

Dairy Propionibacteria: Probiotic Properties and Their Molecular Bases

 , ,
, ,
Abstract
1. Introduction
2. Molecular Traits of Dairy Propionibacteria Relevant to Probiotic Activities
2.1. Genome Features of Dairy Propionibacteria
2.2. Production of Exopolysaccharides
2.3. Cell Surface Proteins
2.4. Extracellular Vesicles
3. Beneficial In Vivo Effects of Dairy Propionibacteria
3.1. Prevention and Mitigation of Intestinal Inflammatory Diseases
3.2. Immunomodulation
3.3. Effects on Obesity
3.4. Anti-Cancer Effects
3.5. Effects of Propionibacterium freudenreichii on Bone Health
4. Antimicrobial Properties of Dairy Propionibacteria
5. Production of Beneficial Metabolites
6. Safety of Dairy Propionibacteria
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACNQ | 2-amino-3-carboxy-1,4-naphthoquinone |
| ALT | Alanine aminotransferase |
| BALs | Bovine alveolar lavage cells |
| BCFA | Branched-chain fatty acid |
| CFU | Colony-forming unit |
| CRISPR-Cas | Clustered regularly interspaced short palindromic repeats |
| CW | Cell wall |
| DAI | Disease activity index |
| DBL | DNA-binding ligand |
| DC | Dendritic cell |
| DFM | Direct fed microbial |
| DHNA | 1,4-dihydroxy-2-naphtoic acid |
| DSS | Dextran sulfate |
| DTPA | Diethylenetriamine pentaacetic acid |
| EFSA | European Food Safety Authority |
| ELISA | Enzyme-linked immunosorbent assay |
| EPS | Exopolysaccharide |
| EVs | Extracellular vesicles |
| FIL-IDF | International Dairy Federation |
| FITC | Fluorescein isothiocyanate |
| 5-FU | 5-fluorouracile |
| GIT | Gastrointestinal tract |
| GRAS | Generally recognized as safe |
| HDL | High-density lipoprotein cholesterol |
| HFD | High-fat diet |
| HGT | Horizontal gene transfer |
| HOMA-IR | Homeostasis model assessment insulin resistance |
| HRP | Horseradish peroxidase |
| IBD | Inflammatory bowel disease |
| ICE | Integrative–conjugative element |
| IEB | Intestinal epithelial barrier |
| IFN-γ | Interferon γ |
| Ig | Immunoglobulin |
| IHC | Immunohistochemistry |
| IL | Interleukin |
| LDL | Low-density lipoprotein cholesterol |
| LPS | Lipopolysaccharide |
| MAdCAM-1 | Mucosal addressin cell adhesion molecule 1 |
| MAPK | Mitogen-activated protein kinase |
| MHC II | Major histocompatibility complex II |
| MLNC | Mesenteric lymph node immune cell |
| mMCP-1 | Mucosal mast cell protease-1 |
| MoDC | Monocyte-derived DC |
| MSCs | Mesenchymal stem cells |
| MTT | 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide |
| NEC | Necrotizing enterocolitis |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK | Natural killer |
| PBMCs | Peripheral blood mononuclear cells |
| QPS | Qualified presumption of safety |
| qRT-PCR | Quantitative reverse transcriptase polymerase chain reaction |
| SCFA | Short-chain fatty acid |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel |
| SLH | Surface layer homology |
| Slp | S-layer protein |
| SP | Surface protein |
| Th | T helper |
| TEER | Trans-epithelial electrical resistance |
| TGF-β | Tumor growth factor-β |
| TNBS | 2,4,6-trinitrobenzenesulfonic acid |
| TNF-α | Tumor necrosis factor α |
| Treg | Regulatory T cell |
| UC | Ulcerative colitis |
| UF | Ultra-filtered |
| XRE | Xenobiotic-responsive element |
| YEL | Yeast extract lactate |
References
- Scholz, C.F.P.; Kilian, M. The Natural History of Cutaneous Propionibacteria, and Reclassification of Selected Species within the Genus Propionibacterium to the Proposed Novel Genera Acidipropionibacterium gen. nov., Cutibacterium gen. nov. and Pseudopropionibacterium gen. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 4422–4432. [Google Scholar] [CrossRef] [PubMed]
- Deptula, P.; Laine, P.K.; Roberts, R.J.; Smolander, O.-P.; Vihinen, H.; Piironen, V.; Paulin, L.; Jokitalo, E.; Savijoki, K.; Auvinen, P.; et al. De Novo Assembly of Genomes from Long Sequence Reads Reveals Uncharted Territories of Propionibacterium freudenreichii. BMC Genom. 2017, 18, 790. [Google Scholar] [CrossRef]
- Deptula, P.; Smolander, O.-P.; Laine, P.; Roberts, R.J.; Edelmann, M.; Peltola, P.; Piironen, V.; Paulin, L.; Storgårds, E.; Savijoki, K.; et al. Acidipropionibacterium virtanenii sp. nov., Isolated from Malted Barley. Int. J. Syst. Evol. Microbiol. 2018, 68, 3175–3183. [Google Scholar] [CrossRef] [PubMed]
- Togo, A.H.; Diop, A.; Camara, A.; Kuete, E.; Konate, S.; Brevaut, V.; Des Robert, C.; Delerce, J.; Armstrong, N.; Roussel, Y.; et al. Lactimicrobium Massiliense gen. nov., sp. nov.; Anaerolactibacter massiliensis gen. nov., sp. nov.; Galactobacillus timonensis gen. nov., sp. nov. and Acidipropionibacterium timonense sp. nov. Isolated from Breast Milk from Healthy Breastfeeding African Women. New Microbes New Infect. 2019, 29, 100537. [Google Scholar] [CrossRef] [PubMed]
- McCubbin, T.; Gonzalez-Garcia, R.A.; Palfreyman, R.W.; Stowers, C.; Nielsen, L.K.; Marcellin, E. A Pan-Genome Guided Metabolic Network Reconstruction of Five Propionibacterium Species Reveals Extensive Metabolic Diversity. Genes 2020, 11, 1115. [Google Scholar] [CrossRef]
- Dank, A.; van Mastrigt, O.; Boeren, S.; Lillevang, S.K.; Abee, T.; Smid, E.J. Propionibacterium freudenreichii Thrives in Microaerobic Conditions by Complete Oxidation of Lactate to CO2. Environ. Microbiol. 2021, 23, 3116–3129. [Google Scholar] [CrossRef]
- de Assis, D.A.; Machado, C.; Matte, C.; Ayub, M.A.Z. High Cell Density Culture of Dairy Propionibacterium Sp. and Acidipropionibacterium Sp.: A Review for Food Industry Applications. Food Bioprocess Technol. 2022, 15, 734–749. [Google Scholar] [CrossRef]
- Cousin, F.J.; Foligné, B.; Deutsch, S.-M.; Massart, S.; Parayre, S.; Le Loir, Y.; Boudry, G.; Jan, G. Assessment of the Probiotic Potential of a Dairy Product Fermented by Propionibacterium freudenreichii in Piglets. J. Agric. Food Chem. 2012, 60, 7917–7927. [Google Scholar] [CrossRef]
- Casanova, M.R.; Azevedo-Silva, J.; Rodrigues, L.R.; Preto, A. Colorectal Cancer Cells Increase the Production of Short Chain Fatty Acids by Propionibacterium freudenreichii Impacting on Cancer Cells Survival. Front. Nutr. 2018, 5, 44. [Google Scholar] [CrossRef]
