
Unravelling the Adiponectin Hallmark and Exploring the Therapeutic Potential of Its Receptor Agonists in Cancer Metabolic Reprogramming

 ,
,
Abstract

1. Introduction
2. Adiponectin
2.1. Physiological Role and Intracellular Signaling
2.2. Involvement in Malignant Diseases

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Cancer | Main Findings | Signaling Pathways | References |
|---|---|---|---|
| Breast | Adiponectin stimulates ERα+ MCF7 while inhibiting ERα− MDA-MB-231 cell proliferation | PI3K/AKT, MAPK | [76] |
| Lung | Adiponectin reduces viability and increases apoptosis in A549 cells | ERK1/2, AKT, CREB | [81,82] |
| Adiponectin inhibits migration and invasion in NSCLC cells | Twist | [83] | |
| Colorectal | Adiponectin represses colon cancer by inducing apoptosis | AMPK/mTOR, ERK1/2 | [84,85,86] |
| Adiponectin stimulates growth and inflammatory cytokine secretion in HT-29 cells | cAMP/PKA | [87] | |
| Adiponectin displays opposite effects in colorectal cancer depending on glucose availability | AMPK, PPARα, AKT | [88] | |
| Thyroid | Adiponectin inhibits proliferation and migration of human papillary thyroid cancer cells | ULK/LC-3 | [91] |
| Endometrial | Adiponectin impairs proliferation and induces apoptosis in human endometrial carcinoma cells | LKB1, AKT | [71,92] |
| Adiponectin deficiency promotes endometrial carcinogenesis in PTEN-null mice | ERK1/2 | [93] | |
| Prostate | Adiponectin inhibits VEGF-mediated cancer neovascularization | AMPK/mTOR/VEGF | [95] |
3. Adiponectin and Cell Metabolism
3.1. Glucose Metabolism
3.2. Lipid Metabolism
4. Adiponectin and Cancer Metabolism
4.1. Glucose Metabolism
4.2. Lipid Metabolism
4.3. Metabolic Oxidation and Oxidative Stress
5. Adiponectin Receptor Agonist AdipoRon in Cancer Metabolism
5.1. Glucose Metabolism
5.2. Lipid Metabolism
6. Other Adiponectin Receptor Agonists
7. Discussion
8. Future Directions
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| AICAR | 5-aminoimidazole-4-carboxamide-1-β-d-ribofuranoside |
| BMI | Body mass index |
| LMW | Low molecular weight |
| MMW | Medium molecular weight |
| HMW | High molecular weight |
| AdipoR1 | Adiponectin receptor 1 |
| AdipoR2 | Adiponectin receptor 2 |
| ECs | Extracellular domains |
| PPAR-α | Peroxisome proliferator-activated receptor alpha |
| APPL1/2 | Phosphotyrosine interacting with PH domain and leucine zipper 1/2 |
| IRS 1/2 | Insulin receptor substrate proteins 1/2 |
| PP2A | Protein phosphatase 2A |
| LKB1 | Liver kinase B1 |
| PKCζ | Protein kinase Cζ |
| CaMKK | Calmodulin-dependent protein kinase |
| PLC | Phospholipase C |
| IP3 | Inositol triphosphate |
| ER | Endoplasmic reticulum |
| SIRT1 | Sirtuin 1 |
| mTOR | Mammalian target of rapamycin |
| MAPK | Mitogen-activated protein kinase |
| PI3K | Phosphatidylinositol 3-kinase |
| AKT | Protein kinase B |
| NF-κB | Nuclear factor kappa B |
| JAK | Janus kinase |
| STAT | Signal transducer and activator of transcription |
| IκBα | Inhibitor of nuclear factor kappa B |
| PKA | Protein kinase A |
| NO | Nitric oxide |
| ROS | Reactive oxygen species |
| S1P | Sphingosine 1-phosphate |
| ERα | Estrogen receptor alpha |
| NSCLC | Non-small-cell lung cancer |
| CREB | Cyclic AMP-responsive element-binding protein |
| ERK 1/2 | Extracellular signal-regulated kinase 1/2 |
| RB | Retinoblastoma-associated protein |
| USP2 | Ubiquitin carboxyl-terminal hydrolase 2 |
| BCL-2 | B-cell leukemia/lymphoma 2 |
| PTEN | Phosphatase and tensin homolog |
| VEGF | Vascular endothelial growth factor |
| TCA | Tricarboxylic acid cycle |
| GLUT4 | Glucose transporter type 4 |
| CD36 | Cluster of differentiation 36 |
| UCP2 | Uncoupling protein 2 |
| HFD | High-fat diet |
| PGC-1α | Peroxisome proliferator-activated receptor gamma coactivator 1 alpha |
| ACC | Acetyl-CoA carboxylase |
| HNF | Hepatocyte nuclear factor |
| SREBP1 | Sterol regulatory element-binding protein 1 |
| ChREBP1 | Carbohydrate-responsive element-binding protein 1 |
| ABCA1 | ATP-binding cassette A1 |
| HDL | High-density lipoprotein |
| LDL | Low-density lipoprotein |
| LPL | Lipoprotein lipase |
| LDHA | Lactate dehydrogenase A |
| ALDOA | Aldolase A |
| PGK1 | Phosphoglycerate kinase 1 |
| PGAM1 | Phosphoglycerate mutase 1 |
| GLUT1 | Glucose transporter 1 |
| PDK1 | Pyruvate dehydrogenase kinase 1 |
| PDK | Pyruvate dehydrogenase complex |
| HIF-1α | Hypoxia-inducible factor 1α |
| FASN | Fatty acid synthase |
| ACLY | ATP citrate lyase |
| mTORC1 | Mammalian target of rapamycin complex 1 |
| OS | Oxidative stress |
| SOD | Superoxide dismutase |
| MnSOD | Manganese superoxide dismutase |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| NOX2 | NADPH oxidase 2 |
| PDAC | Pancreatic ductal adenocarcinoma |
| OCR | Oxygen consumption rate |
| ECAR | Extracellular acidification rate |
| PKM2 | Pyruvate kinase M2 |
| MCT | Monocarboxylate transporter |
| PNPLA2 | Adipose triglyceride lipase |
| GPAT1 | Glycerol-3-phosphate acyltransferase 1 |
| CPT1 | Carnitine palmitoyltransferase 1 |
| LPIN1 | Lipin-1 |
| LPA | Lysophosphatidic acid |
| G3P | Glyceraldehyde 3-phosphate |
| NASH | Non-alcoholic steatohepatitis |
| HSC | Hepatic stellate cell |
References
- Pati, S.; Irfan, W.; Jameel, A.; Ahmed, S.; Shahid, R.K. Obesity and Cancer: A Current Overview of Epidemiology, Pathogenesis, Outcomes, and Management. Cancers 2023, 15, 485. [Google Scholar] [CrossRef] [PubMed]
- Barberio, A.M.; Alareeki, A.; Viner, B.; Pader, J.; Vena, J.E.; Arora, P.; Friedenreich, C.M.; Brenner, D.R. Central body fatness is a stronger predictor of cancer risk than overall body size. Nat. Commun. 2019, 10, 383. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11, 1290. [Google Scholar] [CrossRef]
- Tilg, H.; Ianiro, G.; Gasbarrini, A.; Adolph, T.E. Adipokines: Masterminds of metabolic inflammation. Nat. Rev. Immunol. 2024, 25, 250–265. [Google Scholar] [CrossRef] [PubMed]
- Charles-Messance, H.; Mitchelson, K.A.J.; De Marco Castro, E.; Sheedy, F.J.; Roche, H.M. Regulating metabolic inflammation by nutritional modulation. J. Allergy Clin. Immunol. 2020, 146, 706–720. [Google Scholar] [CrossRef]
- Vitale, E.; Rizzo, A.; Santa, K.; Jirillo, E. Associations between “Cancer Risk”, “Inflammation” and “Metabolic Syndrome”: A Scoping Review. Biology 2024, 13, 352. [Google Scholar] [CrossRef]
- Shan, D.; Zhou, W.; Chang, Y.; Miao, M.; Zhou, W. From cancer inflammation to cancer metabolism: New targets for anticancer therapy. Precis. Nutr. 2024, 3, e00067. [Google Scholar] [CrossRef]
