Wang, T.J.; Zhu, J.M. Regulatory Mechanisms and Clinical Applications of the Long Non-coding RNA PVT1 in Cancer Treatment. Front. Oncol. 2019, 9, 787. [Google Scholar] [CrossRef]
- Qin, S.; Zhao, Y.; Lim, G.; Lin, H.; Zhang, X.; Zhang, X. Circular RNA PVT1 acts as a competing endogenous RNA for miR-497 in promoting non-small cell lung cancer progression. Biomed. Pharmacother. 2019, 111, 244–250. [Google Scholar] [CrossRef]
- Ryter, S.W.; Cloonan, S.M.; Choi, A.M. Autophagy: A critical regulator of cellular metabolism and homeostasis. Mol. Cells 2013, 36, 7–16. [Google Scholar] [CrossRef]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef]
- White, E. The role for autophagy in cancer. J. Clin. Investig. 2015, 125, 42–46. [Google Scholar] [CrossRef]
- Rakesh, R.; PriyaDharshini, L.C.; Sakthivel, K.M.; Rasmi, R.R. Role and regulation of autophagy in cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166400. [Google Scholar] [CrossRef]
- Liu, G.; Pei, F.; Yang, F.; Li, L.; Amin, A.D.; Liu, S.; Buchan, J.R.; Cho, W.C. Role of Autophagy and Apoptosis in Non-Small-Cell Lung Cancer. Int. J. Mol. Sci. 2017, 18, 367. [Google Scholar] [CrossRef]
- Chen, S.; Saeed, A.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G.G.; Rao, L.; Duo, Y. Macrophages in immunoregulation and therapeutics. Signal Transduct. Target. Ther. 2023, 8, 207. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abdalla, F.C.; Abeliovich, H.; Abraham, R.T.; Acevedo-Arozena, A.; Adeli, K.; Agholme, L.; Agnello, M.; Agostinis, P.; Aguirre-Ghiso, J.A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy 2012, 8, 445–544. [Google Scholar] [CrossRef]
- Amaravadi, R.; Kimmelman, A.C.; White, E. Recent insights into the function of autophagy in cancer. Genes Dev. 2016, 30, 1913–1930. [Google Scholar] [CrossRef]
- Parzych, K.R.; Klionsky, D.J. An overview of autophagy: Morphology, mechanism, and regulation. Antioxid. Redox Signal 2014, 20, 460–473. [Google Scholar] [CrossRef]
- Yorimitsu, T.; Klionsky, D.J. Autophagy: Molecular machinery for self-eating. Cell Death Differ. 2005, 12 (Suppl. S2), 1542–1552. [Google Scholar] [CrossRef]
- Ravikumar, B.; Sarkar, S.; Davies, J.E.; Futter, M.; Garcia-Arencibia, M.; Green-Thompson, Z.W.; Jimenez-Sanchez, M.; Korolchuk, V.I.; Lichtenberg, M.; Luo, S.; et al. Regulation of mammalian autophagy in physiology and pathophysiology. Physiol. Rev. 2010, 90, 1383–1435. [Google Scholar] [CrossRef]
- Huang, J.; Wang, J. Selective protein degradation through chaperone-mediated autophagy: Implications for cellular homeostasis and disease (Review). Mol. Med. Rep. 2025, 31, 13. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. The coming of age of chaperone-mediated autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 365–381. [Google Scholar] [CrossRef]
- Yu, X.; Ye, X.; Lin, H.; Feng, N.; Gao, S.; Zhang, X.; Wang, Y.; Yu, H.; Deng, X.; Qian, B. Knockdown of long non-coding RNA LCPAT1 inhibits autophagy in lung cancer. Cancer Biol. Med. 2018, 15, 228–238. [Google Scholar]
- Mizushima, N.; Levine, B. Autophagy in mammalian development and differentiation. Nat. Cell Biol. 2010, 12, 823–830. [Google Scholar] [CrossRef]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef]