- Rossi, F.; Torriani, S.; Dellaglio, F. Identification and Clustering of Dairy Propionibacteria by RAPD-PCR and CGE-REA Methods. J. Appl. Microbiol. 1998, 85, 956–964. [Google Scholar] [CrossRef]
- Rossi, F.; Dellaglio, F. Quality of Silages from Italian Farms as Attested by Number and Identity of Microbial Indicators: Microflora of Farm Made Silages. J. Appl. Microbiol. 2007, 103, 1707–1715. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, R.; Chuat, V.; Madec, M.-N.; Nero, L.A.; Thierry, A.; Valence, F.; de Carvalho, A.F. Biodiversity of Dairy Propionibacterium Isolated from Dairy Farms in Minas Gerais, Brazil. Int. J. Food Microbiol. 2015, 203, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Martínez, E.A.; Babot, J.D.; Lorenzo-Pisarello, M.J.; Apella, M.C.; Chaia, A.P. Feed Supplementation with Avian Propionibacterium acidipropionici Contributes to Mucosa Development in Early Stages of Rearing Broiler Chickens. Benef. Microbes 2016, 7, 687–698. [Google Scholar] [CrossRef]
- Colliou, N.; Ge, Y.; Sahay, B.; Gong, M.; Zadeh, M.; Owen, J.L.; Neu, J.; Farmerie, W.G.; Alonzo, F., 3rd; Liu, K.; et al. Commensal Propionibacterium Strain UF1 Mitigates Intestinal Inflammation via Th17 Cell Regulation. J. Clin. Investig. 2017, 127, 3970–3986. [Google Scholar] [CrossRef]
- Coronas, R.; Zara, G.; Gallo, A.; Rocchetti, G.; Lapris, M.; Petretto, G.L.; Zara, S.; Fancello, F.; Mannazzu, I. Propionibacteria as Promising Tools for the Production of Pro-Bioactive Scotta: A Proof-of-Concept Study. Front. Microbiol. 2023, 14, 1223741. [Google Scholar] [CrossRef]
- Amadoro, C.; Rossi, F.; Pallotta, M.L.; Gasperi, M.; Colavita, G. Traditional Dairy Products Can Supply Beneficial Microorganisms Able to Survive in the Gastrointestinal Tract. LWT 2018, 93, 376–383. [Google Scholar] [CrossRef]
- Rossi, F.; Capilongo, V.; Torriani, S. Confronto tra diversi terreni selettivi per la conta e l’isolamento di batteri propionici in latte ovino. L’Industria Latte 1996, 32, 33–43. [Google Scholar]
- Daghio, M.; Pini, F.; Espinoza-Tofalos, A.; Conte, G.; Mari, E.; Giannerini, F.; Giovannetti, L.; Buccioni, A.; Franzetti, A.; Granchi, L.; et al. Characterization of the Microbial Community in Ripened Pecorino Toscano Cheese Affected by Pink Discoloration. Food Microbiol. 2022, 104, 104006. [Google Scholar] [CrossRef] [PubMed]
- Bücher, C.; Burtscher, J.; Zitz, U.; Domig, K.J. One-Year Monitoring of Prevalence and Diversity of Dairy Propionic Acid Bacteria in Raw Milk by Means of Culture-Dependent and Culture-Independent Methods. Foods 2024, 13, 1921. [Google Scholar] [CrossRef]
- Carafa, I.; Navarro, I.C.; Bittante, G.; Tagliapietra, F.; Gallo, L.; Tuohy, K.; Franciosi, E. Shift in the Cow Milk Microbiota during Alpine Pasture as Analyzed by Culture Dependent and High-Throughput Sequencing Techniques. Food Microbiol. 2020, 91, 103504. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 8, 506–514. [Google Scholar] [CrossRef]
- Mantere-Alhonen, S. Propionibacteria Used as Probiotics—A Review. Lait 1995, 75, 447–452. [Google Scholar] [CrossRef]
- Aprea, G.; Del Matto, I.; Tucci, P.; Marino, L.; Scattolini, S.; Rossi, F. In Vivo Functional Properties of Dairy Bacteria. Microorganisms 2023, 11, 1787. [Google Scholar] [CrossRef]
- Rabah, H.; Rosa do Carmo, F.L.; Jan, G. Dairy Propionibacteria: Versatile Probiotics. Microorganisms 2017, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ). Update of the List of Qualified Presumption of Safety (QPS) Recommended Microbiological Agents Intentionally Added to Food or Feed as Notified to EFSA 21: Suitability of Taxonomic Units Notified to EFSA until September 2024. EFSA J. 2025, 23, e9169. [Google Scholar] [CrossRef]
- Bourdichon, F.; Budde-Niekiel, A.; Dubois, A.; Fritz, D.; Hatte, J.-L.; Laulund, S.; McAuliffe, O.; Ouwehand, A.C.; Yao, S.; Zgoda, A.; et al. Bulletin of the International Dairy Federation Inventory of Microbial Food Cultures with Safety Demonstration in Fermented Food Products; Bulletin of The International Dairy Federation 514/2022; International Dairy Federation: Brussels, Belgium, 2022. [Google Scholar]
- Soto-Serrano, A.; Li, W.; Panah, F.M.; Hui, Y.; Atienza, P.; Fomenkov, A.; Roberts, R.J.; Deptula, P.; Krych, L. Matching Excellence: Oxford Nanopore Technologies’ Rise to Parity with Pacific Biosciences in Genome Reconstruction of Non-Model Bacterium with High G+C Content. Microb. Genom. 2024, 10, 001316. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Dellaglio, F.; Torriani, S. Evaluation of recA Gene as a Phylogenetic Marker in the Classification of Dairy Propionibacteria. Syst. Appl. Microbiol. 2006, 29, 463–469. [Google Scholar] [CrossRef]
- Rossi, F.; Amadoro, C.; Pallotta, M.L.; Colavita, G. Variability of Genetic Characters Associated with Probiotic Functions in Lacticaseibacillus Species. Microorganisms 2022, 10, 1023. [Google Scholar] [CrossRef]
- Deutsch, S.-M.; Le Bivic, P.; Hervé, C.; Madec, M.-N.; LaPointe, G.; Jan, G.; Le Loir, Y.; Falentin, H. Correlation of the Capsular Phenotype in Propionibacterium freudenreichii with the Level of Expression of Gtf, a Unique Polysaccharide Synthase-Encoding Gene. Appl. Environ. Microbiol. 2010, 76, 2740–2746. [Google Scholar] [CrossRef]
- Deutsch, S.-M.; Parayre, S.; Bouchoux, A.; Guyomarc’h, F.; Dewulf, J.; Dols-Lafargue, M.; Baglinière, F.; Cousin, F.J.; Falentin, H.; Jan, G.; et al. Contribution of Surface β-Glucan Polysaccharide to Physicochemical and Immunomodulatory Properties of Propionibacterium freudenreichii. Appl. Environ. Microbiol. 2012, 78, 1765–1775. [Google Scholar] [CrossRef] [PubMed]
- Le Maréchal, C.; Peton, V.; Plé, C.; Vroland, C.; Jardin, J.; Briard-Bion, V.; Durant, G.; Chuat, V.; Loux, V.; Foligné, B.; et al. Surface Proteins of Propionibacterium freudenreichii Are Involved in Its Anti-Inflammatory Properties. J. Proteom. 2015, 113, 447–461. [Google Scholar] [CrossRef]
- Frohnmeyer, E.; Deptula, P.; Nyman, T.A.; Laine, P.K.S.; Vihinen, H.; Paulin, L.; Auvinen, P.; Jokitalo, E.; Piironen, V.; Varmanen, P.; et al. Secretome Profiling of Propionibacterium freudenreichii Reveals Highly Variable Responses Even among the Closely Related Strains. Microb. Biotechnol. 2018, 11, 510–526. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Busetto, M.; Torriani, S. Isolation of Aminopeptidase N Genes of Food Associated Propionibacteria and Observation of Their Transcription in Skim Milk and Acid Whey. Antonie Van Leeuwenhoek 2007, 91, 87–96. [Google Scholar] [CrossRef]