- Nong, S.; Han, X.; Xiang, Y.; Qian, Y.; Wei, Y.; Zhang, T.; Tian, K.; Shen, K.; Yang, J.; Ma, X. Metabolic reprogramming in cancer: Mechanisms and therapeutics. MedComm 2023, 4, e218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, S.; Wang, D.; Liu, S.; Xiao, T.; Gu, W.; Yang, H.; Wang, H.; Yang, M.; Chen, P. Metabolic reprogramming and immune evasion: The interplay in the tumor microenvironment. Biomark. Res. 2024, 12, 96. [Google Scholar] [CrossRef]
- Zheng, S.; Song, Q.; Zhang, P. Metabolic Modifications, Inflammation, and Cancer Immunotherapy. Front. Oncol. 2021, 11, 703681. [Google Scholar] [CrossRef]
- Tsankof, A.; Tziomalos, K. Adiponectin: A player in the pathogenesis of hormone-dependent cancers. Front. Endocrinol. 2022, 13, 1018515. [Google Scholar] [CrossRef] [PubMed]
- de Martino, M.; Leitner, C.V.; Hofbauer, S.L.; Lucca, I.; Haitel, A.; Shariat, S.F.; Klatte, T. Serum Adiponectin Predicts Cancer-specific Survival of Patients with Renal Cell Carcinoma. Eur. Urol. Focus 2016, 2, 197–203. [Google Scholar] [CrossRef]
- Ray, I.; Meira, L.B.; Michael, A.; Ellis, P.E. Adipocytokines and disease progression in endometrial cancer: A systematic review. Cancer Metastasis Rev. 2021, 41, 211–242. [Google Scholar] [CrossRef]
- Bocian-Jastrzębska, A.; Malczewska-Herman, A.; Kos-Kudła, B. Role of Leptin and Adiponectin in Carcinogenesis. Cancers 2023, 15, 4250. [Google Scholar] [CrossRef] [PubMed]
- Hebbard, L.; Ranscht, B. Multifaceted roles of Adiponectin in cancer. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 59–69. [Google Scholar] [CrossRef]
- Goel, S.; Singh, R.; Singh, V.; Singh, H.; Kumari, P.; Chopra, H.; Sharma, R.; Nepovimova, E.; Valis, M.; Kuca, K.; et al. Metformin: Activation of 5′ AMP-activated protein kinase and its emerging potential beyond anti-hyperglycemic action. Front. Genet. 2022, 13, 1022739. [Google Scholar] [CrossRef] [PubMed]
- Višnjić, D.; Lalić, H.; Dembitz, V.; Tomić, B.; Smoljo, T. AICAr, a Widely Used AMPK Activator with Important AMPK-Independent Effects: A Systematic Review. Cells 2021, 10, 1095. [Google Scholar] [CrossRef]
- Parker-Duffen, J.L.; Nakamura, K.; Silver, M.; Kikuchi, R.; Tigges, U.; Yoshida, S.; Denzel, M.S.; Ranscht, B.; Walsh, K. T-cadherin Is Essential for Adiponectin-mediated Revascularization. J Biol Chem 2013, 288, 24886–24897. [Google Scholar] [CrossRef]
- Lee, H.-P.; Lin, C.-Y.; Shih, J.-S.; Fong, Y.-C.; Wang, S.-W.; Li, T.-M.; Tang, C.-H. Adiponectin promotes VEGF-A-dependent angiogenesis in human chondrosarcoma through PI3K, Akt, mTOR, and HIF-α pathway. Oncotarget 2015, 6, 36746–36761. [Google Scholar] [CrossRef]
- Bråkenhielm, E.; Veitonmäki, N.; Cao, R.; Kihara, S.; Matsuzawa, Y.; Zhivotovsky, B.; Funahashi, T.; Cao, Y. Adiponectin-induced antiangiogenesis and antitumor activity involve caspase-mediated endothelial cell apoptosis. Proc. Natl. Acad. Sci. USA 2004, 101, 2476–2481. [Google Scholar] [CrossRef]
- Pham, D.V.; Park, P.H. Tumor Metabolic Reprogramming by Adipokines as a Critical Driver of Obesity-Associated Cancer Progression. Int. J. Mol. Sci. 2021, 22, 1444. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Daniele, A.; Salzillo, A.; Ragone, A.; Naviglio, S.; Sapio, L. AdipoRon and Other Adiponectin Receptor Agonists as Potential Candidates in Cancer Treatments. Int. J. Mol. Sci. 2021, 22, 5569. [Google Scholar] [CrossRef] [PubMed]
- Barbalho, S.M.; Méndez-Sánchez, N.; Fornari Laurindo, L. AdipoRon and ADP355, adiponectin receptor agonists, in Metabolic-associated Fatty Liver Disease (MAFLD) and Nonalcoholic Steatohepatitis (NASH): A systematic review. Biochem. Pharmacol. 2023, 218, 115871. [Google Scholar] [CrossRef]
- Akimoto, M.; Maruyama, R.; Kawabata, Y.; Tajima, Y.; Takenaga, K. Antidiabetic adiponectin receptor agonist AdipoRon suppresses tumour growth of pancreatic cancer by inducing RIPK1/ERK-dependent necroptosis. Cell Death Dis. 2018, 9, 804. [Google Scholar] [CrossRef]
- Malih, S.; Najafi, R. AdipoRon: A possible drug for colorectal cancer prevention? Tumor Biol. 2015, 36, 6673–6675. [Google Scholar] [CrossRef]
- Ramzan, A.A.; Bitler, B.G.; Hicks, D.; Barner, K.; Qamar, L.; Behbakht, K.; Powell, T.; Jansson, T.; Wilson, H. Adiponectin receptor agonist AdipoRon induces apoptotic cell death and suppresses proliferation in human ovarian cancer cells. Mol. Cell. Biochem. 2019, 461, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Manley, S.J.; Olou, A.A.; Jack, J.L.; Ruckert, M.T.; Walsh, R.M.; Eades, A.E.; Bye, B.A.; Ambrose, J.; Messaggio, F.; Anant, S.; et al. Synthetic adiponectin-receptor agonist, AdipoRon, induces glycolytic dependence in pancreatic cancer cells. Cell Death Dis. 2022, 13, 114. [Google Scholar] [CrossRef]
- Kafeel, S.; Ragone, A.; Salzillo, A.; Palmiero, G.; Naviglio, S.; Sapio, L. Adiponectin Receptor Agonist AdipoRon Inhibits Proliferation and Drives Glycolytic Dependence in Non-Small-Cell Lung Cancer Cells. Cancers 2024, 16, 2633. [Google Scholar] [CrossRef]
- Dubuisson, N.; Versele, R.; Davis-López de Carrizosa, M.A.; Selvais, C.M.; Noel, L.; Planchon, C.; Van den Bergh, P.Y.K.; Brichard, S.M.; Abou-Samra, M. The Adiponectin Receptor Agonist, ALY688: A Promising Therapeutic for Fibrosis in the Dystrophic Muscle. Cells 2023, 12, 2101. [Google Scholar] [CrossRef]
- Ma, O.K.-F.; Ronsisvalle, S.; Basile, L.; Xiang, A.W.; Tomasella, C.; Sipala, F.; Pappalardo, M.; Chan, K.-H.; Milardi, D.; Ng, R.C.-L.; et al. Identification of a novel adiponectin receptor and opioid receptor dual acting agonist as a potential treatment for diabetic neuropathy. Biomed. Pharmacother. 2023, 158, 114141. [Google Scholar] [CrossRef]
- Straub, L.G.; Scherer, P.E. Metabolic Messengers: Adiponectin. Nat. Metab. 2019, 1, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.V.; Scherer, P.E. Adiponectin, the past two decades. J. Mol. Cell Biol. 2016, 8, 93–100. [Google Scholar] [CrossRef]
- Sayeed, M.; Gautam, S.; Verma, D.P.; Afshan, T.; Kumari, T.; Srivastava, A.K.; Ghosh, J.K. A collagen domain-derived short adiponectin peptide activates APPL1 and AMPK signaling pathways and improves glucose and fatty acid metabolisms. J. Biol. Chem. 2018, 293, 13509–13523. [Google Scholar] [CrossRef] [PubMed]
- Tumminia, A.; Vinciguerra, F.; Parisi, M.; Graziano, M.; Sciacca, L.; Baratta, R.; Frittitta, L. Adipose Tissue, Obesity and Adiponectin: Role in Endocrine Cancer Risk. Int. J. Mol. Sci. 2019, 20, 2863. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, D.; Vatner, D.F.; Goedeke, L.; Hirabara, S.M.; Zhang, Y.; Perry, R.J.; Shulman, G.I. Mechanisms by which adiponectin reverses high fat diet-induced insulin resistance in mice. Proc. Natl. Acad. Sci. USA 2020, 117, 32584–32593. [Google Scholar] [CrossRef]
