
Ultrastructural Aspects of Physiological Mineralization: A Comparative Study in Different Hard Tissues
 , , ,
, , ,  , and
, and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
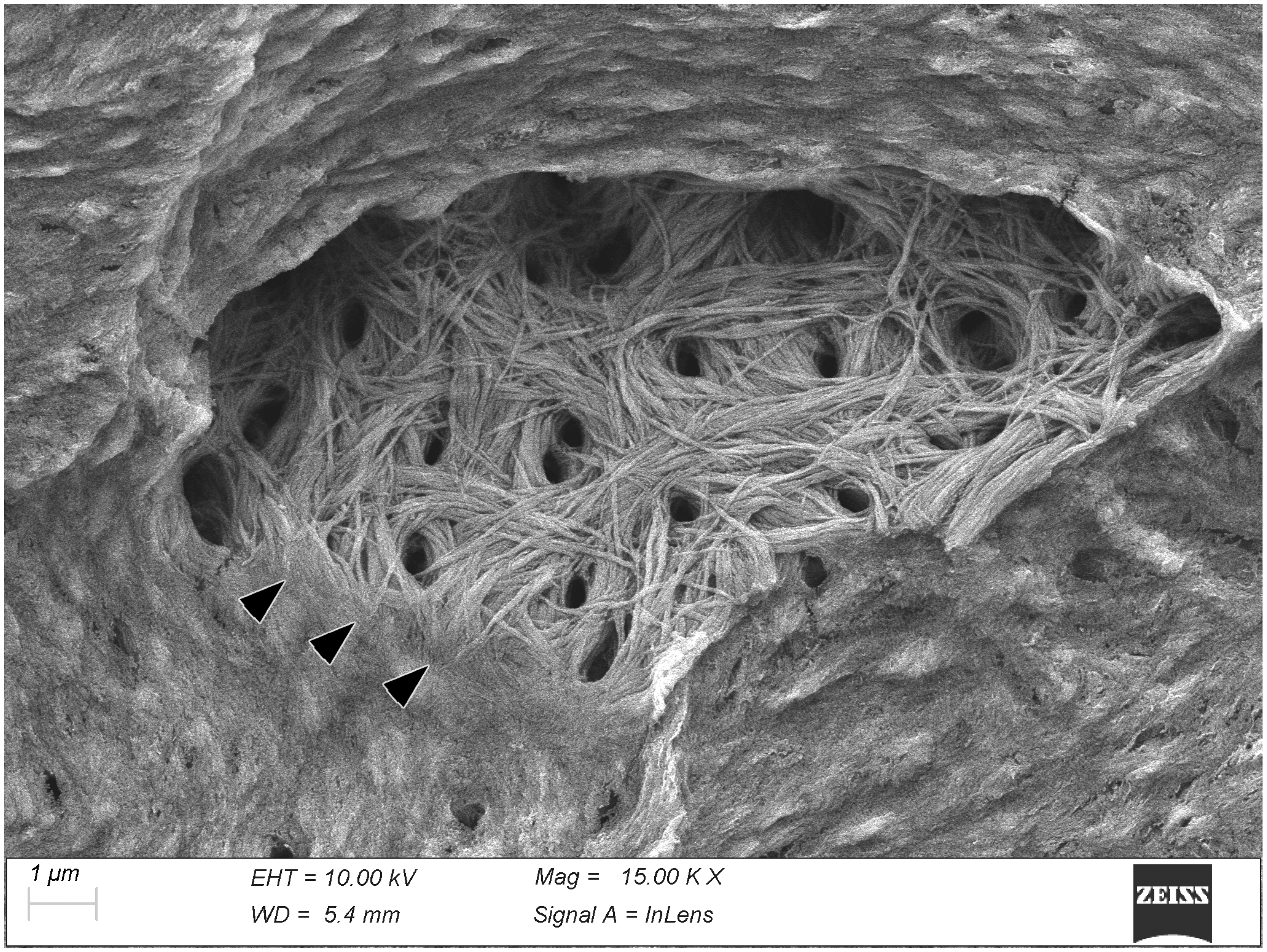
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahar, R.; Weiner, S. Open questions on the 3D structures of collagen containing vertebrate mineralized tissues: A perspective. J. Struct. Biol. 2018, 201, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Lausch, A.J.; Quan, B.C.; Miklas, J.W.; Sone, E.D. Extracellular Matrix Control of Collagen Mineralization In Vitro. Adv. Funct. Mater. 2013, 23, 4906–4912. [Google Scholar] [CrossRef]
- Reznikov, N.; Hoac, B.; Buss, D.J.; Addison, W.N.; Barros, N.M.T.; McKee, M.D. Biological stenciling of mineralization in the skeleton: Local enzymatic removal of inhibitors in the extracellular matrix. Bone 2020, 138, 115447. [Google Scholar] [CrossRef] [PubMed]
- Oosterlaken, B.M.; Vena, M.P.; de With, G. In Vitro Mineralization of Collagen. Adv. Mater. 2021, 33, 2004418. [Google Scholar] [CrossRef]
- Christoffersen, J.; Landis, W.J. A contribution with review to the description of mineralization of bone and other calcified tissues in vivo. Anat. Rec. 1991, 230, 435–450. [Google Scholar] [CrossRef]
- Landis, W.J.; Hodgens, K.J.; McKee, M.D.; Nanci, A.; Song, M.J.; Kivonaga, S.; Arena, J.; McEwen, B.F. Extracellular vesiscles of calcifying turkey leg tendon characterized by immunocytochemistry and high voltage electron microscopic tomography and 3D graphic image reconstruction. Bone Miner. 1992, 17, 237–241. [Google Scholar] [CrossRef]
- Mohr, W.; Görz, E. Does arteriosclerotic calcinosis of vessel walls imitate osteogenesis? Pathomorphological studies of arteriosclerotic plaque. Z. Kardiol. 2002, 91, 212–232. [Google Scholar] [CrossRef]
- Kirsch, T. Determinants of pathological mineralization. Curr. Opin. Rheumatol. 2006, 18, 174–180. [Google Scholar] [CrossRef]
- Duer, M.J.; Friscić, T.; Proudfoot, D.; Reid, D.G.; Schoppet, M.; Shanahan, C.M.; Skepper, J.N.; Wise, E.R. Mineral surface in calcified plaque is like that of bone: Further evidence for regulated mineralization. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 2030–2034. [Google Scholar] [CrossRef]
- Kim, K.M. Apoptosis and Calcification. Scanning Microsc. 1995, 9, 1137–1175. [Google Scholar]
- Li, M.; Wang, Z.W.; Fang, L.J.; Cheng, S.Q.; Wang, X.; Liu, N.F. Programmed cell death in atherosclerosis and vascular calcification. Cell Death Dis. 2022, 13, 467. [Google Scholar] [CrossRef]
- Bonetti, A.; Marchini, M.; Ortolani, F. Ectopic mineralization in heart valves: New insights from in vivo and in vitro procalcific models and promising perspectives on noncalcifiable bioengineered valves. J. Thorac. Dis. 2019, 11, 2126–2143. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, A.; Contin, M.; Marchini, M.; Ortolani, F. Ultrastructural and Immunohistochemical Detection of Hydroxyapatite Nucleating Role by rRNA and Nuclear Chromatin Derivatives in Aortic Valve Calcification: In Vitro and In Vivo Pro-Calcific Animal Models and Actual Calcific Disease in Humans. Int. J. Mol. Sci. 2023, 24, 2667. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Buchanan, A.V.; Weiss, K.M. Biomineralization in Humans: Making the Hard Choices in Life. Annu. Rev. Genet. 2009, 43, 119–142. [Google Scholar] [CrossRef] [PubMed]