- do Carmo, F.L.R.; Silva, W.M.; Tavares, G.C.; Ibraim, I.C.; Cordeiro, B.F.; Oliveira, E.R.; Rabah, H.; Cauty, C.; da Silva, S.H.; Canário Viana, M.V.; et al. Mutation of the Surface Layer Protein SlpB Has Pleiotropic Effects in the Probiotic Propionibacterium freudenreichii CIRM-BIA 129. Front. Microbiol. 2018, 9, 1807. [Google Scholar] [CrossRef]
- Deutsch, S.-M.; Mariadassou, M.; Nicolas, P.; Parayre, S.; Le Guellec, R.; Chuat, V.; Peton, V.; Le Maréchal, C.; Burati, J.; Loux, V.; et al. Identification of Proteins Involved in the Anti-Inflammatory Properties of Propionibacterium freudenreichii by Means of a Multi-Strain Study. Sci. Rep. 2017, 7, 46409. [Google Scholar] [CrossRef] [PubMed]
- do Carmo, F.L.R.; Rabah, H.; Huang, S.; Gaucher, F.; Deplanche, M.; Dutertre, S.; Jardin, J.; Le Loir, Y.; Azevedo, V.; Jan, G. Propionibacterium freudenreichii Surface Protein SlpB Is Involved in Adhesion to Intestinal HT-29 Cells. Front. Microbiol. 2017, 8, 1033. [Google Scholar] [CrossRef]
- Rabah, H.; Ménard, O.; Gaucher, F.; do Carmo, F.L.R.; Dupont, D.; Jan, G. Cheese Matrix Protects the Immunomodulatory Surface Protein SlpB of Propionibacterium freudenreichii during in Vitro Digestion. Food Res. Int. 2018, 106, 712–721. [Google Scholar] [CrossRef]
- Mantel, M.; Durand, T.; Bessard, A.; Pernet, S.; Beaudeau, J.; Guimaraes-Laguna, J.; Maillard, M.-B.; Guédon, E.; Neunlist, M.; Le Loir, Y.; et al. Propionibacterium freudenreichii CIRM-BIA 129 Mitigates Colitis through S Layer Protein B-Dependent Epithelial Strengthening. Am. J. Physiol. Gastrointest. Liver Physiol. 2024, 326, G163–G175. [Google Scholar] [CrossRef]
- Rodovalho, V.d.R.; da Luz, B.S.R.; Rabah, H.; do Carmo, F.L.R.; Folador, E.L.; Nicolas, A.; Jardin, J.; Briard-Bion, V.; Blottière, H.; Lapaque, N.; et al. Extracellular Vesicles Produced by the Probiotic Propionibacterium freudenreichii CIRM-BIA 129 Mitigate Inflammation by Modulating the NF-κB Pathway. Front. Microbiol. 2020, 11, 1544. [Google Scholar] [CrossRef]
- de Rezende Rodovalho, V.; da Luz, B.S.R.; Nicolas, A.; do Carmo, F.L.R.; Jardin, J.; Briard-Bion, V.; Jan, G.; Le Loir, Y.; de Carvalho Azevedo, V.A.; Guedon, E. Environmental Conditions Modulate the Protein Content and Immunomodulatory Activity of Extracellular Vesicles Produced by the Probiotic Propionibacterium freudenreichii. Appl. Environ. Microbiol. 2021, 87, e02263-20. [Google Scholar] [CrossRef] [PubMed]
- Rodovalho, V.d.R.; da Luz, B.S.R.; Nicolas, A.; Jardin, J.; Briard-Bion, V.; Folador, E.L.; Santos, A.R.; Jan, G.; Loir, Y.L.; Azevedo, V.A.d.C.; et al. Different Culture Media and Purification Methods Unveil the Core Proteome of Propionibacterium freudenreichii-Derived Extracellular Vesicles. Microlife 2023, 4, uqad029. [Google Scholar] [CrossRef] [PubMed]
- Uchida, M.; Mogami, O. Milk Whey Culture with Propionibacterium freudenreichii ET-3 Is Effective on the Colitis Induced by 2,4,6-Trinitrobenzene Sulfonic Acid in Rats. J. Pharmacol. Sci. 2005, 99, 329–334. [Google Scholar] [CrossRef]
- Okada, Y.; Tsuzuki, Y.; Miyazaki, J.; Matsuzaki, K.; Hokari, R.; Komoto, S.; Kato, S.; Kawaguchi, A.; Nagao, S.; Itoh, K.; et al. Propionibacterium freudenreichii Component 1.4-Dihydroxy-2-Naphthoic Acid (DHNA) Attenuates Dextran Sodium Sulphate Induced Colitis by Modulation of Bacterial Flora and Lymphocyte Homing. Gut 2006, 55, 681–688. [Google Scholar] [CrossRef]
- Fukumoto, S.; Toshimitsu, T.; Matsuoka, S.; Maruyama, A.; Oh-Oka, K.; Takamura, T.; Nakamura, Y.; Ishimaru, K.; Fujii-Kuriyama, Y.; Ikegami, S.; et al. Identification of a Probiotic Bacteria-Derived Activator of the Aryl Hydrocarbon Receptor That Inhibits Colitis. Immunol. Cell Biol. 2014, 92, 460–465. [Google Scholar] [CrossRef]
- Plé, C.; Richoux, R.; Jardin, J.; Nurdin, M.; Briard-Bion, V.; Parayre, S.; Ferreira, S.; Pot, B.; Bouguen, G.; Deutsch, S.-M.; et al. Single-strain starter experimental cheese reveals anti-inflammatory effect of Propionibacterium freudenreichii CIRM BIA 129 in TNBS-colitis model. J. Funct. Foods 2015, 18, 575–585. [Google Scholar] [CrossRef]
- Plé, C.; Breton, J.; Richoux, R.; Nurdin, M.; Deutsch, S.-M.; Falentin, H.; Hervé, C.; Chuat, V.; Lemée, R.; Maguin, E.; et al. Combining Selected Immunomodulatory Propionibacterium freudenreichii and Lactobacillus delbrueckii Strains: Reverse Engineering Development of an Anti-Inflammatory Cheese. Mol. Nutr. Food Res. 2016, 60, 935–948. [Google Scholar] [CrossRef]
- Rabah, H.; do Carmo, F.L.R.; Carvalho, R.D.d.O.; Cordeiro, B.F.; da Silva, S.H.; Oliveira, E.R.; Lemos, L.; Cara, D.C.; Faria, A.M.C.; Garric, G.; et al. Beneficial Propionibacteria within a Probiotic Emmental Cheese: Impact on Dextran Sodium Sulphate-Induced Colitis in Mice. Microorganisms 2020, 8, 380. [Google Scholar] [CrossRef]
- Mantel, M.; da Silva, T.F.; Gloria, R.; Vassaux, D.; Vital, K.D.; Cardoso, V.N.; Fernandes, S.O.A.; Guédon, É.; Le Loir, Y.; Faria, A.M.C.; et al. Fat Matters: Fermented Whole Milk Potentiates the Anti-Colitis Effect of Propionibacterium freudenreichii. J. Funct. Foods 2023, 106, 105614. [Google Scholar] [CrossRef]
- Ma, S.; Yeom, J.; Lim, Y.-H. Dairy Propionibacterium freudenreichii Ameliorates Acute Colitis by Stimulating MUC2 Expression in Intestinal Goblet Cell in a DSS-Induced Colitis Rat Model. Sci. Rep. 2020, 10, 5523. [Google Scholar] [CrossRef]
- Yang, S.; Shang, J.; Liu, L.; Tang, Z.; Meng, X. Strains Producing Different Short-Chain Fatty Acids Alleviate DSS-Induced Ulcerative Colitis by Regulating Intestinal Microecology. Food Funct. 2022, 13, 12156–12169. [Google Scholar] [CrossRef] [PubMed]