- Tanabe, H.; Fujii, Y.; Okada-Iwabu, M.; Iwabu, M.; Nakamura, Y.; Hosaka, T.; Motoyama, K.; Ikeda, M.; Wakiyama, M.; Terada, T.; et al. Crystal structures of the human adiponectin receptors. Nature 2015, 520, 312–316. [Google Scholar] [CrossRef]
- Chang, M.L.; Yang, Z.E.; Yang, S.S. Roles of Adipokines in Digestive Diseases: Markers of Inflammation, Metabolic Alteration and Disease Progression. Int. J. Mol. Sci. 2020, 21, 8308. [Google Scholar] [CrossRef]
- Muratore, M.; Komai, A.M. Theoretical study of the adiponectin receptors: Binding site characterization and molecular dynamics of possible ligands for drug design. SN Appl. Sci. 2020, 2, 533. [Google Scholar] [CrossRef]
- Pilon, M. Paradigm shift: The primary function of the “Adiponectin Receptors” is to regulate cell membrane composition. Lipids Health Dis. 2021, 20, 43. [Google Scholar] [CrossRef] [PubMed]
- Obata, Y.; Kita, S.; Koyama, Y.; Fukuda, S.; Takeda, H.; Takahashi, M.; Fujishima, Y.; Nagao, H.; Masuda, S.; Tanaka, Y.; et al. Adiponectin/T-cadherin system enhances exosome biogenesis and decreases cellular ceramides by exosomal release. JCI Insight 2018, 3, e99680. [Google Scholar] [CrossRef]
- Rubina, K.A.; Semina, E.V.; Kalinina, N.I.; Sysoeva, V.Y.; Balatskiy, A.V.; Tkachuk, V.A. Revisiting the multiple roles of T-cadherin in health and disease. Eur. J. Cell Biol. 2021, 100, 151183. [Google Scholar] [CrossRef]
- Denzel, M.S.; Scimia, M.-C.; Zumstein, P.M.; Walsh, K.; Ruiz-Lozano, P.; Ranscht, B. T-cadherin is critical for adiponectin-mediated cardioprotection in mice. J. Clin. Investig. 2010, 120, 4342–4352. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D. Adiponectin: Role in physiology and pathophysiology. Int. J. Prev. Med. 2020, 11, 136. [Google Scholar] [CrossRef]
- Roy, B.; Palaniyandi, S.S. Tissue-specific role and associated downstream signaling pathways of adiponectin. Cell Biosci. 2021, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Liu, M. Adipose tissue in control of metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef]
- Nagaraju, G.P.; Rajitha, B.; Aliya, S.; Kotipatruni, R.P.; Madanraj, A.S.; Hammond, A.; Park, D.; Chigurupati, S.; Alam, A.; Pattnaik, S. The role of adiponectin in obesity-associated female-specific carcinogenesis. Cytokine Growth Factor Rev. 2016, 31, 37–48. [Google Scholar] [CrossRef]
- Liu, Z.; Xiao, T.; Peng, X.; Li, G.; Hu, F. APPLs: More than just adiponectin receptor binding proteins. Cell. Signal. 2017, 32, 76–84. [Google Scholar] [CrossRef]
- Shi, P.; Tian, Y.; Xu, F.; Liu, L.-N.; Wu, W.-H.; Shi, Y.-Z.; Dai, A.-Q.; Fang, H.-Y.; Li, K.-X.; Xu, C. Assessment of pathogenicity and functional characterization of APPL1 gene mutations in diabetic patients. World J. Diabetes 2024, 15, 275–286. [Google Scholar] [CrossRef]
- Deepa, S.S.; Zhou, L.; Ryu, J.; Wang, C.; Mao, X.; Li, C.; Zhang, N.; Musi, N.; DeFronzo, R.A.; Liu, F.; et al. APPL1 Mediates Adiponectin-Induced LKB1 Cytosolic Localization Through the PP2A-PKCζ Signaling Pathway. Mol. Endocrinol. 2011, 25, 1773–1785. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Deepa, S.S.; Etzler, J.C.; Ryu, J.; Mao, X.; Fang, Q.; Liu, D.D.; Torres, J.M.; Jia, W.; Lechleiter, J.D.; et al. Adiponectin Activates AMP-activated Protein Kinase in Muscle Cells via APPL1/LKB1-dependent and Phospholipase C/Ca2+/Ca2+/Calmodulin-dependent Protein Kinase Kinase-dependent Pathways. J. Biol. Chem. 2009, 284, 22426–22435. [Google Scholar] [CrossRef] [PubMed]
- Grossini, E.; Farruggio, S.; Qoqaiche, F.; Raina, G.; Camillo, L.; Sigaudo, L.; Mary, D.; Surico, N.; Surico, D. Monomeric adiponectin modulates nitric oxide release and calcium movements in porcine aortic endothelial cells in normal/high glucose conditions. Life Sci. 2016, 161, 1–9. [Google Scholar] [CrossRef]
- Göransson, O.; Kopietz, F.; Rider, M.H. Metabolic control by AMPK in white adipose tissue. Trends Endocrinol. Metab. 2023, 34, 704–717. [Google Scholar] [CrossRef]
- Hardie, D.G.; Schaffer, B.E.; Brunet, A. AMPK: An Energy-Sensing Pathway with Multiple Inputs and Outputs. Trends Cell Biol. 2016, 26, 190–201. [Google Scholar] [CrossRef]
- Hadley, J.T.; Ryu, J.; Dong, L.Q. Adiponectin and Adiponectin Signaling. In Cellular Endocrinology in Health and Disease; Academic Press: Cambridge, MA, USA, 2021; pp. 261–287. [Google Scholar]
- Fernández, G.; de Fernández, C.A.; Amores, F.B.C.; Arráiz, G.F.; Cabrera, M. Adiponectin: Ceramidase activity and stress control of the endoplasmic reticle in obesity. Rev. Latinoam. Hiperte. 2023, 18, 170–178. [Google Scholar] [CrossRef]
- Choi, C.H.J.; Cohen, P. Adipose crosstalk with other cell types in health and disease. Exp. Cell Res. 2017, 360, 6–11. [Google Scholar] [CrossRef]
- Di Zazzo, E.; Polito, R.; Bartollino, S.; Nigro, E.; Porcile, C.; Bianco, A.; Daniele, A.; Moncharmont, B. Adiponectin as Link Factor between Adipose Tissue and Cancer. Int. J. Mol. Sci. 2019, 20, 839. [Google Scholar] [CrossRef] [PubMed]
- Macis, D.; Guerrieri-Gonzaga, A.; Gandini, S. Circulating adiponectin and breast cancer risk: A systematic review and meta-analysis. Int. J. Epidemiol. 2014, 43, 1226–1236. [Google Scholar] [CrossRef]
- Ye, J.J.; Jia, J.; Dong, S.J.; Zhang, C.L.; Yu, S.Q.; Li, L.X.; Mao, C.M.; Wang, D.; Chen, J.J.; Yuan, G.Y. Circulating adiponectin levels and the risk of breast cancer: A meta-analysis. Eur. J. Cancer Prev. 2014, 23, 158–165. [Google Scholar] [CrossRef]
- Petridou, E.T.; Mitsiades, N.; Gialamas, S.; Angelopoulos, M.; Skalkidou, A.; Dessypris, N.; Hsi, A.; Lazaris, N.; Polyzos, A.; Syrigos, C.; et al. Circulating adiponectin levels and expression of adiponectin receptors in relation to lung cancer: Two case-control studies. Oncology 2007, 73, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Perrotta, F.; Monaco, M.L.; Polito, R.; Pafundi, P.C.; Matera, M.G.; Daniele, A.; Bianco, A. Implications of the Adiponectin System in Non-Small Cell Lung Cancer Patients: A Case-Control Study. Biomolecules 2020, 10, 926. [Google Scholar] [CrossRef] [PubMed]