- Landis, W.J. Mineral characterization in calcifying tissues: Atomic, molecular and macromolecular perspectives. Connect. Tissue Res. 1996, 34, 239–246. [Google Scholar] [CrossRef]
- Reznikov, N.; Bilton, M.; Lari, L.; Stevens, M.M.; Kröger, R. Fractal-like hierarchical organization of bone begins at the nanoscale. Science 2018, 360, 507. [Google Scholar] [CrossRef]
- Xu, Y.F.; Nudelman, F.; Eren, E.D.; Wirix, M.J.M.; Cantaert, B.; Nijhuis, W.H.; Hermida-Merino, D.; Portale, G.; Bomans, P.H.H.; Ottmann, C.; et al. Intermolecular channels direct crystal orientation in mineralized collagen. Nat. Commun. 2020, 11, 5068. [Google Scholar] [CrossRef]
- Landis, W.J.; Song, M.J.; Leith, A.; McEwen, L.; McEwen, B.F. Mineral and organic matrix interaction in normally calcifying tendon visualized in three dimensions by high-voltage electron microscopic tomography and graphic image reconstruction. J. Struct. Biol. 1993, 110, 39–54. [Google Scholar] [CrossRef]
- Siperko, L.M.; Landis, W.J. Aspects of mineral structure in normally calcifying avian tendon. J. Struct. Biol. 2001, 135, 313–320. [Google Scholar] [CrossRef]
- Landis, W.J.; Silver, F.H. The structure and function of normally mineralizing avian tendons. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 1135–1157. [Google Scholar] [CrossRef]
- Landis, W.J.; Hodgens, K.J.; Arena, J.; Song, M.J.; McEwen, B.F. Structural relations between collagen and mineral in bone as determined by high voltage electron microscopic tomography. Micros. Res. Tech. 1996, 33, 192–202. [Google Scholar] [CrossRef]
- Sorokina, L.; Shahbazian-Yassar, R.; Shokhufar, T. Collagen biomineralization: Pathways, mechanisms and thermodynamics. Emergent Mater. 2021, 4, 1025–1224. [Google Scholar] [CrossRef]
- Buss, D.J.; Rechav, K.; Reznikov, N.; McKee, M.D. Mineral tessellation in mouse enthesis fibrocartilage, Achilles tendon, and Hyp calcifying enthesopathy: A shared 3D mineralization pattern. Bone 2023, 174, 116818. [Google Scholar] [CrossRef] [PubMed]
- McKee, M.D.; Buss, D.J.; Reznikov, N. Mineral tessellation in bone and the stenciling principle for extracellular matrix mineralization. J. Struct. Biol. 2022, 214, 107823. [Google Scholar] [CrossRef]
- Micheletti, C.; Hurley, A.; Gourrier, A.; Palmquist, A.; Tang, T.; Shah, F.A.; Grandfield, K. Bone mineral organization at the mesoscale: A review of mineral ellipsoids in bone and at bone interfaces. Acta Biomater. 2022, 142, 1–13. [Google Scholar] [CrossRef]
- Kirsch, T.; Nah, H.-D.; Shapiro, I.M.; Pacifici, M. Regulated Production of Mineralization-competent Matrix Vesicles in Hypertrophic Chondrocytes. J. Cell Biol. 1997, 137, 1149–1160. [Google Scholar] [CrossRef]
- Kirsch, T.; Claassen, H. Matrix vesicles mediate mineralization of human thyroid cartilage. Calcif. Tissue Int. 2000, 66, 292–297. [Google Scholar] [CrossRef]