- do Carmo, F.L.R.; Rabah, H.; Cordeiro, B.F.; da Silva, S.H.; Pessoa, R.M.; Fernandes, S.O.A.; Cardoso, V.N.; Gagnaire, V.; Deplanche, M.; Savassi, B.; et al. Probiotic Propionibacterium freudenreichii Requires SlpB Protein to Mitigate Mucositis Induced by Chemotherapy. Oncotarget 2019, 10, 7198–7219. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Gong, M.; Colliou, N.; Zadeh, M.; Li, J.; Jones, D.P.; Li, S.; Mohamadzadeh, M. Neonatal Intestinal Immune Regulation by the Commensal Bacterium, P. UF1. Mucosal Immunol. 2019, 12, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.C.; Lean, M.L.; Hitchick, N.C.; Beagley, K.W. The Efficacy of Propionibacterium jensenii 702 to Stimulate a Cell-Mediated Response to Orally Administered Soluble Mycobacterium tuberculosis Antigens Using a Mouse Model. Lait 2005, 85, 75–84. [Google Scholar] [CrossRef]
- Kwon, G.; Lee, J.; Lim, Y.-H. Dairy Propionibacterium Extends the Mean Lifespan of Caenorhabditis elegans via Activation of the Innate Immune System. Sci. Rep. 2016, 6, 31713. [Google Scholar] [CrossRef]
- Misme-Aucouturier, B.; Gagnaire, V.; LeCorre, E.; DeCarvalho, M.; Jan, G.; Bouchaud, G. Propionibacterium freudenreichii Prevents Food Allergy in Mice via the Surface Layer Protein SlpB. J. Agric. Food Chem. 2024, 72, 27495–27503. [Google Scholar] [CrossRef]
- Oksaharju, A.; Kooistra, T.; Kleemann, R.; van Duyvenvoorde, W.; Miettinen, M.; Lappalainen, J.; Lindstedt, K.A.; Kovanen, P.T.; Korpela, R.; Kekkonen, R.A. Effects of Probiotic Lactobacillus Rhamnosus GG and Propionibacterium freudenreichii ssp. Shermanii JS Supplementation on Intestinal and Systemic Markers of Inflammation in ApoE*3Leiden Mice Consuming a High-Fat Diet. Br. J. Nutr. 2013, 110, 77–85. [Google Scholar] [CrossRef]
- An, M.; Park, Y.-H.; Lim, Y.-H. Antiobesity and Antidiabetic Effects of the Dairy Bacterium Propionibacterium freudenreichii MJ2 in High-Fat Diet-Induced Obese Mice by Modulating Lipid Metabolism. Sci. Rep. 2021, 11, 2481. [Google Scholar] [CrossRef]
- An, M.; Lim, Y.-H. Surface-Exposed Chaperonin 60 Derived from Propionibacterium freudenreichii MJ2 Inhibits Adipogenesis by Decreasing the Expression of C/EBPα/PPARγ. Sci. Rep. 2023, 13, 19251. [Google Scholar] [CrossRef]
- Miyamoto, J.; Ando, Y.; Yamano, M.; Nishida, A.; Murakami, K.; Kimura, I. Acidipropionibacterium acidipropionici, a Propionate-Producing Bacterium, Contributes to GPR41 Signaling and Metabolic Regulation in High-Fat Diet-Induced Obesity in Mice. Front. Nutr. 2025, 12, 1542196. [Google Scholar] [CrossRef]
- Dikeocha, I.J.; Al-Kabsi, A.M.; Ahmeda, A.F.; Mathai, M.; Alshawsh, M.A. Investigation into the Potential Role of Propionibacterium freudenreichii in Prevention of Colorectal Cancer and Its Effects on the Diversity of Gut Microbiota in Rats. Int. J. Mol. Sci. 2023, 24, 8080. [Google Scholar] [CrossRef]
- Pérez Chaia, A.; Zarate, G.; Oliver, G. The Probiotic Properties of Propionibacteria. Lait 1999, 79, 175–185. [Google Scholar] [CrossRef]
- Lan, A.; Bruneau, A.; Bensaada, M.; Philippe, C.; Bellaud, P.; Rabot, S.; Jan, G. Increased Induction of Apoptosis by Propionibacterium freudenreichii TL133 in Colonic Mucosal Crypts of Human Microbiota-Associated Rats Treated with 1,2-Dimethylhydrazine. Br. J. Nutr. 2008, 100, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- Zárate, G.; Pérez Chaia, A. Feeding with Dairy Propionibacterium Acidipropionici CRL 1198 Reduces the Incidence of Concanavalin-A Induced Alterations in Mouse Small Intestinal Epithelium. Food Res. Int. 2012, 47, 13–22. [Google Scholar] [CrossRef]
- Yeom, J.; Ma, S.; Yim, D.J.; Lim, Y.-H. Surface Proteins of Propionibacterium freudenreichii MJ2 Inhibit RANKL-Induced Osteoclast Differentiation by Lipocalin-2 Upregulation and Lipocalin-2-Mediated NFATc1 Inhibition. Sci. Rep. 2023, 13, 15644. [Google Scholar] [CrossRef]
- Woo, H.-E.; Cho, J.-Y.; Lim, Y.-H. Propionibacterium freudenreichii MJ2-Derived Extracellular Vesicles Inhibit RANKL-Induced Osteoclastogenesis and Improve Collagen-Induced Rheumatoid Arthritis. Sci. Rep. 2024, 14, 24973. [Google Scholar] [CrossRef]
- Yeom, J.; Yim, D.J.; Ma, S.; Lim, Y.-H. Propionibacterium freudenreichii Inhibits RANKL-Induced Osteoclast Differentiation and Ameliorates Rheumatoid Arthritis in Collagen-Induced Arthritis Mice. Microorganisms 2021, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.V.T.; Johnson, T.J.; Noll, S.L.; Kollanoor Johny, A. Effect of Supplementation of a Dairy-Originated Probiotic Bacterium, Propionibacterium freudenreichii subsp. freudenreichii, on the Cecal Microbiome of Turkeys Challenged with Multidrug-Resistant Salmonella Heidelberg. Poult. Sci. 2021, 100, 283–295. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Vazhakkattu Thomas, J.; Dewi, G.; Brannon, J.; Noll, S.L.; Johnson, T.J.; Cox, R.B.; Kollanoor Johny, A. Propionibacterium freudenreichii freudenreichii B3523 Reduces Cecal Colonization and Internal Organ Dissemination of Multidrug-Resistant Salmonella Heidelberg in Finishing Turkeys. J. Appl. Poult. Res. 2021, 30, 100107. [Google Scholar] [CrossRef]
- Manjankattil, S.; Dewi, G.; Peichel, C.; Creek, M.; Bina, P.; Lerohl, K.; Deniz, K.; Akhtar, L.; Porter, R.; Johnson, T.J.; et al. Dairy-origin Propionibacterium freudenreichii, turkey-origin Lactobacillus salivarius, and a Salmonella typhimurium vaccine elicit comparable colonization resistance on drug-resistant Salmonella serotypes (S. reading, S. agona, and S. saintpaul) in growing turkeys after oral challenge. J. Appl. Poult. Res. 2024, 33, 100428. [Google Scholar] [CrossRef]
- Huang, Y.; Kotula, L.; Adams, M.C. The in Vivo Assessment of Safety and Gastrointestinal Survival of an Orally Administered Novel Probiotic, Propionibacterium jensenii 702, in a Male Wistar Rat Model. Food Chem. Toxicol. 2003, 41, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Suomalainen, T.; Sigvart-Mattila, P.; Mättö, J.; Tynkkynen, S. In Vitro and in Vivo Gastrointestinal Survival, Antibiotic Susceptibility and Genetic Identification of Propionibacterium freudenreichii ssp. Shermanii JS. Int. Dairy J. 2008, 18, 271–278. [Google Scholar] [CrossRef]
- Stein, D.R.; Allen, D.T.; Perry, E.B.; Bruner, J.C.; Gates, K.W.; Rehberger, T.G.; Mertz, K.; Jones, D.; Spicer, L.J. Effects of Feeding Propionibacteria to Dairy Cows on Milk Yield, Milk Components, and Reproduction. J. Dairy Sci. 2006, 89, 111–125. [Google Scholar] [CrossRef]