- Karapanagiotou, E.M.; Tsochatzis, E.A.; Dilana, K.D.; Tourkantonis, L.; Gratsias, I.; Syrigos, K.N. The significance of leptin, adiponectin, and resistin serum levels in non-small cell lung cancer (NSCLC). Lung Cancer 2008, 61, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Macleod, A.; Scheurlen, K.M.; Burton, J.F.; Parks, M.A.; Sumy, M.S.A.; Gaskins, J.T.; Galandiuk, S. Systemic adiponectin levels in colorectal cancer and adenoma: A systematic review and meta-analysis. Int. J. Obes. 2023, 47, 911–921. [Google Scholar] [CrossRef]
- Polito, R.; Nigro, E.; Fei, L.; De Magistris, L.; Monaco, M.L.; D’Amico, R.; Naviglio, S.; Signoriello, G.; Daniele, A. Adiponectin Is Inversely Associated With Tumour Grade in Colorectal Cancer Patients. Anticancer Res. 2020, 40, 3751–3757. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Fu, X.; Li, J.; Liu, L.; Alkohlani, A.; Tan, S.C.; Low, T.Y.; Hou, Y. Association of circulating leptin and adiponectin levels with colorectal cancer risk: A systematic review and meta-analysis of case-control studies. Cancer Epidemiol. 2021, 73, 101958. [Google Scholar] [CrossRef]
- Dossus, L.; Franceschi, S.; Biessy, C.; Navionis, A.S.; Travis, R.C.; Weiderpass, E.; Scalbert, A.; Romieu, I.; Tjonneland, A.; Olsen, A.; et al. Adipokines and inflammation markers and risk of differentiated thyroid carcinoma: The EPIC study. Int. J. Cancer 2018, 142, 1332–1342. [Google Scholar] [CrossRef]
- Abooshahab, R.; Yaghmaei, P.; Ghadaksaz, H.G.; Hedayati, M. Lack of Association between Serum Adiponectin/Leptin Levels and Medullary Thyroid Cancer. Asian Pac. J. Cancer Prev. 2016, 17, 3861–3864. [Google Scholar]
- Li, R.; Dong, F.; Zhang, L.; Ni, X.; Lin, G. Role of adipocytokines in endometrial cancer progression. Front. Pharmacol. 2022, 13, 1090227. [Google Scholar] [CrossRef]
- Slabuszewska-Józwiak, A.; Lukaszuk, A.; Janicka-Kosnik, M.; Wdowiak, A.; Jakiel, G. Role of Leptin and Adiponectin in Endometrial Cancer. Int. J. Mol. Sci. 2022, 23, 5307. [Google Scholar] [CrossRef]
- Soliman, P.T.; Cui, X.; Zhang, Q.; Hankinson, S.E.; Lu, K.H. Circulating adiponectin levels and risk of endometrial cancer: The prospective Nurses’ Health Study. Am. J. Obstet. Gynecol. 2011, 204, e161–e167. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.B.; Xu, H.; Hu, J.M.; Zhu, W.H.; Yang, T.; Jiang, H.W.; Ding, Q. Genetic polymorphisms in leptin, adiponectin and their receptors affect risk and aggressiveness of prostate cancer: Evidence from a meta-analysis and pooled-review. Oncotarget 2016, 7, 81049–81061. [Google Scholar] [CrossRef]
- Burton, A.J.; Gilbert, R.; Tilling, K.; Langdon, R.; Donovan, J.L.; Holly, J.M.P.; Martin, R.M. Circulating adiponectin and leptin and risk of overall and aggressive prostate cancer: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 320. [Google Scholar] [CrossRef]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and cancer risk: Emerging biological mechanisms and perspectives. Metabolism 2019, 92, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Mauro, L.; Pellegrino, M.; De Amicis, F.; Ricchio, E.; Giordano, F.; Rizza, P.; Catalano, S.; Bonofiglio, D.; Sisci, D.; Panno, M.L.; et al. Evidences that estrogen receptor α interferes with adiponectin effects on breast cancer cell growth. Cell Cycle 2014, 13, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Otvos, L.; Knappe, D.; Hoffmann, R.; Kovalszky, I.; Olah, J.; Hewitson, T.D.; Stawikowska, R.; Stawikowski, M.; Cudic, P.; Lin, F.; et al. Development of second generation peptides modulating cellular adiponectin receptor responses. Front. Chem. 2014, 2, 93. [Google Scholar] [CrossRef]
- Naimo, G.D.; Gelsomino, L.; Catalano, S.; Mauro, L.; Andò, S. Interfering Role of ERα on Adiponectin Action in Breast Cancer. Front. Endocrinol. 2020, 11, 66. [Google Scholar] [CrossRef]
- Mauro, L.; Naimo, G.D.; Ricchio, E.; Panno, M.L.; Andò, S. Cross-talk between adiponectin and IGF-IR in breast cancer. Front. Oncol. 2015, 5, 157. [Google Scholar] [CrossRef]
- Denzel, M.S.; Hebbard, L.W.; Shostak, G.; Shapiro, L.; Cardiff, R.D.; Ranscht, B. Adiponectin Deficiency Limits Tumor Vascularization in the MMTV-PyV-mT Mouse Model of Mammary Cancer. Clin. Cancer Res. 2009, 15, 3256–3264. [Google Scholar] [CrossRef]
- Nigro, E.; Scudiero, O.; Sarnataro, D.; Mazzarella, G.; Sofia, M.; Bianco, A.; Daniele, A. Adiponectin affects lung epithelial A549 cell viability counteracting TNFa and IL-1ß toxicity through AdipoR1. Int. J. Biochem. Cell Biol. 2013, 45, 1145–1153. [Google Scholar] [CrossRef]
- Illiano, M.; Nigro, E.; Sapio, L.; Caiafa, I.; Spina, A.; Scudiero, O.; Bianco, A.; Esposito, S.; Mazzeo, F.; Pedone, P.V.; et al. Adiponectin down-regulates CREB and inhibits proliferation of A549 lung cancer cells. Pulm. Pharmacol. Ther. 2017, 45, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Cui, E.; Guo, H.; Shen, M.; Yu, H.; Gu, D.; Mao, W.; Wang, X. Adiponectin inhibits migration and invasion by reversing epithelial-mesenchymal transition in non-small cell lung carcinoma. Oncol. Rep. 2018, 40, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, M.; Takahashi, H.; Hosono, K.; Endo, H.; Kato, S.; Yoneda, K.; Nozaki, Y.; Fujita, K.; Yoneda, M.; Wada, K.; et al. Adiponectin inhibits colorectal cancer cell growth through the AMPK/mTOR pathway. Int. J. Oncol. 2009, 34, 339–344. [Google Scholar] [CrossRef]
- Kim, A.Y.; Lee, Y.S.; Kim, K.H.; Lee, J.H.; Lee, H.K.; Jang, S.H.; Kim, S.E.; Lee, G.Y.; Lee, J.W.; Jung, S.A.; et al. Adiponectin Represses Colon Cancer Cell Proliferation via AdipoR1-and-R2-Mediated AMPK Activation. Mol. Endocrinol. 2010, 24, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Schettino, P.; Polito, R.; Scudiero, O.; Monaco, M.L.; De Palma, G.D.; Daniele, A. Adiponectin and colon cancer: Evidence for inhibitory effects on viability and migration of human colorectal cell lines. Mol. Cell. Biochem. 2018, 448, 125–135. [Google Scholar] [CrossRef]
- Ogunwobi, O.O.; Beales, I.L.P. Adiponectin stimulates proliferation and cytokine secretion in colonic epithelial cells. Regul Pept. 2006, 134, 105–113. [Google Scholar] [CrossRef]
- Habeeb, B.S.; Kitayama, J.; Nagawa, H. Adiponectin supports cell survival in glucose deprivation through enhancement of autophagic response in colorectal cancer cells. Cancer Sci. 2011, 102, 999–1006. [Google Scholar] [CrossRef]
- Mitsiades, N.; Pazaitou-Panayiotou, K.; Aronis, K.N.; Moon, H.S.; Chamberland, J.P.; Liu, X.W.; Diakopoulos, K.N.; Kyttaris, V.; Panagiotou, V.; Mylvaganam, G.; et al. Circulating Adiponectin Is Inversely Associated with Risk of Thyroid Cancer: In vivo and in vitro studies. J. Clin. Endocr. Metab. 2011, 96, E2023–E2028. [Google Scholar] [CrossRef]