- Cai, Q.; Wang, S.; Jin, L.; Weng, M.; Zhou, D.; Wang, J.; Tang, Z.; Quan, Z.J.M.c. Long non-coding RNA GBCDRlnc1 induces chemoresistance of gallbladder cancer cells by activating autophagy. Mol. Cancer 2019, 18, 82. [Google Scholar] [CrossRef]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef]
- Jahangiri, L.; Ishola, T.; Pucci, P.; Trigg, R.M.; Pereira, J.; Williams, J.A.; Cavanagh, M.L.; Gkoutos, G.V.; Tsaprouni, L.; Turner, S.D. The role of autophagy and lncRNAs in the maintenance of cancer stem cells. Cancers 2021, 13, 1239. [Google Scholar] [CrossRef]
- Pustovalova, M.; Alhaddad, L.; Blokhina, T.; Smetanina, N.; Chigasova, A.; Chuprov-Netochin, R.; Eremin, P.; Gilmutdinova, I.; Osipov, A.N.; Leonov, S. The CD44high subpopulation of multifraction irradiation-surviving NSCLC cells exhibits partial EMT-program activation and DNA damage response depending on their p53 status. Int. J. Mol. Sci. 2021, 22, 2369. [Google Scholar] [CrossRef]
- Li, N.; Gao, W.J.; Liu, N.S. LncRNA BCAR4 promotes proliferation, invasion and metastasis of non-small cell lung cancer cells by affecting epithelial-mesenchymal transition. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2075–2086. [Google Scholar]
- Huang, L.; Liu, X.; Chen, Q.; Yang, J.; Zhang, D.; Zhao, Y.; Xu, L.; Li, Z.; Liu, X.; Shao, S.; et al. TGF-β-induced lncRNA TBUR1 promotes EMT and metastasis in lung adenocarcinoma via hnRNPC-mediated GRB2 mRNA stabilization. Cancer Lett. 2024, 600, 217153. [Google Scholar] [CrossRef]
- Qin, P.; Li, Q.; Zu, Q.; Dong, R.; Qi, Y. Natural products targeting autophagy and apoptosis in NSCLC: A novel therapeutic strategy. Front. Oncol. 2024, 14, 1379698. [Google Scholar] [CrossRef]
- Liu, M.; Fan, Y.; Li, D.; Han, B.; Meng, Y.; Chen, F.; Liu, T.; Song, Z.; Han, Y.; Huang, L. Ferroptosis inducer erastin sensitizes NSCLC cells to celastrol through activation of the ROS–mitochondrial fission–mitophagy axis. Mol. Oncol. 2021, 15, 2084–2105. [Google Scholar] [CrossRef]
- Ariosa, A.R.; Lahiri, V.; Lei, Y.; Yang, Y.; Yin, Z.; Zhang, Z.; Klionsky, D.J. A perspective on the role of autophagy in cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166262. [Google Scholar] [CrossRef]
- Ma, Q.; Chen, K.; Xiao, H. Rapamycin combined with osimertinib alleviated non-small cell lung cancer by regulating the PARP, Akt/mTOR, and MAPK/ERK signaling pathways. Front. Mol. Biosci. 2025, 12, 1548810. [Google Scholar] [CrossRef]
- Ávalos, Y.; Canales, J.; Bravo-Sagua, R.; Criollo, A.; Lavandero, S.; Quest, A.F. Tumor suppression and promotion by autophagy. Biomed. Res. Int. 2014, 2014, 603980. [Google Scholar] [CrossRef]
- Stefanou, D.T.; Kouvela, M.; Stellas, D.; Voutetakis, K.; Papadodima, O.; Syrigos, K.; Souliotis, V.L. Oxidative stress and deregulated DNA damage response network in lung cancer patients. Biomedicines 2022, 10, 1248. [Google Scholar] [CrossRef]
- Carretero-Fernández, M.; Cabrera-Serrano, A.J.; Sánchez-Maldonado, J.M.; Ruiz-Durán, L.; Jiménez-Romera, F.; García-Verdejo, F.J.; González-Olmedo, C.; Cardús, A.; Díaz-Beltrán, L.; Gutiérrez-Bautista, J.F.; et al. Autophagy and Oxidative Stress in Solid Tumors: Mechanisms and Therapeutic Opportunities. Crit. Rev. Oncol. Hematol. 2025, 212, 104820. [Google Scholar] [CrossRef]