- Adams, M.C.; Luo, J.; Rayward, D.; King, S.; Gibson, R.; Moghaddam, G.H. Selection of a Novel Direct-Fed Microbial to Enhance Weight Gain in Intensively Reared Calves. Anim. Feed Sci. Technol. 2008, 145, 41–52. [Google Scholar] [CrossRef]
- Luo, J.; King, S.; Adams, M.C. Effect of Probiotic Propionibacterium jensenii 702 Supplementation on Layer Chicken Performance. Benef. Microbes 2010, 1, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Campaniello, D.; Bevilacqua, A.; Sinigaglia, M.; Altieri, C. Screening of Propionibacterium spp. for Potential Probiotic Properties. Anaerobe 2015, 34, 169–173. [Google Scholar] [CrossRef]
- Ibrahim, M.K.; Effat, B.A.M.; Tawfik, N.F.; Mehanna, N.S.; Soliman, N.R. Evaluation of probiotic potential of dairy propionibacteria. J. Innov. Pharm. Biol. Sci. JIPBS 2017, 4, 34–42. [Google Scholar]
- Barouei, J.; Moussavi, M.; Hodgson, D.M. Effect of Maternal Probiotic Intervention on HPA Axis, Immunity and Gut Microbiota in a Rat Model of Irritable Bowel Syndrome. PLoS ONE 2012, 7, e46051. [Google Scholar] [CrossRef]
- Eastwood, J.; van Hemert, S.; Poveda, C.; Elmore, S.; Williams, C.; Lamport, D.; Walton, G. The Effect of Probiotic Bacteria on Composition and Metabolite Production of Faecal Microbiota Using in Vitro Batch Cultures. Nutrients 2023, 15, 2563. [Google Scholar] [CrossRef]
- Rabah, H.; Ferret-Bernard, S.; Huang, S.; Le Normand, L.; Cousin, F.J.; Gaucher, F.; Jeantet, R.; Boudry, G.; Jan, G. The Cheese Matrix Modulates the Immunomodulatory Properties of Propionibacterium freudenreichii CIRM-BIA 129 in Healthy Piglets. Front. Microbiol. 2018, 9, 2584. [Google Scholar] [CrossRef]
- Wang, Y.; Su, M.A.; Wan, Y.Y. An Essential Role of the Transcription Factor GATA-3 for the Function of Regulatory T Cells. Immunity 2011, 35, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Rabah, H.; Ferret-Bernard, S.; Le Normand, L.; Gaucher, F.; Guerin, S.; Nogret, I.; Le Loir, Y.; Chen, X.D.; Jan, G.; et al. Propionic Fermentation by the Probiotic Propionibacterium freudenreichii to Functionalize Whey. J. Funct. Foods 2019, 52, 620–628. [Google Scholar] [CrossRef]
- Fong, F.L.Y.; El-Nezami, H.; Mykkänen, O.; Kirjavainen, P.V. The Effects of Single Strains and Mixtures of Probiotic Bacteria on Immune Profile in Liver, Spleen, and Peripheral Blood. Front. Nutr. 2022, 9, 773298. [Google Scholar] [CrossRef] [PubMed]
- Fukaya, T.; Murakami, R.; Takagi, H.; Sato, K.; Sato, Y.; Otsuka, H.; Ohno, M.; Hijikata, A.; Ohara, O.; Hikida, M.; et al. Conditional Ablation of CD205+ Conventional Dendritic Cells Impacts the Regulation of T-Cell Immunity and Homeostasis in Vivo. Proc. Natl. Acad. Sci. USA 2012, 109, 11288–11293. [Google Scholar] [CrossRef]
- Eicher, S.D.; Chitko-McKown, C.G.; Bryan, K.A. Variation in the Response of Bovine Alveolar Lavage Cells to Diverse Species of Probiotic Bacteria. BMC Res. Notes 2020, 13, 159. [Google Scholar] [CrossRef] [PubMed]
- Tinrat, S.; Jiraprasertwong, O. Isolation and Assessment of Probiotic Potential of Acidipropionibacterium acidipropionici C03B-STR from Goat Milk with Cholesterol-Lowering Capability. Folia Microbiol. 2024. [Google Scholar] [CrossRef]
- Onal Darilmaz, D.; Beyatli, Y. Bile Salt Deconjugation Activity of Propionibacterium Strains and Their Cholesterol Co-precipitation Abilities. Int. J. Dairy Technol. 2019, 72, 551–558. [Google Scholar] [CrossRef]
- Jan, G.; Belzacq, A.-S.; Haouzi, D.; Rouault, A.; Métivier, D.; Kroemer, G.; Brenner, C. Propionibacteria Induce Apoptosis of Colorectal Carcinoma Cells via Short-Chain Fatty Acids Acting on Mitochondria. Cell Death Differ. 2002, 9, 179–188. [Google Scholar] [CrossRef]
- Lan, A.; Lagadic-Gossmann, D.; Lemaire, C.; Brenner, C.; Jan, G. Acidic Extracellular pH Shifts Colorectal Cancer Cell Death from Apoptosis to Necrosis upon Exposure to Propionate and Acetate, Major End-Products of the Human Probiotic Propionibacteria. Apoptosis 2007, 12, 573–591. [Google Scholar] [CrossRef]
- Zárate, G.; Sáez, G.D.; Pérez Chaia, A. Dairy Propionibacteria Prevent the Proliferative Effect of Plant Lectins on SW480 Cells and Protect the Metabolic Activity of the Intestinal Microbiota In Vitro. Anaerobe 2017, 44, 58–65. [Google Scholar] [CrossRef]
- Cousin, F.J.; Jouan-Lanhouet, S.; Dimanche-Boitrel, M.-T.; Corcos, L.; Jan, G. Milk Fermented by Propionibacterium freudenreichii Induces Apoptosis of HGT-1 Human Gastric Cancer Cells. PLoS ONE 2012, 7, e31892. [Google Scholar] [CrossRef]
- Hatakka, K.; Holma, R.; El-Nezami, H.; Suomalainen, T.; Kuisma, M.; Saxelin, M.; Poussa, T.; Mykkänen, H.; Korpela, R. The Influence of Lactobacillus rhamnosus LC705 Together with Propionibacterium freudenreichii ssp. Shermanii JS on Potentially Carcinogenic Bacterial Activity in Human Colon. Int. J. Food Microbiol. 2008, 128, 406–410. [Google Scholar] [CrossRef]
- El-Nezami, H.S.; Polychronaki, N.N.; Ma, J.; Zhu, H.; Ling, W.; Salminen, E.K.; Juvonen, R.O.; Salminen, S.J.; Poussa, T.; Mykkänen, H.M. Probiotic Supplementation Reduces a Biomarker for Increased Risk of Liver Cancer in Young Men from Southern China. Am. J. Clin. Nutr. 2006, 83, 1199–1203. [Google Scholar] [CrossRef]
- El-Nezami, H.; Mykkanen, H.; Kankaanpaa, P.; Suomalainen, T.; Salminen, S.; Ahokas, J. Ability of a Mixture of Lactobacillus and Propionibacterium to Influence the Faecal Aflatoxin Content in Healthy Egyptian Volunteers: A Pilot Clinical Study. Biosci. Microflora 2000, 19, 41–45. [Google Scholar] [CrossRef]
- Vesterlund, S.; Karp, M.; Salminen, S.; Ouwehand, A.C. Staphylococcus aureus Adheres to Human Intestinal Mucus but Can Be Displaced by Certain Lactic Acid Bacteria. Microbiology 2006, 152 Pt 6, 1819–1826. [Google Scholar] [CrossRef]
- Hajfarajollah, H.; Mokhtarani, B.; Noghabi, K.A. Newly Antibacterial and Antiadhesive Lipopeptide Biosurfactant Secreted by a Probiotic Strain, Propionibacterium freudenreichii. Appl. Biochem. Biotechnol. 2014, 174, 2725–2740. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Kollanoor Johny, A. Characterizing the Antimicrobial Function of a Dairy-Originated Probiotic, Propionibacterium freudenreichii, against Multidrug-Resistant Salmonella Enterica Serovar Heidelberg in Turkey Poults. Front. Microbiol. 2018, 9, 1475. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Kollanoor-Johny, A. Effect of Propionibacterium freudenreichii on Salmonella Multiplication, Motility, and Association with Avian Epithelial Cells. Poult. Sci. 2017, 96, 1376–1386. [Google Scholar] [CrossRef]