- Zhou, Y.Y.; Yang, Y.; Zhou, T.C.; Li, B.; Wang, Z.J. Adiponectin and Thyroid Cancer: Insight into the Association between Adiponectin and Obesity. Aging Dis. 2021, 12, 597–613. [Google Scholar] [CrossRef]
- Li, C.L.; Zhang, J.; Dionigi, G.; Liang, N.; Guan, H.X.; Sun, H. Adiponectin Inhibits the Progression of Obesity-Associated Papillary Thyroid Carcinoma Through Autophagy. Endocrinology 2024, 165, bqae030. [Google Scholar] [CrossRef]
- Cong, L.; Gasser, J.; Zhao, J.; Yang, B.; Li, F.; Zhao, A.Z. Human adiponectin inhibits cell growth and induces apoptosis in human endometrial carcinoma cells, HEC-1-A and RL95-2. Endocr.-Relat. Cancer 2007, 14, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.J.; Shi, H.; Zhao, Z.G.; Wang, S.; Zhou, S.J.; Mu, Y.P.; Ding, N.; Lai, Y.M.; Zhao, A.Z.; Cheng, L.X.; et al. Adiponectin deficiency promotes endometrial carcinoma pathogenesis and development via activation of mitogen-activated protein kinase. J. Pathol. 2022, 257, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Philp, L.K.; Rockstroh, A.; Lehman, M.; Sadowski, M.C.; Bartonicek, N.; Wade, J.D.; Otvos, L.; Nelson, C.C. Adiponectin receptor activation inhibits prostate cancer xenograft growth. Endocr.-Relat. Cancer 2020, 27, 711–729. [Google Scholar] [CrossRef]
- Gao, Q.R.; Zheng, J.H.; Yao, X.D.; Peng, B. Adiponectin inhibits VEGF-A in prostate cancer cells. Tumor Biol. 2015, 36, 4287–4292. [Google Scholar] [CrossRef]
- Li, Y.; Onodera, T.; Scherer, P.E. Adiponectin. Trends Endocrinol. Metab. 2024, 35, 674–675. [Google Scholar] [CrossRef] [PubMed]
- Field, B.C.; Gordillo, R.; Scherer, P.E. The Role of Ceramides in Diabetes and Cardiovascular Disease Regulation of Ceramides by Adipokines. Front. Endocrinol. 2020, 11, 569250. [Google Scholar] [CrossRef]
- Entezari, M.; Hashemi, D.; Taheriazam, A.; Zabolian, A.; Mohammadi, S.; Fakhri, F.; Hashemi, M.; Hushmandi, K.; Ashrafizadeh, M.; Zarrabi, A.; et al. AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: A pre-clinical and clinical investigation. Biomed. Pharmacother. 2022, 146, 112563. [Google Scholar] [CrossRef]
- Ghadge, A.A.; Khaire, A.A.; Kuvalekar, A.A. Adiponectin: A potential therapeutic target for metabolic syndrome. Cytokine Growth Factor Rev. 2018, 39, 151–158. [Google Scholar] [CrossRef]
- Begum, M.; Choubey, M.; Tirumalasetty, M.B.; Arbee, S.; Mohib, M.M.; Wahiduzzaman, M.; Mamun, M.A.; Uddin, M.B.; Mohiuddin, M.S. Adiponectin: A Promising Target for the Treatment of Diabetes and Its Complications. Life 2023, 13, 2213. [Google Scholar] [CrossRef]
- Krause, M.P.; Milne, K.J.; Hawke, T.J. Adiponectin—Consideration for its Role in Skeletal Muscle Health. Int. J. Mol. Sci. 2019, 20, 1528. [Google Scholar] [CrossRef]
- Ryu, J.; Hadley, J.T.; Li, Z.; Dong, F.; Xu, H.; Xin, X.; Zhang, Y.; Chen, C.; Li, S.; Guo, X.; et al. Adiponectin Alleviates Diet-Induced Inflammation in the Liver by Suppressing MCP-1 Expression and Macrophage Infiltration. Diabetes 2021, 70, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, S.; Aiello, G.; Mansilla Di Martino, E.; Campaci, D.; Muthanna, F.M.S.; Lombardo, M. The Role of Adipose Tissue and Nutrition in the Regulation of Adiponectin. Nutrients 2024, 16, 2436. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Yuan, B.B.; Lo, K.A.; Patterson, H.C.; Sun, Y.T.; Lodish, H.F. Adiponectin regulates expression of hepatic genes critical for glucose and lipid metabolism. Proc. Natl. Acad. Sci. USA 2012, 109, 14568–14573. [Google Scholar] [CrossRef]
- Ceddia, R.B.; Somwar, R.; Maida, A.; Fang, X.; Bikopoulos, G.; Sweeney, G. Globular adiponectin increases GLUT4 translocation and glucose uptake but reduces glycogen synthesis in rat skeletal muscle cells. Diabetologia 2004, 48, 132–139. [Google Scholar] [CrossRef]
- Richter, E.A.; Hargreaves, M. Exercise, GLUT4, and Skeletal Muscle Glucose Uptake. Physiol. Rev. 2013, 93, 993–1017. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Dong, L.Q. Adiponectin signaling and function in insulin target tissues. J. Mol. Cell Biol. 2016, 8, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Martínez Báez, A.; Ayala, G.; Pedroza-Saavedra, A.; González-Sánchez, H.M.; Chihu Amparan, L. Phosphorylation Codes in IRS-1 and IRS-2 Are Associated with the Activation/Inhibition of Insulin Canonical Signaling Pathways. Curr. Issues Mol. Biol. 2024, 46, 634–649. [Google Scholar] [CrossRef]
- Su, J.; Tang, L.; Luo, Y.; Xu, J.; Ouyang, S. Research progress on drugs for diabetes based on insulin receptor/insulin receptor substrate. Biochem. Pharmacol. 2023, 217, 115830. [Google Scholar] [CrossRef]
- Awazawa, M.; Ueki, K.; Inabe, K.; Yamauchi, T.; Kubota, N.; Kaneko, K.; Kobayashi, M.; Iwane, A.; Sasako, T.; Okazaki, Y.; et al. Adiponectin Enhances Insulin Sensitivity by Increasing Hepatic IRS-2 Expression via a Macrophage-Derived IL-6-Dependent Pathway. Cell Metab. 2011, 13, 401–412. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef]
- Onodera, T.; Wang, M.-Y.; Rutkowski, J.M.; Deja, S.; Chen, S.; Balzer, M.S.; Kim, D.-S.; Sun, X.; An, Y.A.; Field, B.C.; et al. Endogenous renal adiponectin drives gluconeogenesis through enhancing pyruvate and fatty acid utilization. Nat. Commun. 2023, 14, 6531. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.H.; Combs, T.P.; Du, X.; Brownlee, M.; Scherer, P.E. The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat. Med. 2001, 7, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Duval, F.; Dos Santos, E.; Maury, B.; Serazin, V.; Fathallah, K.; Vialard, F.; Dieudonné, M.-N. Adiponectin regulates glycogen metabolism at the human fetal–maternal interface. J. Mol. Endocrinol. 2018, 61, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Talley, J.T.; Mohiuddin, S.S. Biochemistry, Fatty Acid Oxidation. In StatPearls; Disclosure: Shamim Mohiuddin Declares no Relevant Financial Relationships with Ineligible Companies; Ineligible Companies: Treasure Island, FL, USA, 2025. [Google Scholar]
- Merritt, J.L.; MacLeod, E.; Jurecka, A.; Hainline, B. Clinical manifestations and management of fatty acid oxidation disorders. Rev. Endocr. Metab. Disord. 2020, 21, 479–493. [Google Scholar] [CrossRef]
- Samovski, D.; Jacome-Sosa, M.; Abumrad, N.A. Fatty Acid Transport and Signaling: Mechanisms and Physiological Implications. Annu. Rev. Physiol. 2023, 85, 317–337. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, S.; Li, Z. Adipokines in glucose and lipid metabolism. Adipocyte 2023, 12, 2202976. [Google Scholar] [CrossRef]