- Kumar, P.; Choudhary, A.; Kinger, S.; Jagtap, Y.A.; Prajapati, V.K.; Chitkara, D.; Chinnathambi, S.; Verma, R.K.; Mishra, A. Autophagy as a potential therapeutic target in regulating improper cellular proliferation. Front. Pharmacol. 2025, 16, 1579183. [Google Scholar] [CrossRef]
- Chen, X.; He, Q.; Zeng, S.; Xu, Z. Upregulation of nuclear division cycle 80 contributes to therapeutic resistance via the promotion of autophagy-related protein-7-dependent autophagy in lung cancer. Front. Pharmacol. 2022, 13, 985601. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, P.-J.; Zhang, D.; Chen, Z.-H.; Cao, X.-C.; Yu, Y.; Ge, J. An autophagy-associated lncRNAs model for predicting the survival in non-small cell lung cancer patients. Front. Genet. 2022, 13, 919857. [Google Scholar] [CrossRef]
- Liu, T.-T.; Li, R.; Liu, X.; Zhou, X.-J.; Huo, C.; Li, J.-P.; Qu, Y.-Q. LncRNA XIST acts as a MicroRNA-520 sponge to regulate the Cisplatin resistance in NSCLC cells by mediating BAX through CeRNA network. Int. J. Med. Sci. 2021, 18, 419. [Google Scholar] [CrossRef]
- Kumar, A.; Girisa, S.; Alqahtani, M.S.; Abbas, M.; Hegde, M.; Sethi, G.; Kunnumakkara, A.B. Targeting Autophagy Using Long Non-Coding RNAs (LncRNAs): New Landscapes in the Arena of Cancer Therapeutics. Cells 2023, 12, 810. [Google Scholar] [CrossRef]
- Wang, Y.; Salai, A.; Luo, D.; Lv, H.; Gao, S.; Kamili, A.; Aishanjiang, D.; Liu, Y. Construction of a prognostic model for autophagy-related LncRNAs in lung adenocarcinoma. Medicine 2025, 104, e42122. [Google Scholar] [CrossRef]
- Yang, D.; Ma, X.; Song, P. A prognostic model of non small cell lung cancer based on TCGA and ImmPort databases. Sci. Rep. 2022, 12, 437. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, Y.; Zhang, B.; Sheng, Z.; Sun, N.; Yuan, B.; Wu, X. Identification of lncRNA, miRNA and mRNA expression profiles and ceRNA Networks in small cell lung cancer. BMC Genom. 2023, 24, 217. [Google Scholar] [CrossRef]
- Braga, E.A.; Fridman, M.V.; Burdennyy, A.M.; Loginov, V.I.; Dmitriev, A.A.; Pronina, I.V.; Morozov, S.G. Various LncRNA Mechanisms in Gene Regulation Involving miRNAs or RNA-Binding Proteins in Non-Small-Cell Lung Cancer: Main Signaling Pathways and Networks. Int. J. Mol. Sci. 2023, 24, 13617. [Google Scholar] [CrossRef]
- Ding, D.; Zhang, J.; Luo, Z.; Wu, H.; Lin, Z.; Liang, W.; Xue, X. Analysis of the lncRNA–miRNA–mRNA network reveals a potential regulatory mechanism of EGFR-TKI resistance in NSCLC. Front. Genet. 2022, 13, 851391. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, J.; Wang, C.; Zhang, Q.; Zeng, A.; Song, L. Autophagy-related lncRNAs in tumor progression and drug resistance: A double-edged sword. Genes Dis. 2024, 11, 367–381. [Google Scholar] [CrossRef]
- Jiao, J.; Zhao, Y.; Li, Q.; Jin, S.; Liu, Z. LncRNAs in tumor metabolic reprogramming and tumor microenvironment remodeling. Front. Immunol. 2024, 15, 1467151. [Google Scholar] [CrossRef]
- Li, C.; Liu, J.-H.; Su, J.; Lin, W.-J.; Zhao, J.-Q.; Zhang, Z.-H.; Wu, Q. LncRNA XIST knockdown alleviates LPS-induced acute lung injury by inactivation of XIST/miR-132-3p/MAPK14 pathway: XIST promotes ALI via miR-132-3p/MAPK14 axis. Mol. Cell. Biochem. 2021, 476, 4217–4229. [Google Scholar] [CrossRef]
- Huang, F.X.; Chen, H.J.; Zheng, F.X.; Gao, Z.Y.; Sun, P.F.; Peng, Q.; Liu, Y.; Deng, X.; Huang, Y.H.; Zhao, C.; et al. LncRNA BLACAT1 is involved in chemoresistance of non-small cell lung cancer cells by regulating autophagy. Int. J. Oncol. 2019, 54, 339–347. [Google Scholar] [CrossRef]