- Zárate, G.; Palacios, J.M.; Villena, J.; Zúñiga-Hansen, M.E. Inhibition of Enteropathogens Adhesion to Human Enterocyte-like HT-29 Cells by a Dairy Strain of Propionibacterium acidipropionici. Benef. Microbes 2016, 7, 431–441. [Google Scholar] [CrossRef]
- Dyshlyuk, L.S.; Milentyeva, I.S.; Asyakina, L.K.; Ostroumov, L.A.; Osintsev, A.M.; Pozdnyakova, A.V. Using Bifidobacterium and Propionibacterium Strains in Probiotic Consortia to Normalize the Gastrointestinal Tract. Braz. J. Biol. 2022, 84, e256945. [Google Scholar] [CrossRef]
- Fijan, S.; Kocbek, P.; Steyer, A.; Vodičar, P.M.; Strauss, M. The Antimicrobial Effect of Various Single-Strain and Multi-Strain Probiotics, Dietary Supplements or Other Beneficial Microbes against Common Clinical Wound Pathogens. Microorganisms 2022, 10, 2518. [Google Scholar] [CrossRef]
- Savijoki, K.; San-Martin-Galindo, P.; Pitkänen, K.; Edelmann, M.; Sillanpää, A.; van der Velde, C.; Miettinen, I.; Patel, J.Z.; Yli-Kauhaluoma, J.; Parikka, M.; et al. Food-Grade Bacteria Combat Pathogens by Blocking AHL-Mediated Quorum Sensing and Biofilm Formation. Foods 2023, 12, 90. [Google Scholar] [CrossRef]
- Adams, M.C.; Huang, Y.; Kotula, L.; Blake, R.J.; Garg, M.L. The efficacy of a potential new probiotic, Propionibacterium jensenii 702, to correct vitamin B12 levels in an in vivo deficient animal model. Proc. Nutr. Soc. Aust. 2002, 26. [Google Scholar]
- Yajnik, C.; Kasture, S.; Kantikar, V.; Lubree, H.; Bhat, D.; Raut, D.; Memane, N.; Bhalerao, A.; Ladkat, R.; Yajnik, P.; et al. Efficacy of B12 Fortified Nutrient Bar and Yogurt in Improving Plasma B12 Concentrations-Results from 2 Double-Blind Randomized Placebo Controlled Trials. Food Nutr. Bull. 2021, 42, 480–489. [Google Scholar] [CrossRef]
- Zahed, O.; Khosravi-Darani, K.; Mortazavian Farsani, S.A.; Mohammadi, A. Bacterial Conjugated Linoleic Acid Bio-Fortification of Synbiotic Yogurts Using Propionibacterium freudenreichii as Adjunct Culture. Ital. J. Food Sci. 2021, 33, 1–11. [Google Scholar] [CrossRef]
- Zahed, O.; Khosravi-Darani, K.; Mortazavian, A.M.; Mohammadi, A. Effects of Cultivation Conditions on Biofortification of Yogurt with Natural Folate by Propionibacterium freudenreichii. Biocatal. Agric. Biotechnol. 2022, 39, 102267. [Google Scholar] [CrossRef]
- Kurmindla, H.K.; Chavannavar, S.V.; Bharathula, S.; Rayasandhra, A.U.; Deshpande, B. Vitamin B12 enriched milk based nutraceutical production using Propionibacterium freudenreichii. Pharma Innov. J. PIJ 2022, 11, 46–50. [Google Scholar]
- Zhang, Y.; Momoisea, P.; Lin, Q.; Liang, J.; Burrow, K.; Serventi, L. Evaluation of Sensory and Physicochemical Characteristics of Vitamin B12 Enriched Whole-Meal Sourdough Bread Fermented with Propionibacterium freudenreichii. Sustainability 2023, 15, 8157. [Google Scholar] [CrossRef]
- Tindjau, R.; Chua, J.-Y.; Liu, S.-Q. Utilization of Propionic Acid Bacteria in the Biotransformation of Soy (Tofu) Whey: Growth and Metabolite Changes. J. Food Sci. 2024, 89, 540–551. [Google Scholar] [CrossRef]
- Punniyamoorthy, S.; Subramani, T.; Shanmugavel, K.; Sampathrajan, V.; Thiyagamoorthy, U.M. Development of millet beverage for combating vitamin B12 deficiency in order to achieve food security. J. Food Sci. Technol. 2025. [Google Scholar] [CrossRef]
- Wu, X.; Liu, H.; Han, J.; Zhou, Z.; Chen, J.; Liu, X. Introducing Bacillus Natto and Propionibacterium shermanii into Soymilk Fermentation: A Promising Strategy for Quality Improvement and Bioactive Peptide Production during in Vitro Digestion. Food Chem. 2024, 455, 139585. [Google Scholar] [CrossRef]
- Loivamaa, I.; Greis, M.; Nikander, V.; Edelmann, M.; Pöysä, M.; Varmanen, P.; Saris, P.E.J. Two-Step Fermentation to Produce Vitamin B12 Containing Beer Using Propionibacterium freudenreichii and Yeast. Food Biosci. 2025, 63, 105807. [Google Scholar] [CrossRef]
- Sabater, C.; Fara, A.; Palacios, J.; Corzo, N.; Requena, T.; Montilla, A.; Zárate, G. Synthesis of Prebiotic Galactooligosaccharides from Lactose and Lactulose by Dairy Propionibacteria. Food Microbiol. 2019, 77, 93–105. [Google Scholar] [CrossRef]
- Kruk, M.; Varmanen, P.; Edelmann, M.; Chamlagain, B.; Trząskowska, M. Food By-Product Valorisation in Nutrients through Spent Brewer’s Yeast Bioprocessing with Propionibacterium freudenreichii. J. Clean. Prod. 2024, 434, 140102. [Google Scholar] [CrossRef]
- Meile, L.; Le Blay, G.; Thierry, A. Safety Assessment of Dairy Microorganisms: Propionibacterium and Bifidobacterium. Int. J. Food Microbiol. 2008, 126, 316–320. [Google Scholar] [CrossRef]
- Deptula, P.; Loivamaa, I.; Smolander, O.-P.; Laine, P.; Roberts, R.J.; Piironen, V.; Paulin, L.; Savijoki, K.; Auvinen, P.; Varmanen, P. Red-Brown Pigmentation of AcidiPropionibacterium jensenii Is Tied to Haemolytic Activity and cyl-Like Gene Cluster. Microorganisms 2019, 7, 512. [Google Scholar] [CrossRef]
- Uchida, M.; Yoda, N.; Terahara, M.; Seki, K.; Choi, S.S.H.; Roberts, A. Safety Evaluation of Propionibacterium freudenreichii ET-3 Culture. Regul. Toxicol. Pharmacol. 2011, 60, 249–261. [Google Scholar] [CrossRef]
- Uchida, M.; Tsuboi, H.; Takahashi Arita, M.; Nemoto, A.; Seki, K.; Tsunoo, H.; Martyres, S.; Roberts, A. Safety of High Doses of Propionibacterium freudenreichii ET-3 Culture in Healthy Adult Subjects. Regul. Toxicol. Pharmacol. 2011, 60, 262–267. [Google Scholar] [CrossRef]
- Tapiovaara, L.; Lehtoranta, L.; Poussa, T.; Mäkivuokko, H.; Korpela, R.; Pitkäranta, A. Absence of Adverse Events in Healthy Individuals Using Probiotics–Analysis of Six Randomised Studies by One Study Group. Benef. Microbes 2016, 7, 161–169. [Google Scholar] [CrossRef]
- Technical Committee: ISO/TC 34/SC 5 CS: 67.100.01. SO 10932:2010 | IDF 223:2010; Milk and Milk Products—Determination of the Minimal Inhibitory Concentration (MIC) of Antibiotics Applicable to Bifidobacteria and Non-Enterococcal Lactic Acid Bacteria (LAB). Edition 1. 2010. Available online: https://www.iso.org/standard/46434.html (accessed on 6 June 2025).