- Azoidis, I.; Cox, S.C.; Davies, O.G. The role of extracellular vesicles in biomineralisation: Current perspective and application in regenerative medicine. Tissue Eng. 2018, 9, 2041731418810130. [Google Scholar] [CrossRef]
- Yi, G.; Ma, Y.; Chen, Y.; Yang, X.; Yang, B.; Tian, W. A review of the functions of matrix vesicles in periodontal tissues. Stem Cells Dev. 2021, 30, 165–176. [Google Scholar] [CrossRef]
- Boyan, B.D.; Asmussen, N.C.; Lin, Z.; Schwartz, Z. The Role of Matrix-Bound Extracellular Vesicles in the Regulation of Endochondral Bone Formation. Cells 2022, 11, 1619. [Google Scholar] [CrossRef]
- Iwayama, T.; Bhongsatiern, P.; Takedachi, M.; Murakami, S. Matrix Vesicle–Mediated Mineralization and Potential Applications. J. Dent. Res. 2022, 101, 1554–1562. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Hongo, H.; Yamamoto, T.; Abe, M.; Yoshino, H.; Haraguchi-Kitakamae, M.; Ishizu, H.; Shimizu, T.; Iwasaki, N.; Amizuka, N. Matrix Vesicle-Mediated Mineralization and Osteocytic Regulation of Bone Mineralization. Int. J. Mol. Sci. 2022, 23, 9941. [Google Scholar] [CrossRef] [PubMed]
- Veschi, E.A.; Bolean, M.; da Silva Andrilli, L.H.; Gobbi Sebinelli, H.; Strzelecka-Kiliszek, A.; Bandorowicz-Pikula, J.; Pikula, S.; Granjon, T.; Mebarek, S.; Magne, D.; et al. Mineralization Profile of Annexin A6-Harbouring Proteoliposomes: Shedding Light on the Role of Annexin A6 on Matrix Vesicle-Mediated Mineralization. Int. J. Mol. Sci. 2022, 23, 8945. [Google Scholar] [CrossRef] [PubMed]
- Jaroszewicz, J.; Bazarnik, P.; Osiecka-Iwan, A.; Hyc, A.; Choinska, E.; Chlanda, A.; Swieszkowski, W.; Moskalewski, S. From Matrix Vesicles to Miniature Rocks: Evolution of Calcium Deposits in Calf Costochondral Junctions. Cartilage 2021, 13, 326S–335S. [Google Scholar] [CrossRef] [PubMed]
- Zecca, P.A.; Reguzzoni, M.; Protasoni, M.; Raspanti, M. The chondro-osseous junction of articular cartilage. Tissue Cell 2023, 80, 101993. [Google Scholar] [CrossRef]
- Jaroszewicz, J.; Kosowska, A.; Hutmacher, D.; Swieszkowski, W.; Moskalewski, S. Insight into characteristic features of cartilage growth plate as a physiological template for bone formation. J. Biomed. Mater. Res. A 2016, 104, 357–366. [Google Scholar] [CrossRef]
- Zecca, P.A.; Reguzzoni, M.; Borgese, M.; Protasoni, M.; Filibian, M.; Raspanti, M. Investigating the Interfaces of the Epiphyseal Plate: An Integrated Approach of Histochemistry, Microtomography and SEM. J. Anat. 2023, 243, 870–877. [Google Scholar] [CrossRef]
- Lories, R.J.; Luyten, F.P. The bone-cartilage unit in osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 43–49. [Google Scholar] [CrossRef]
- Zhou, X.; Cao, H.; Yuan, Y.; Wu, W. Biochemical Signals Mediate the Crosstalk between Cartilage and Bone in Osteoarthritis. BioMed Res. Int. 2020, 6, 5720360. [Google Scholar] [CrossRef]
- Raspanti, M.; Guizzardi, S.; De Pasquale, V.; Martini, D.; Ruggeri, A. Ultrastructure of heat-deproteinated compact bone. Biomaterials 1994, 15, 433–437. [Google Scholar] [CrossRef]