- Borzi, C.; Ganzinelli, M.; Caiola, E.; Colombo, M.; Centonze, G.; Boeri, M.; Signorelli, D.; Caleca, L.; Rulli, E.; Busico, A. LKB1 Down-Modulation by miR-17 Identifies Patients With NSCLC Having Worse Prognosis Eligible for Energy-Stress–Based Treatments. J. Thorac. Oncol. 2021, 16, 1298–1311. [Google Scholar] [CrossRef]
- Li, J.; Gan, J.; Shi, S.; Huang, J.; Yang, Y. The potential of targeting autophagy-related non-coding RNAs in the treatment of lung cancer. Front. Pharmacol. 2025, 16, 1551258. [Google Scholar] [CrossRef]
- Luong, T.V.; Cao, M.T.T.; Nguyen, N.V.D.; Dang, H.N.N.; Nguyen, T.T. Roles of autophagy and long non-coding RNAs in gastric cancer. World J. Gastroenterol. 2025, 31, 101124. [Google Scholar] [CrossRef]
- Xu, L.; Huang, X.; Lou, Y.; Xie, W.; Zhao, H. Regulation of apoptosis, autophagy and ferroptosis by non-coding RNAs in metastatic non-small cell lung cancer. Exp. Ther. Med. 2022, 23, 352. [Google Scholar] [CrossRef]
- Zheng, T.; Li, D.; He, Z.; Feng, S.; Zhao, S. Long noncoding RNA NBAT1 inhibits autophagy via suppression of ATG7 in non-small cell lung cancer. Am. J. Cancer Res. 2018, 8, 1801. [Google Scholar]
- Sun, W.; Zu, Y.; Fu, X.; Deng, Y.J.O.r. Knockdown of lncRNA-XIST enhances the chemosensitivity of NSCLC cells via suppression of autophagy. Oncol. Rep. 2017, 38, 3347–3354. [Google Scholar] [CrossRef]
- Giordo, R.; Ahmadi, F.A.M.; Husaini, N.A.; Al-Nuaimi, N.; Ahmad, S.M.S.; Pintus, G.; Zayed, H. microRNA 21 and long non-coding RNAs interplays underlie cancer pathophysiology: A narrative review. Non-Coding RNA Res. 2024, 9, 831–852. [Google Scholar] [CrossRef]
- Rama Ballesteros, A.R.; Quiñonero Muñoz, F.J.; Mesas Hernández, C.; Melguizo Alonso, C.; Prados Salazar, J.C. Synthetic Circular miR-21 Sponge as Tool for Lung Cancer Treatment. Int. J. Mol. Sci. 2022, 23, 2963. [Google Scholar] [CrossRef]
- Li, Y.; Liang, Z.; He, H.; Huang, X.; Mo, Z.; Tan, J.; Guo, W.; Zhao, Z.; Wei, S. The lncRNA HOTAIR regulates autophagy and affects lipopolysaccharide-induced acute lung injury through the miR-17-5p/ATG2/ATG7/ATG16 axis. J. Cell Mol. Med. 2021, 25, 8062–8073. [Google Scholar] [CrossRef]
- Zheng, Y.; Jia, H.; Wang, P.; Liu, L.; Chen, Z.; Xing, X.; Wang, J.; Tan, X.; Wang, C. Silencing TRAIP suppresses cell proliferation and migration/invasion of triple negative breast cancer via RB-E2F signaling and EMT. Cancer Gene Ther. 2023, 30, 74–84. [Google Scholar] [CrossRef]
- Shen, W.; Hong, X.; Jin, C.; Xi, Y. LncRNA PSMG3AS1 promotes proliferation of non-small cell lung cancer cells by sponging miR-613 to upregulate SphK1. Cell Cycle 2021, 20, 829–838. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, X.; Du, S. Long non-coding RNA ACTA2-AS1 inhibits the cisplatin resistance of non-small cell lung cancer cells through inhibiting autophagy by suppressing TSC2. Cell Cycle 2022, 21, 368–378. [Google Scholar] [CrossRef]
- Vessoni, A.T.; Filippi-Chiela, E.C.; Menck, C.F.; Lenz, G. Autophagy and genomic integrity. Cell Death Differ. 2013, 20, 1444–1454. [Google Scholar] [CrossRef]