- European Food Safety Authority Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance. EFSA J. 2012, 10, 2740. [Google Scholar] [CrossRef]
- Babot, J.D.; Argañaraz-Martínez, E.; Saavedra, L.; Apella, M.C.; Chaia, A.P. Compatibility and Safety of Five Lectin-Binding Putative Probiotic Strains for the Development of a Multi-Strain Protective Culture for Poultry. Benef. Microbes 2018, 9, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Piwowarek, K.; Lipińska, E.; Hać-Szymańczuk, E.; Kieliszek, M.; Kot, A.M. Sequencing and Analysis of the Genome of Propionibacterium freudenreichii T82 Strain: Importance for Industry. Biomolecules 2020, 10, 348. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Amadoro, C.; Gasperi, M.; Colavita, G. Lactobacilli Infection Case Reports in the Last Three Years and Safety Implications. Nutrients 2022, 14, 1178. [Google Scholar] [CrossRef]
- Esaiassen, E.; Hjerde, E.; Cavanagh, J.P.; Simonsen, G.S.; Klingenberg, C. Bifidobacterium Bacteremia: Clinical Characteristics and a Genomic Approach to Assess Pathogenicity. J. Clin. Microbiol. 2017, 55, 2234–2248. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.L.; Moreira, J.; Lorenzo, A.; Lamas, C.C. Infectious Complications Following Probiotic Ingestion: A Potentially Underestimated Problem? A Systematic Review of Reports and Case Series. BMC Complement. Altern. Med. 2018, 18, 329. [Google Scholar] [CrossRef]
- Hwang, J.-B.; Jang, H.-J. Saccharomyces boulardii as a Single Trigger of Food Protein-Induced Enterocolitis Syndrome: Seven Case Reports. World J. Clin. Cases 2025, 13, 98111. [Google Scholar] [CrossRef]
- Tian, Q.; Ye, H.; Zhou, X.; Wang, J.; Zhang, L.; Sun, W.; Duan, C.; Fan, M.; Zhou, W.; Bi, C.; et al. Evaluating the Health Risk of Probiotic Supplements from the Perspective of Antimicrobial Resistance. Microbiol. Spectr. 2025, 13, e0001924. [Google Scholar] [CrossRef]
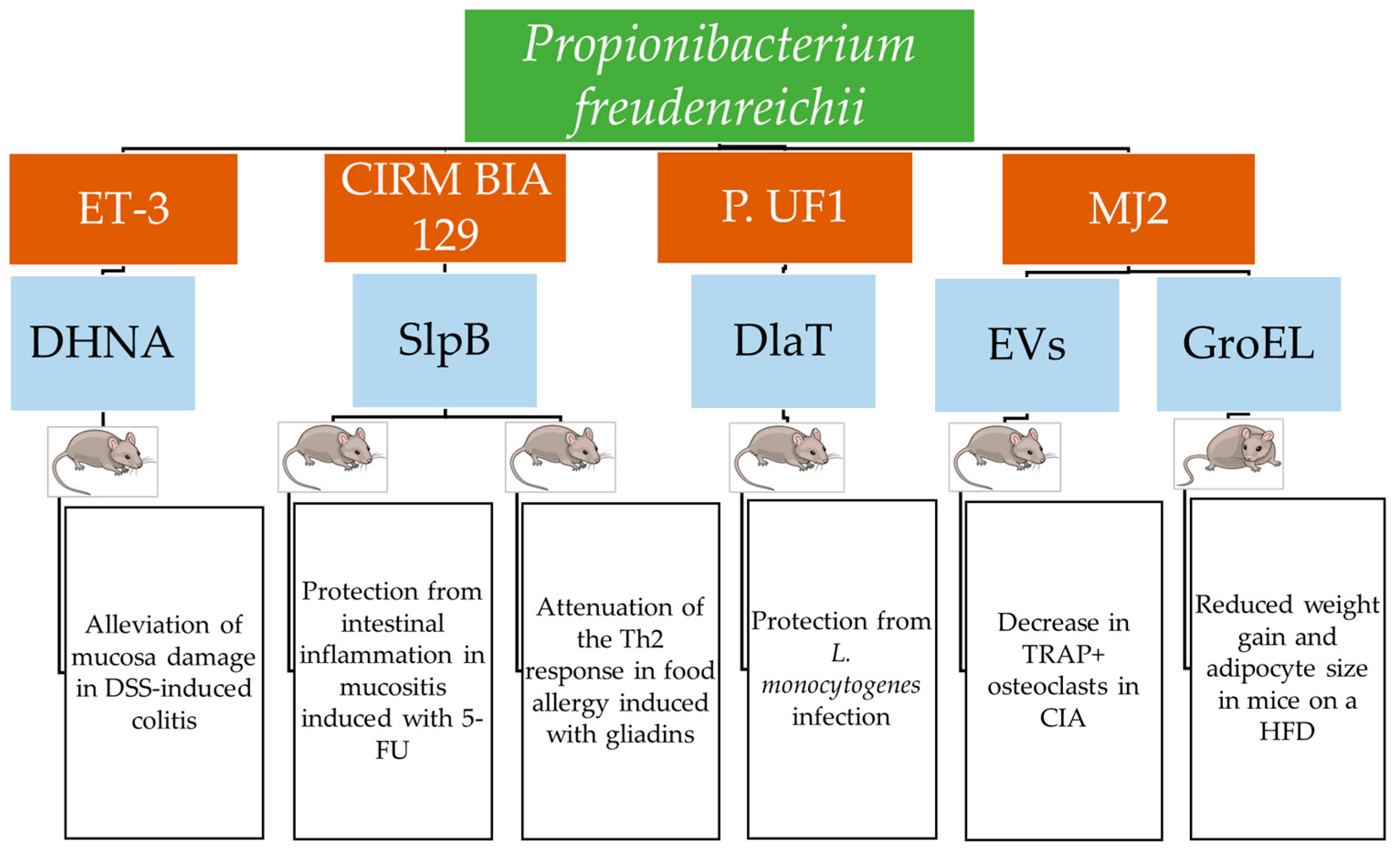
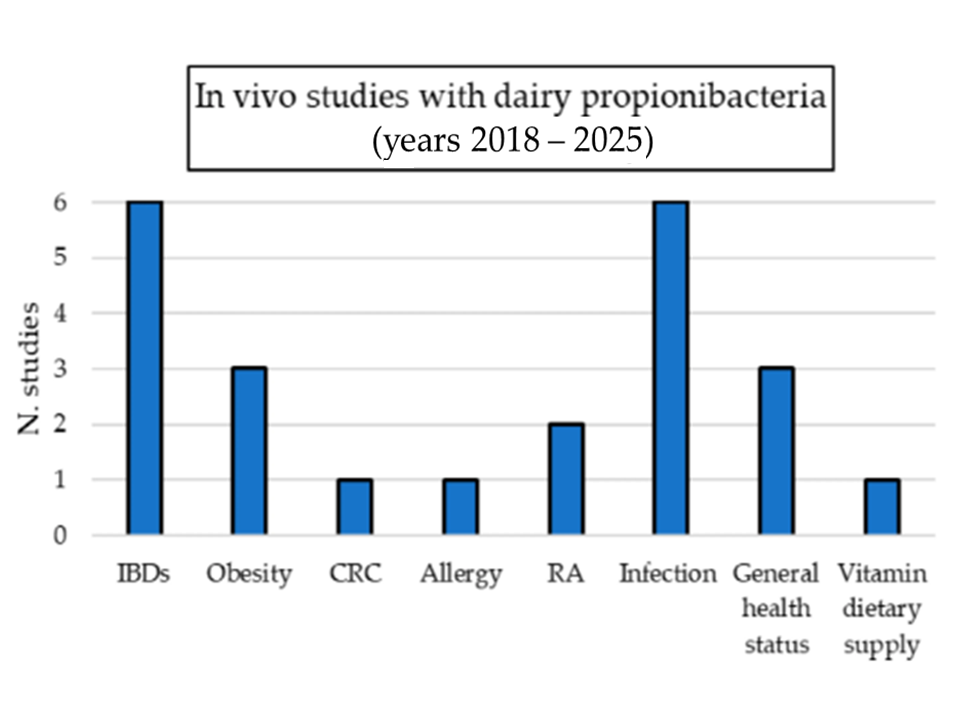

{kind=link}
{kind=link}
| Strain | Disease | Animal Model | Active Molecule | Induced Effects | Reference |
|---|---|---|---|---|---|
| P. freudenreichii ET-3 | TNBS-induced colitis | Rats | Propionate | Ulcer healing | [43] |
| P. freudenreichii ET-3 | DSS-induced colitis | Mice | DHNA | Reduction in molecules favoring leucocyte infiltration and vascular adhesion. Downregulation of IL-1β, IL-6, and TNF-α. Increase in lactobacilli and SCFAs. Activation of AhR-regulated pathways. Increase in colitis suppressing C-type RegIII lectins | [44,45] |
| P. freudenreichii CIRM BIA129 | TNBS-induced colitis | Mice | Cell surface proteins | DAI reduction. Attenuation of inflammation (increased expression of Pparγ) and oxidative stress (decreased expression of cox2 and Hmox). Restoration of intestinal barrier (increased expression of ZO-1). Decrease in IL-1β, IL-6, and IL-17 | [39,46,47,48,49] |
| P.freudenreichii KCTC 1063 | DSS-induced colitis | Rats | SCFAs | DAI reduction. Reduced crypt damage and leucocyte infiltration. Maintenance of mucin MUC2 expression level | [50] |
| P. freudenreichii B1 | DSS-induced colitis | Mice | Not investigated | Decrease in IL-8, IL-1β, and TNFα. Increase in ZO-1, claudin-1, and Rspo3. Downregulation of RHO kinase ROCK-1, and Axin2, which inactivate Wnt/β-catenin regeneration pathway. | [51] |
| P. freudenreichii CIRM BIA 129 | Mucositis induced with 5-FU | Mice | SlpB | Reduction in leucocyte infiltration and ulceration. Restored height of villi. Reduction in gut permeability (increased expression of cld1). Decreased expression of IL-17a, IL-12, and IL-1β | [52] |
| P. freudenreichii P.UF1 | NEC-like injury | Newborn mice | DlaT | Downregulation of nitric oxide synthase iNOS and interleukins Il-1b, Il-6, and Il-23. Increase in ILC3 expressing IL-17A and IL-22 | [14] |