- Esfahani, M.; Movahedian, A.; Baranchi, M.; Goodarzi, M.T. Adiponectin: An adipokine with protective features against metabolic syndrome. Iran. J. Basic Med. Sci. 2015, 18, 430–442. [Google Scholar]
- Yoon, M.J.; Lee, G.Y.; Chung, J.J.; Ahn, Y.H.; Hong, S.H.; Kim, J.B. Adiponectin increases fatty acid oxidation in skeletal muscle cells by sequential activation of AMP-activated protein kinase, p38 mitogen-activated protein kinase, and peroxisome proliferator-activated receptor α. Diabetes 2006, 55, 2562–2570. [Google Scholar] [CrossRef]
- Abou-Samra, M.; Selvais, C.M.; Dubuisson, N.; Brichard, S.M. Adiponectin and Its Mimics on Skeletal Muscle: Insulin Sensitizers, Fat Burners, Exercise Mimickers, Muscling Pills … or Everything Together? Int. J. Mol. Sci. 2020, 21, 2620. [Google Scholar] [CrossRef]
- Otu, L.I.; Otu, A. Adiponectin and the Control of Metabolic Dysfunction: Is Exercise the Magic Bullet? Front. Physiol. 2021, 12, 651732. [Google Scholar] [CrossRef]
- Hayhurst, G.P.; Lee, Y.H.; Lambert, G.; Ward, J.M.; Gonzalez, F.J. Hepatocyte nuclear factor 4α (nuclear receptor 2A1) is essential for maintenance of hepatic gene expression and lipid homeostasis. Mol. Cell Biol. 2001, 21, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Sifuentes, A.; Holland, W.L. Regulation of glucose and lipid homeostasis by adiponectin: Effects on hepatocytes, pancreatic β cells and adipocytes. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhao, Z.-W.; Zeng, P.-H.; Zhou, Y.-J.; Yin, W.-J. Molecular mechanisms for ABCA1-mediated cholesterol efflux. Cell Cycle 2022, 21, 1121–1139. [Google Scholar] [CrossRef]
- Matsuura, F.; Oku, H.; Koseki, M.; Sandoval, J.C.; Yuasa-Kawase, M.; Tsubakio-Yamamoto, K.; Masuda, D.; Maeda, N.; Tsujii, K.I.; Ishigami, M.; et al. Adiponectin accelerates reverse cholesterol transport by increasing high density lipoprotein assembly in the liver. Biochem. Biophys. Res. Commun. 2007, 358, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- van Stijn, C.M.; Kim, J.; Barish, G.D.; Tietge, U.J.; Tangirala, R.K. Adiponectin expression protects against angiotensin II-mediated inflammation and accelerated atherosclerosis. PLoS ONE 2014, 9, e86404. [Google Scholar] [CrossRef]
- Okamoto, Y.; Kihara, S.; Funahashi, T.; Matsuzawa, Y.; Libby, P. Adiponectin: A key adipocytokine in metabolic syndrome. Clin. Sci. 2006, 110, 267–278. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Zhu, J.J.; Thompson, C.B. The hallmarks of cancer metabolism: Still emerging. Cell Metab. 2022, 34, 355–377. [Google Scholar] [CrossRef]
- Paul, S.; Ghosh, S.; Kumar, S. Tumor glycolysis, an essential sweet tooth of tumor cells. Semin. Cancer Biol. 2022, 86, 1216–1230. [Google Scholar] [CrossRef]
- Barba, I.; Carrillo-Bosch, L.; Seoane, J. Targeting the Warburg Effect in Cancer: Where Do We Stand? Int. J. Mol. Sci. 2024, 25, 3142. [Google Scholar] [CrossRef]
- Scheid, M.P.; Sweeney, G. The role of adiponectin signaling in metabolic syndrome and cancer. Rev. Endocr. Metab. Disord. 2013, 15, 157–167. [Google Scholar] [CrossRef]
- Yang, S.; Sun, Y.; Guo, Y.; Zhao, Z.; Hu, F.; Cong, L. The glycolysis-related AMPK/ULK signaling pathway mediates the inhibitory effect of adiponectin in prostate cancer cells. Mol. Cell. Endocrinol. 2024, 593, 112338. [Google Scholar] [CrossRef] [PubMed]
- Claps, G.; Faouzi, S.; Quidville, V.; Chehade, F.; Shen, S.; Vagner, S.; Robert, C. The multiple roles of LDH in cancer. Nat. Rev. Clin. Oncol. 2022, 19, 749–762. [Google Scholar] [CrossRef]
- Hou, X.M.; Yuan, S.Q.; Zhao, D.; Liu, X.J.; Wu, X.A. LDH-A promotes malignant behavior via activation of epithelial-to-mesenchymal transition in lung adenocarcinoma (volume 39, pg 631, 2019). Biosci. Rep. 2020, 40, BSR20181476. [Google Scholar] [CrossRef]
- Wang, H.Q.; Zhou, R.; Sun, L.; Xia, J.L.; Yang, X.C.; Pan, C.Q.; Huang, N.; Shi, M.; Bin, J.P.; Liao, Y.L.; et al. deficiency promotes GC invasion and migration via the enhancements of LDHA expression and aerobic glycolysis. Endocr.-Relat. Cancer 2017, 24, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Hanai, J.I.; Ren, J.G.; Kats, L.; Burgess, K.; Bhargava, P.; Signoretti, S.; Billiard, J.; Duffy, K.J.; Grant, A.; et al. Targeting Lactate Dehydrogenase-A Inhibits Tumorigenesis and Tumor Progression in Mouse Models of Lung Cancer and Impacts Tumor-Initiating Cells. Cell Metab. 2014, 19, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, M.; Bartoloni, B.; Cantini, G.; Nencioni, E.; Magherini, F.; Luconi, M.; Modesti, A.; Gamberi, T.; Fiaschi, T. STAT3 Signalling Drives LDH Up-Regulation and Adiponectin Down-Regulation in Cachectic Adipocytes. Int. J. Mol. Sci. 2023, 24, 6343. [Google Scholar] [CrossRef]
- Zheng, N.; Wei, J.; Wu, D.; Xu, Y.; Guo, J. Master kinase PDK1 in tumorigenesis. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2023, 1878, 188971. [Google Scholar] [CrossRef]
- Parida, S.; Siddharth, S.; Sharma, D. Adiponectin, Obesity, and Cancer: Clash of the Bigwigs in Health and Disease. Int. J. Mol. Sci. 2019, 20, 2519. [Google Scholar] [CrossRef]
- Faubert, B.; Boily, G.; Izreig, S.; Griss, T.; Samborska, B.; Dong, Z.F.; Dupuy, F.; Chambers, C.; Fuerth, B.J.; Viollet, B.; et al. AMPK Is a Negative Regulator of the Warburg Effect and Suppresses Tumor Growth In Vivo. Cell Metab. 2013, 17, 113–124. [Google Scholar] [CrossRef]
- Koundouros, N.; Poulogiannis, G. Reprogramming of fatty acid metabolism in cancer. Br. J. Cancer 2020, 122, 4–22. [Google Scholar] [CrossRef]
- Liu, J.; Xu, A.; Lam, K.S.-L.; Wong, N.-S.; Chen, J.; Shepherd, P.R.; Wang, Y. Cholesterol-induced mammary tumorigenesis is enhanced by adiponectin deficiency: Role of LDL receptor upregulation. Oncotarget 2013, 4, 1804–1818. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.; Godlewski, G.; Earley, B.J.; Zhou, L.; Jourdan, T.; Szanda, G.; Cinar, R.; Kunos, G. Role of adiponectin in the metabolic effects of cannabinoid type 1 receptor blockade in mice with diet-induced obesity. Am. J. Physiol. -Endocrinol. Metab. 2014, 306, E457–E468. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.-W.; Lee, S.; Chun, Y.-S. How cancer cells remodel lipid metabolism: Strategies targeting transcription factors. Lipids Health Dis. 2021, 20, 163. [Google Scholar] [CrossRef]
- Chung, S.J.; Nagaraju, G.P.; Nagalingam, A.; Muniraj, N.; Kuppusamy, P.; Walker, A.; Woo, J.; Győrffy, B.; Gabrielson, E.; Saxena, N.K.; et al. ADIPOQ/adiponectin induces cytotoxic autophagy in breast cancer cells through STK11/LKB1-mediated activation of the AMPK-ULK1 axis. Autophagy 2017, 13, 1386–1403. [Google Scholar] [CrossRef]