- Gadhave, D.G.; Sugandhi, V.V.; Jha, S.K.; Nangare, S.N.; Gupta, G.; Singh, S.K.; Dua, K.; Cho, H.; Hansbro, P.M.; Paudel, K.R. Neurodegenerative disorders: Mechanisms of degeneration and therapeutic approaches with their clinical relevance. Ageing Res. Rev. 2024, 99, 102357. [Google Scholar] [CrossRef]
- Chen, Z.H.; Cao, J.F.; Zhou, J.S.; Liu, H.; Che, L.Q.; Mizumura, K.; Li, W.; Choi, A.M.; Shen, H.H. Interaction of caveolin-1 with ATG12-ATG5 system suppresses autophagy in lung epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 306, L1016–L1025. [Google Scholar] [CrossRef]
- Nakano, Y.; Isobe, K.; Yoshizawa, T.; Urabe, N.; Homma, S.; Kishi, K. Upregulation of long non-coding RNA LINC00460 in EGFR-mutant lung cancer indicates a poor prognosis in patients treated with osimertinib. Oncol. Lett. 2023, 26, 380. [Google Scholar] [CrossRef]
- Jin, K.T.; Lu, Z.B.; Lv, J.Q.; Zhang, J.G. The role of long non-coding RNAs in mediating chemoresistance by modulating autophagy in cancer. RNA Biol. 2020, 17, 1727–1740. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, X.; Guo, Y.; Zhao, H.; Jiang, P.; Yu, Q.Q. The regulatory role of lncRNA in tumor drug resistance: Refracting light through a narrow aperture. Oncol. Res. 2025, 33, 837–849. [Google Scholar] [CrossRef]
- Dong, Y.; He, Y.; Geng, Y.; Wei, M.; Zhou, X.; Lian, J.; Hallajzadeh, J. Autophagy-related lncRNAs and exosomal lncRNAs in colorectal cancer: Focusing on lncRNA-targeted strategies. Cancer Cell Int. 2024, 24, 328. [Google Scholar] [CrossRef]
- Lei, X.; Zheng, Y.; Su, W. RNA-binding proteins and autophagy in lung cancer: Mechanistic insights and therapeutic perspectives. Discov. Oncol. 2025, 16, 599. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Tang, J.M.; Yang, M.Q.; Yang, Z.H.; Xia, J.Z. The role of LncRNA-mediated autophagy in cancer progression. Front. Cell Dev. Biol. 2024, 12, 1348894. [Google Scholar] [CrossRef]
- She, K.; He, S.; Lu, X.; Yu, S.; Li, M.; Xiong, W.; Zhou, M. LncRNA SNHG7 promotes non-small cell lung cancer progression and cisplatin resistance by inducing autophagic activity. J. Thorac. Dis. 2023, 15, 155–167. [Google Scholar] [CrossRef]
- Sun, Y.; Jin, S.D.; Zhu, Q.; Han, L.; Feng, J.; Lu, X.Y.; Wang, W.; Wang, F.; Guo, R.H. Long non-coding RNA LUCAT1 is associated with poor prognosis in human non-small lung cancer and regulates cell proliferation via epigenetically repressing p21 and p57 expression. Oncotarget 2017, 8, 28297–28311. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, X.; Chen, Y.; Huang, Y.; He, J.; Luo, H. miR-186-5p targeting SIX1 inhibits cisplatin resistance in non-small-cell lung cancer cells (NSCLCs). Neoplasma 2019, 67, 147–157. [Google Scholar] [CrossRef]
- Xi, S.; Ming, D.J.; Zhang, J.H.; Guo, M.M.; Wang, S.Y.; Cai, Y.; Liu, M.Y.; Wang, D.Q.; Zhang, Y.J.; Li, Y.; et al. Downregulation of N6-methyladenosine-modified LINC00641 promotes EMT, but provides a ferroptotic vulnerability in lung cancer. Cell Death Dis. 2023, 14, 359. [Google Scholar] [CrossRef]
- Mu, D.; Han, B.; Huang, H.; Zheng, Y.; Zhang, J.; Shi, Y. Unraveling the advances of non-coding RNAs on the tumor microenvironment: Innovative strategies for cancer therapies. J. Transl. Med. 2025, 23, 614. [Google Scholar] [CrossRef]
- Du, Y.; Zhu, S.; Liu, X.; Sun, Y.; Cui, T.; Liu, J.; Zhang, W.; Shao, S. LncRNA HOTAIR regulates the expression of MRP1 gene through the mir-6807-5p/Egr1 axis to affect the multidrug resistance of lung cancer cells. Gene 2025, 940, 149216. [Google Scholar] [CrossRef]