| P. freudenreichii P.UF1 | Infection with L. monocytogenes | Mice | DlaT | Reduction in IL-1β, IL-6, and IL-12/IL23p40 produced by DCs. Reduction in Th1 cells producing IFNγ and increase in Th17 cells and IL-10+ Treg cells. | [14] |
| Strain | Disease | Animal Model | Active Molecule | Induced Effects | Reference |
|---|---|---|---|---|---|
| A. jensenii 702 | Administration of M. tuberculosis culture filtrate | Rats | Not defined | IFNγ levels 2 to 3 Log higher than IL-4, indicative of Th1 response effective in protection from tuberculosis | [54] |
| P. freudenreichii KCTC 1063 | No disease | C. elegans | Not defined | Upregulation of innate immunity-related pathways DBL/TGF-β, P38 MAPK, and Daf-2/DAF-16 involved in IIS; upregulation of antimicrobial peptide genes lys-7 and lys-8 | [55] |
| P. freudenreichii CIRM BIA 129 | Food allergy induced with wheat gliadins | Mice | SlpB | Prevention of body temperature increase; prevention of gliadin-specific IgE and IgG1 increase in serum; increase in gliadin-specific IgG2; prevention of intestinal permeabilization | [56] |
| Strain | Disease | Animal Model | Active Molecule | Induced effects | Reference |
|---|---|---|---|---|---|
| P. freudenreichii JS | HFD-induced obesity | ApoE*3 Leiden transgenic mice | Not investigated | Reduced weight gain and gonadal adipose tissue Decreased levels of VCAM-1 vascular inflammation marker Decreased mast cell number and TNF-α levels | [57] |
| P. freudenreichii MJ2 | HFD-induced obesity | Mice | GroEL | Reduced fat accumulation Pre-adipocyte stage maintenance by upregulation of Pref-1 and downregulation of PPARγ, C/EBPα, FAS, SCD-1, ACC, and lipolytic enzymes | [58,59] |
| A. acidipropionici OB7439 | HFD-induced obesity | Mice | Not investigated | Increased insulin secretion Decreased Tnfα, F4/80, ColIα, Fas, and Chrebp; increased Ppara in liver | [60] |
| Strain | Carcinogen | Animal Model | Induced Effects | Reference |
|---|---|---|---|---|
| P. freudenreichii DSM 20271 | AOM | Rats | Reduced formation of aberrant crypt foci | [61] |
| P. freudenreichii TL133 | DMH | Rats | Increased apoptosis and decreased proliferation of crypt cells | [89] |
| A. acidipropionici CRL 1198 | Concanavalin A | Mice | Reduced proliferation of intestinal epithelial cells; preserved microvilli structure | [90] |
| Strain | Disease | Animal Model | Active Molecule | Induced Effects | Reference |
|---|---|---|---|---|---|
| P. freudenreichii MJ2 | CIA | Mice | Surface protein extracts | Increased OPG/RANKL expression ratio | [64,65] |
| P. freudenreichii MJ2 | CIA | Mice | EVs | Decrease in IL-6, TNF-α and IL-17 Increase in IL-10 Increased OPG/RANKL expression ratio | [66] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, F.; Santonicola, S.; Giaccone, V.; Truant, A.; Colavita, G. Dairy Propionibacteria: Probiotic Properties and Their Molecular Bases. Biomolecules 2025, 15, 886. https://doi.org/10.3390/biom15060886
Rossi F, Santonicola S, Giaccone V, Truant A, Colavita G. Dairy Propionibacteria: Probiotic Properties and Their Molecular Bases. Biomolecules. 2025; 15(6):886. https://doi.org/10.3390/biom15060886
Chicago/Turabian StyleRossi, Franca, Serena Santonicola, Valerio Giaccone, Alessandro Truant, and Giampaolo Colavita. 2025. "Dairy Propionibacteria: Probiotic Properties and Their Molecular Bases" Biomolecules 15, no. 6: 886. https://doi.org/10.3390/biom15060886
APA StyleRossi, F., Santonicola, S., Giaccone, V., Truant, A., & Colavita, G. (2025). Dairy Propionibacteria: Probiotic Properties and Their Molecular Bases. Biomolecules, 15(6), 886. https://doi.org/10.3390/biom15060886