- Pham, D.-V.; Park, P.-H. Adiponectin triggers breast cancer cell death via fatty acid metabolic reprogramming. J. Exp. Clin. Cancer Res. 2022, 41, 9. [Google Scholar] [CrossRef]
- Chen, X.-W.; Don, A.S.; Fedorova, M.; Harayama, T.; Krahmer, N.; Nagata, S.; Narayan, P.; Nicholas, D.; Nowinski, S.M.; Saheki, Y.; et al. Time for lipid cell biology. Nat. Cell Biol. 2025, 27, 169–174. [Google Scholar] [CrossRef]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xie, C.; Fan, X.-X.; Jiang, Z.-B.; Wong, V.K.-W.; Xu, J.-H.; Yao, X.-J.; Liu, L.; Leung, E.L.-H. Novel direct AMPK activator suppresses non-small cell lung cancer through inhibition of lipid metabolism. Oncotarget 2017, 8, 96089–96102. [Google Scholar] [CrossRef]
- Hosios, A.M.; Wilkinson, M.E.; McNamara, M.C.; Kalafut, K.C.; Torrence, M.E.; Asara, J.M.; Manning, B.D. mTORC1 regulates a lysosome-dependent adaptive shift in intracellular lipid species. Nat. Metab. 2022, 4, 1792–1811. [Google Scholar] [CrossRef]
- Porporato, P.E.; Filigheddu, N.; Bravo-San Pedro, J.M.; Kroemer, G.; Galluzzi, L. Mitochondrial metabolism and cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef]
- Punter, K.B.; Chu, C.; Chan, E.Y.W. Mitochondrial dynamics and oxidative phosphorylation as critical targets in cancer. Endocr.-Relat. Cancer 2023, 30, e220229. [Google Scholar] [CrossRef]
- Uslu, C.; Kapan, E.; Lyakhovich, A. Cancer resistance and metastasis are maintained through oxidative phosphorylation. Cancer Lett. 2024, 587, 216705. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Sulejczak, D.; Kleczkowska, P.; Bukowska-Ośko, I.; Kucia, M.; Popiel, M.; Wietrak, E.; Kramkowski, K.; Wrzosek, K.; Kaczyńska, K. Mitochondrial Oxidative Stress—A Causative Factor and Therapeutic Target in Many Diseases. Int. J. Mol. Sci. 2021, 22, 3384. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Iqbal, M.J.; Kabeer, A.; Abbas, Z.; Siddiqui, H.A.; Calina, D.; Sharifi-Rad, J.; Cho, W.C. Interplay of oxidative stress, cellular communication and signaling pathways in cancer. Cell Commun. Signal. 2024, 22, 7. [Google Scholar] [CrossRef]
- McDowell, J.A.; Kosmacek, E.A.; Baine, M.J.; Adebisi, O.; Zheng, C.; Bierman, M.M.; Myers, M.S.; Chatterjee, A.; Liermann-Wooldrik, K.T.; Lim, A.; et al. Exogenous APN protects normal tissues from radiation-induced oxidative damage and fibrosis in mice and prostate cancer patients with higher levels of APN have less radiation-induced toxicities. Redox Biol. 2024, 73. [Google Scholar] [CrossRef]
- Lin, H.; Yu, C.H.; Jen, C.Y.; Cheng, C.F.; Chou, Y.; Chang, C.C.; Juan, S.H. Adiponectin-Mediated Heme Oxygenase-1 Induction Protects Against Iron-Induced Liver Injury via a PPARα-Dependent Mechanism. Am. J. Pathol. 2010, 177, 1697–1709. [Google Scholar] [CrossRef]
- Kim, M.J.; Nagy, L.E.; Park, P.H. Globular Adiponectin Inhibits Ethanol-Induced Reactive Oxygen Species Production through Modulation of NADPH Oxidase in Macrophages: Involvement of Liver Kinase B1/AMP-Activated Protein Kinase Pathway. Mol. Pharmacol. 2014, 86, 284–296. [Google Scholar] [CrossRef]
- Ren, Y.H.; Li, Y.; Yan, J.; Ma, M.K.; Zhou, D.M.; Xue, Z.Y.; Zhang, Z.M.; Liu, H.K.; Yang, H.P.; Jia, L.; et al. Adiponectin modulates oxidative stress-induced mitophagy and protects C2C12 myoblasts against apoptosis. Sci. Rep. 2017, 7, 3209. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.P.; Hou, Z.F.; Duivenvoorden, W.C.; Whelan, K.; Honig, A.; Pinthus, J.H. Adiponectin inhibits oxidative stress in human prostate carcinoma cells. Prostate Cancer Prostatic Dis. 2012, 15, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Okada-Iwabu, M.; Yamauchi, T.; Iwabu, M.; Honma, T.; Hamagami, K.; Matsuda, K.; Yamaguchi, M.; Tanabe, H.; Kimura-Someya, T.; Shirouzu, M.; et al. A small-molecule AdipoR agonist for type 2 diabetes and short life in obesity. Nature 2013, 503, 493. [Google Scholar] [CrossRef]
- Messaggio, F.; Mendonsa, A.M.; Castellanos, J.; Nagathihalli, N.S.; Gorden, L.; Merchant, N.B.; VanSaun, M.N. Adiponectin receptor agonists inhibit leptin induced pSTAT3 and in vivo pancreatic tumor growth. Oncotarget 2017, 8, 85378–85391. [Google Scholar] [CrossRef]
- Sapio, L.; Nigro, E.; Ragone, A.; Salzillo, A.; Illiano, M.; Spina, A.; Polito, R.; Daniele, A.; Naviglio, S. AdipoRon Affects Cell Cycle Progression and Inhibits Proliferation in Human Osteosarcoma Cells. J. Oncol. 2020, 2020, 7262479. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Dionigi, G.; Liang, N.; Guan, H.; Sun, H. Uncovering the connection between obesity and thyroid cancer: The therapeutic potential of adiponectin receptor agonist in the AdipoR2-ULK axis. Cell Death Dis. 2024, 15, 708. [Google Scholar] [CrossRef]
- Ragone, A.; Salzillo, A.; Spina, A.; Naviglio, S.; Sapio, L. Integrating Gemcitabine-Based Therapy With AdipoRon Enhances Growth Inhibition in Human PDAC Cell Lines. Front. Pharmacol. 2022, 13, 837503. [Google Scholar] [CrossRef]
- Sapio, L.; Ragone, A.; Spina, A.; Salzillo, A.; Naviglio, S. AdipoRon and Pancreatic Ductal Adenocarcinoma: A future perspective in overcoming chemotherapy-induced resistance? Cancer Drug Resist. 2022, 5, 625–636. [Google Scholar] [CrossRef]
- Kafeel, S.; Palmiero, G.; Salzillo, A.; Ragone, A.; Naviglio, S.; Sapio, L. Combining AdipoRon with Paclitaxel Unveils Synergistic Potential in Non-Small Cell Lung Cancer Cells via AMPK-ERK1/2 Signaling. Cells 2025, 14, 602. [Google Scholar] [CrossRef]
- Massart, I.S.; Kouakou, A.N.; Pelet, N.; Lause, P.; Schakman, O.; Loumaye, A.; Abou-Samra, M.; Deldicque, L.; Bindels, L.B.; Brichard, S.M.; et al. Administration of adiponectin receptor agonist AdipoRon relieves cancer cachexia by mitigating inflammation in tumour-bearing mice. J. Cachexia Sarcopeni 2024, 15, 919–933. [Google Scholar] [CrossRef]
- Salinas, M.L.; Fuentes, N.R.; Choate, R.; Wright, R.C.; McMurray, D.N.; Chapkin, R.S. AdipoRon Attenuates Wnt Signaling by Reducing Cholesterol-Dependent Plasma Membrane Rigidity. Biophys. J. 2020, 118, 885–897. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, Q.; Song, N.; Yan, Z.; Lin, R.; Wu, S.; Jiang, L.; Hong, S.; Xie, J.; Zhou, H.; et al. AdipoR1/AdipoR2 dual agonist recovers nonalcoholic steatohepatitis and related fibrosis via endoplasmic reticulum-mitochondria axis. Nat. Commun. 2020, 11, 5807. [Google Scholar] [CrossRef]