- Raspanti, M.; Cesari, C.; De Pasquale, V.; Ottani, V.; Strocchi, R.; Zucchelli, G.; Ruggeri, A. A histological and electron-microscopic study of the architecture and ultrastructure of human periodontal tissues. Arch. Oral. Biol. 2000, 45, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, H. The functional significance of dentin sialoprotein-phosphophoryn and dentin sialoprotein (Review). Int. J. Oral. Sci. 2018, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- Anada, R.; Satoshi Hara, E.; Nagaoka, N.; Okada, M.; Kamioka, H.; Matsumoto, T. Important roles of odontoblast membrane phospholipids in early dentin mineralization. J. Mater. Chem. B 2023, 11, 657–666. [Google Scholar] [CrossRef]
- Silver, F.H.; Landis, W.J. Deposition of apatite in mineralizing vertebrate extracellular matrices: A model of possible nucleation sites on type I collagen. Connect. Tissue Res. 2011, 52, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Ortolani, F.; Giordano, M.; Marchini, M. A model for type II collagen fibrils. Distinctive D-band patterns in native and reconstituted fibrils compared with sequence data for helix and telopeptide domains. Biopolymers 2000, 54, 448–463. [Google Scholar] [CrossRef]
- Antipova, O.; Orgel, J.P.R.O. In Situ D-periodic Molecular Structure of Type II Collagen. J. Biol. Chem. 2010, 285, 7087–7096. [Google Scholar] [CrossRef]
- Müller, K.H.; Hayward, R.; Rajan, R.; Whitehead, M.; Cobb, A.M.; Ahmad, S.; Sun, M.; Goldberga, I.; Li, R.; Bashtanova, U.; et al. Poly(ADP-Ribose) Links the DNA Damage Response and Biomineralization. Cell Rep. 2019, 27, 3124–3138. [Google Scholar] [CrossRef]
- Zecca, M.A.; Greer, H.F.; Duer, M.J. Poly(ADP-ribose) binding sites on collagen I fibrils for nucleating intrafibrillar bone mineral. bioRxiv 2024. [Google Scholar] [CrossRef]
- Gordon, M.K.; Gerecke, D.R.; Dublet, B.; van der Rest, M.; Olsen, B. Type XII Collagen: A Large Multidomain Molecule With Partial Homology to Type IX Collagen. J. Biol. Chem. 1989, 264, 19772–19778. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borgese, M.; Raspanti, M.; Protasoni, M.; Zecca, P.A.; Ortolani, F.; Reguzzoni, M. Ultrastructural Aspects of Physiological Mineralization: A Comparative Study in Different Hard Tissues. Biomolecules 2025, 15, 932. https://doi.org/10.3390/biom15070932
Borgese M, Raspanti M, Protasoni M, Zecca PA, Ortolani F, Reguzzoni M. Ultrastructural Aspects of Physiological Mineralization: A Comparative Study in Different Hard Tissues. Biomolecules. 2025; 15(7):932. https://doi.org/10.3390/biom15070932
Chicago/Turabian StyleBorgese, Marina, Mario Raspanti, Marina Protasoni, Piero Antonio Zecca, Fulvia Ortolani, and Marcella Reguzzoni. 2025. "Ultrastructural Aspects of Physiological Mineralization: A Comparative Study in Different Hard Tissues" Biomolecules 15, no. 7: 932. https://doi.org/10.3390/biom15070932
APA StyleBorgese, M., Raspanti, M., Protasoni, M., Zecca, P. A., Ortolani, F., & Reguzzoni, M. (2025). Ultrastructural Aspects of Physiological Mineralization: A Comparative Study in Different Hard Tissues. Biomolecules, 15(7), 932. https://doi.org/10.3390/biom15070932