- You, H.; Wang, L.; Meng, H.; Li, J.; Fang, G. Autophagy: Shedding Light on the Mechanisms and Multifaceted Roles in Cancers. Biomolecules 2025, 15, 915. [Google Scholar] [CrossRef]
- Chen, B.; Dragomir, M.P.; Yang, C.; Li, Q.; Horst, D.; Calin, G.A. Targeting non-coding RNAs to overcome cancer therapy resistance. Signal Transduct. Target. Ther. 2022, 7, 121. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, X.; Yin, C.; Liu, Z.; Yu, H. Identification of circRNA-Based Biomarkers and ceRNA Mechanism in Non-Small Cell Lung Cancer. Cell Biochem. Biophys. 2025. Online ahead of print. [Google Scholar] [CrossRef]
- Ju, Z.S.; Sun, B.; Bao, D.; Zhang, X.F. Effect of lncRNA-BLACAT1 on drug resistance of non-small cell lung cancer cells in DDP chemotherapy by regulating cyclin D1 expression. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 9465–9472. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, X.; Chen, Z.; Gao, C.; Zhao, L.; Cui, Y. Silencing of lncRNA XIST inhibits non-small cell lung cancer growth and promotes chemosensitivity to cisplatin. Aging 2020, 12, 4711–4726. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, C.; Yang, Y.; Guo, L.; Huang, J.; Liu, X.; Wu, C.; Zou, J. Silencing of LncRNA-HOTAIR decreases drug resistance of Non-Small Cell Lung Cancer cells by inactivating autophagy via suppressing the phosphorylation of ULK1. Biochem. Biophys. Res. Commun. 2018, 497, 1003–1010. [Google Scholar] [CrossRef]
- Ke, C.; Feng, X.; Li, J.; Chen, S.; Hu, X. Association between long non-coding RNA HOTAIR polymorphism and lung cancer risk: A systematic review and meta-analysis. Exp. Ther. Med. 2022, 24, 540. [Google Scholar] [CrossRef]
- Suri, C.; Swarnkar, S.; Bhaskar, L.; Verma, H.K. Non-Coding RNA as a Biomarker in Lung Cancer. Non-Coding RNA 2024, 10, 50. [Google Scholar] [CrossRef]
- Rasul, M.F.; Hussen, B.M.; Salihi, A.; Ismael, B.S.; Jalal, P.J.; Zanichelli, A.; Jamali, E.; Baniahmad, A.; Ghafouri-Fard, S.; Basiri, A.; et al. Strategies to overcome the main challenges of the use of CRISPR/Cas9 as a replacement for cancer therapy. Mol. Cancer 2022, 21, 64. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, S.; Zhong, Y.; Lu, J.; Song, L. The Regulatory Role of LncRNAs in Modulating Autophagy and Drug Resistance in Non-Small-Cell Lung Cancer: Focus on Targeted Therapeutic Approaches. Biomolecules 2025, 15, 968. https://doi.org/10.3390/biom15070968
Dai S, Zhong Y, Lu J, Song L. The Regulatory Role of LncRNAs in Modulating Autophagy and Drug Resistance in Non-Small-Cell Lung Cancer: Focus on Targeted Therapeutic Approaches. Biomolecules. 2025; 15(7):968. https://doi.org/10.3390/biom15070968
Chicago/Turabian StyleDai, Shuncai, Yuxin Zhong, Jianfu Lu, and Linjiang Song. 2025. "The Regulatory Role of LncRNAs in Modulating Autophagy and Drug Resistance in Non-Small-Cell Lung Cancer: Focus on Targeted Therapeutic Approaches" Biomolecules 15, no. 7: 968. https://doi.org/10.3390/biom15070968
APA StyleDai, S., Zhong, Y., Lu, J., & Song, L. (2025). The Regulatory Role of LncRNAs in Modulating Autophagy and Drug Resistance in Non-Small-Cell Lung Cancer: Focus on Targeted Therapeutic Approaches. Biomolecules, 15(7), 968. https://doi.org/10.3390/biom15070968