- Goto, T.; Teraminami, A.; Lee, J.-Y.; Ohyama, K.; Funakoshi, K.; Kim, Y.-I.; Hirai, S.; Uemura, T.; Yu, R.; Takahashi, N.; et al. Tiliroside, a glycosidic flavonoid, ameliorates obesity-induced metabolic disorders via activation of adiponectin signaling followed by enhancement of fatty acid oxidation in liver and skeletal muscle in obese–diabetic mice. J. Nutr. Biochem. 2012, 23, 768–776. [Google Scholar] [CrossRef]
- Bourouh, M.; Marignani, P.A. The Tumor Suppressor Kinase LKB1: Metabolic Nexus. Front. Cell Dev. Biol. 2022, 10, 881297. [Google Scholar] [CrossRef]
- Trelford, C.B.; Shepherd, T.G. Insights into targeting LKB1 in tumorigenesis. Genes Dis. 2025, 12, 101402. [Google Scholar] [CrossRef]
- Saxena, N.K.; Sharma, D. Metastasis suppression by adiponectin: LKB1 rises up to the challenge. Cell Adhes. Migr. 2010, 4, 358–362. [Google Scholar] [CrossRef]
- Xia, J.Y.; Sun, K.; Hepler, C.; Ghaben, A.L.; Gupta, R.K.; An, Y.A.; Holland, W.L.; Morley, T.S.; Adams, A.C.; Gordillo, R.; et al. Acute loss of adipose tissue-derived adiponectin triggers immediate metabolic deterioration in mice. Diabetologia 2017, 61, 932–941. [Google Scholar] [CrossRef]
- Cai, L.; Ying, M.; Wu, H. Microenvironmental Factors Modulating Tumor Lipid Metabolism: Paving the Way to Better Antitumoral Therapy. Front. Oncol. 2021, 11, 777273. [Google Scholar] [CrossRef]
- Peretti, D.; Kim, S.; Tufi, R.; Lev, S. Lipid Transfer Proteins and Membrane Contact Sites in Human Cancer. Front. Cell Dev. Biol. 2020, 7, 371. [Google Scholar] [CrossRef]
- Nong, H.; Song, X.; Li, Y.; Xu, Y.; Wang, F.; Wang, Y.; Zhang, J.; Chen, C.; Li, J. AdipoRon reduces cisplatin-induced ototoxicity in hair cells:possible relation to the regulation of mitochondrial biogenesis. Neurosci. Lett. 2024, 819, 137577. [Google Scholar] [CrossRef]
- Salmons, H.I.; Gow, C.; Limberg, A.K.; Bettencourt, J.W.; Carstens, M.F.; Payne, A.N.; Morrey, M.E.; Sanchez-Sotelo, J.; Berry, D.J.; Dudakovic, A.; et al. The Safety of Adiponectin Receptor Agonist AdipoRon in a Rabbit Model of Arthrofibrosis. Tissue Eng. Part C Methods 2023, 29, 154–159. [Google Scholar] [CrossRef]
- Sall, K.; Foulks, G.; Pflugfelder, S.C.; Evans, D.; Shettle, P.; Stewart, J.; Hsu, H. A Phase 1/2a Study of ALY688 Ophthalmic Solution in Dry Eye Subjects. Investig. Ophthalmol. Vis. Sci. 2023, 64, 3959. [Google Scholar]
- Xiao, Y.; Yu, T.-J.; Xu, Y.; Ding, R.; Wang, Y.-P.; Jiang, Y.-Z.; Shao, Z.-M. Emerging therapies in cancer metabolism. Cell Metab. 2023, 35, 1283–1303. [Google Scholar] [CrossRef]
- Lord, S.R.; Harris, A.L. Is it still worth pursuing the repurposing of metformin as a cancer therapeutic? Br. J. Cancer 2023, 128, 958–966. [Google Scholar] [CrossRef]
- Zhu, L.; Yang, K.; Ren, Z.; Yin, D.; Zhou, Y. Metformin as anticancer agent and adjuvant in cancer combination therapy: Current progress and future prospect. Transl. Oncol. 2024, 44, 101945. [Google Scholar] [CrossRef]
- Masuo, H.; Kubota, K.; Shimizu, A.; Notake, T.; Miyazaki, S.; Yoshizawa, T.; Sakai, H.; Hayashi, H.; Soejima, Y. Increased mitochondria are responsible for the acquisition of gemcitabine resistance in pancreatic cancer cell lines. Cancer Sci 2023, 114, 4388–4400. [Google Scholar] [CrossRef]
- Dettoni, R.; Bahamondes, C.; Yevenes, C.; Cespedes, C.; Espinosa, J. The effect of obesity on chronic diseases in USA: A flexible copula approach. Sci. Rep. 2023, 13, 1831. [Google Scholar] [CrossRef]
- Olateju, I.V.; Opaleye-Enakhimion, T.; Udeogu, J.E.; Asuquo, J.; Olaleye, K.T.; Osa, E.; Oladunjoye, A.F. A systematic review on the effectiveness of diet and exercise in the management of obesity. Diabetes Metab. Syndr. Clin. Res. Rev. 2023, 17, 102759. [Google Scholar] [CrossRef]
- Janiszewska, J.; Ostrowska, J.; Szostak-Węgierek, D. The Influence of Nutrition on Adiponectin—A Narrative Review. Nutrients 2021, 13, 1394. [Google Scholar] [CrossRef]
- Sanchis, P.; Calvo, P.; Pujol, A.; Rivera, R.; Berga, F.; Fortuny, R.; Costa-Bauza, A.; Grases, F.; Masmiquel, L. Daily phytate intake increases adiponectin levels among patients with diabetes type 2: A randomized crossover trial. Nutr. Diabetes 2023, 13, 2. [Google Scholar] [CrossRef]
- García-Hermoso, A.; Ceballos-Ceballos, R.J.M.; Poblete-Aro, C.E.; Hackney, A.C.; Mota, J.; Ramírez-Vélez, R. Exercise, adipokines and pediatric obesity: A meta-analysis of randomized controlled trials. Int. J. Obes. 2016, 41, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Becic, T.; Studenik, C.; Hoffmann, G. Exercise Increases Adiponectin and Reduces Leptin Levels in Prediabetic and Diabetic Individuals: Systematic Review and Meta-Analysis of Randomized Controlled Trials. Med. Sci. 2018, 6, 97. [Google Scholar] [CrossRef]
- Mallardo, M.; D’Alleva, M.; Lazzer, S.; Giovanelli, N.; Graniero, F.; Billat, V.; Fiori, F.; Marinoni, M.; Parpinel, M.; Daniele, A.; et al. Improvement of adiponectin in relation to physical performance and body composition in young obese males subjected to twenty-four weeks of training programs. Heliyon 2023, 9, e15790. [Google Scholar] [CrossRef]
- Zaidi, H.; Byrkjeland, R.; Njerve, I.U.; Åkra, S.; Solheim, S.; Arnesen, H.; Seljeflot, I.; Opstad, T.B. Adiponectin in relation to exercise and physical performance in patients with type 2 diabetes and coronary artery disease. Adipocyte 2021, 10, 612–620. [Google Scholar] [CrossRef]
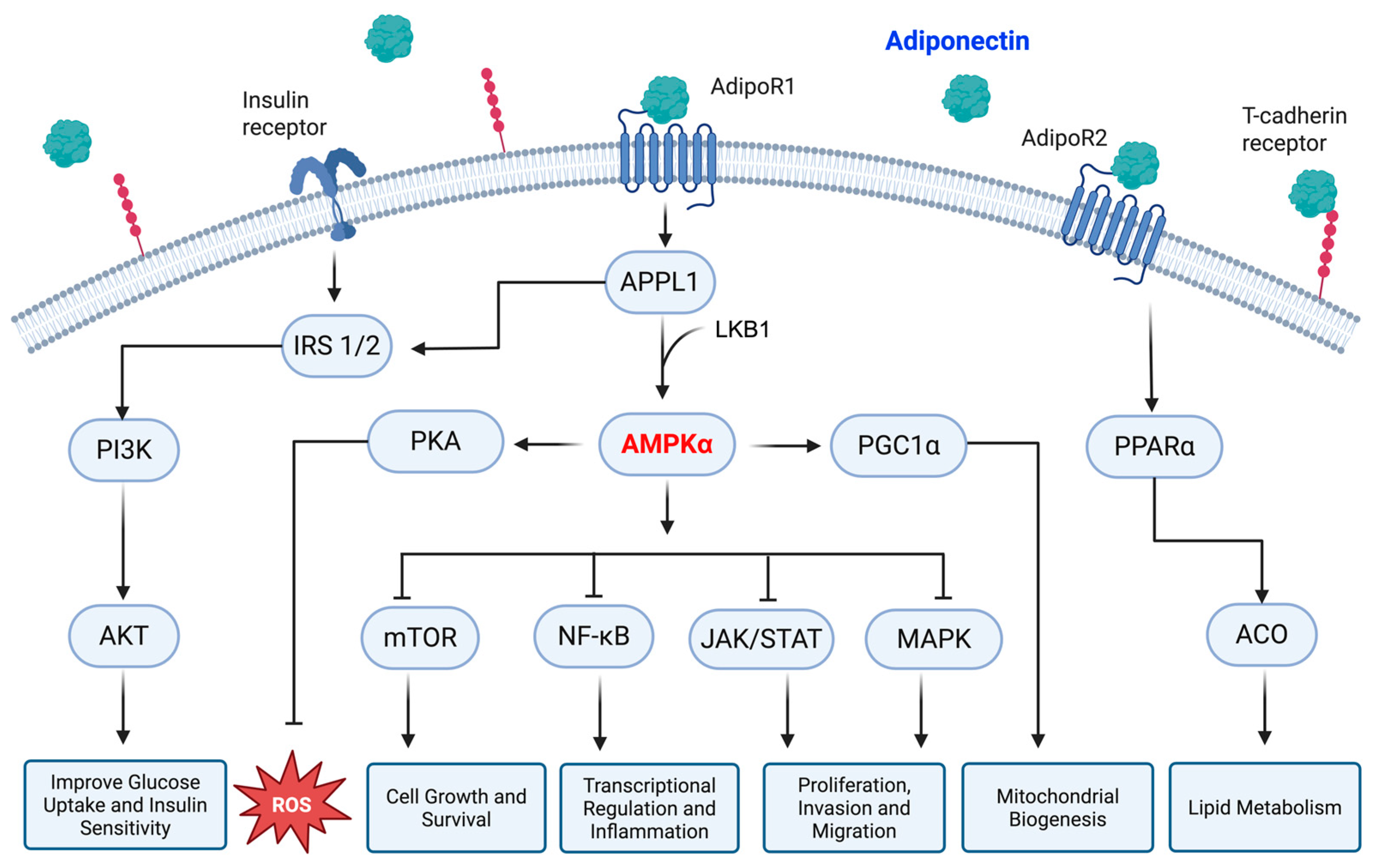
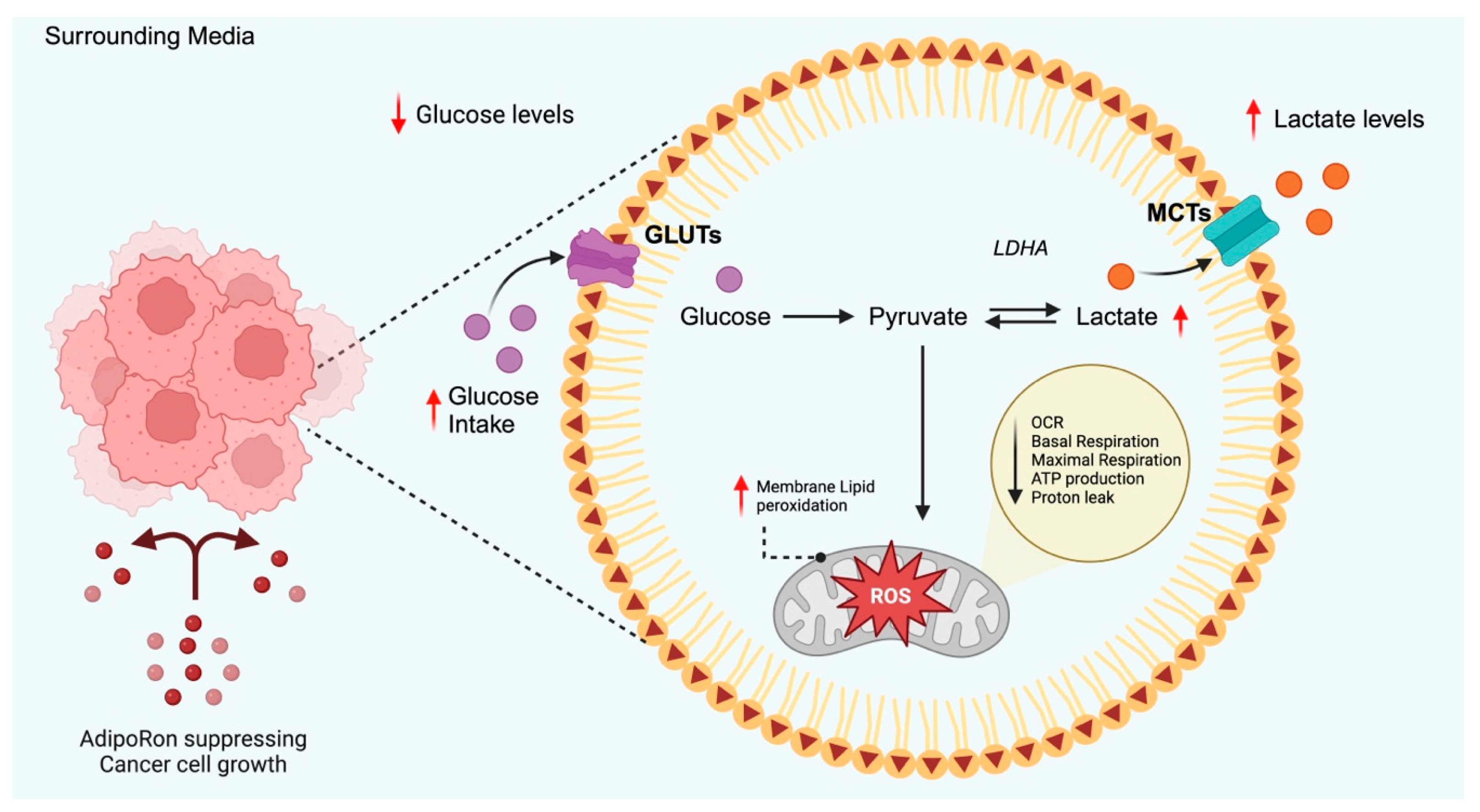
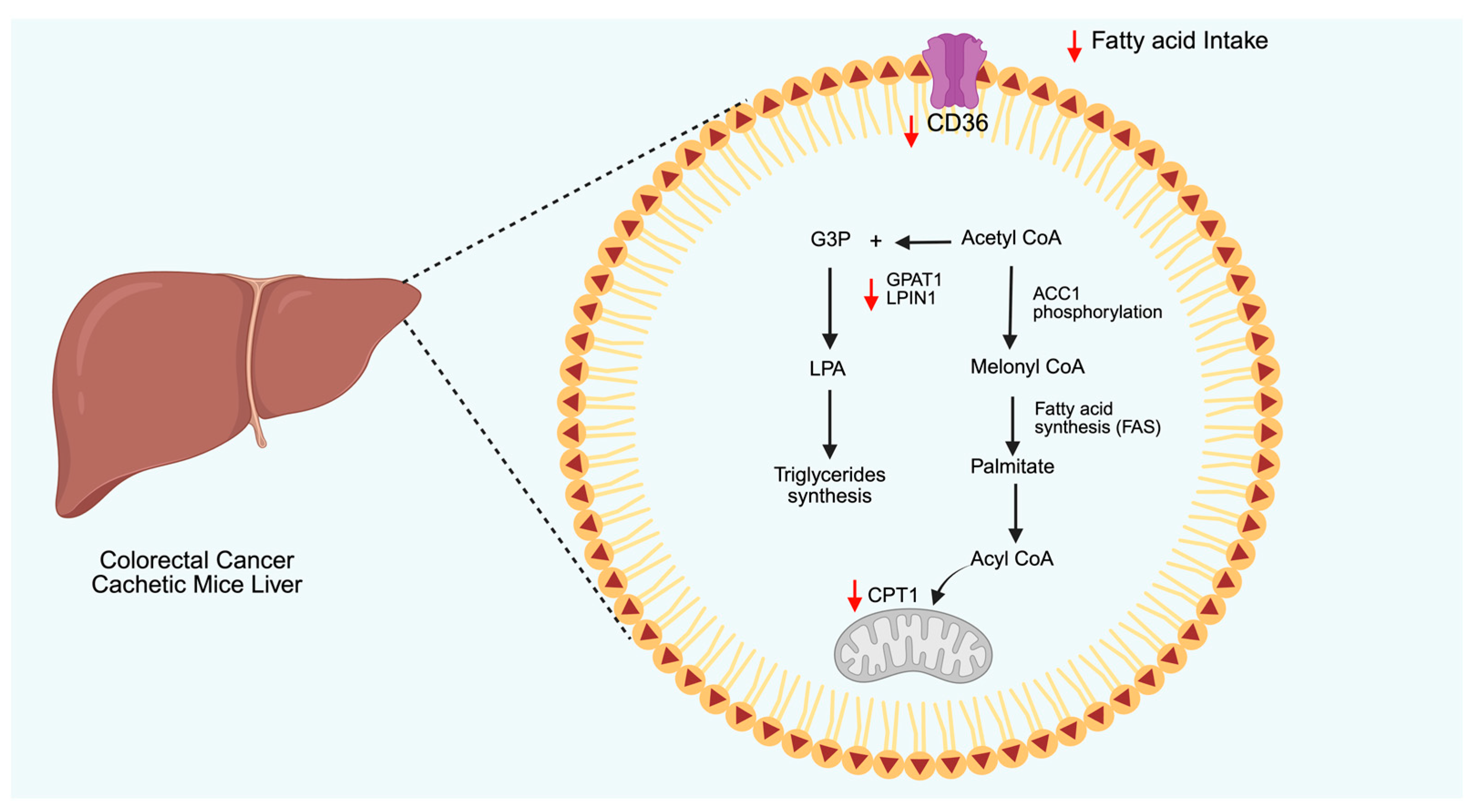
| Type of Cancer | Supporting Data | Conflicting Data |
|---|---|---|
| Breast | Lower adiponectin is associated with a higher risk of breast cancer in postmenopausal women [60,61] | Circulating adiponectin is not related to breast cancer risk in premenopausal women [61] |
| Lung | Adiponectin levels are reduced in lung adenocarcinoma, as well as in advanced patients [62,63] | Bloodstream adiponectin in lung cancer does not differ from that in healthy subjects [62,64] |
| Colorectal | Colorectal cancer cases exhibit diminished adiponectin than those without malignancy [65,66] | Serum adiponectin is not associated with the risk of colorectal cancer [67] |
| Thyroid | Adiponectin is inversely correlated with the risk of thyroid cancer, expressly in women [68] | Thyroid carcinomas and controls display comparable adiponectin levels in blood [69] |
| Endometrial | Lower adiponectin levels increase the risk of endometrial cancer and correlate with its grade [70,71] | Plasma concentration of adiponectin is not predictive of endometrial cancer risk [72] |
| Prostate | Reduced adiponectin expression is linked with a higher degree and stage of prostate cancer [73] | Adiponectin unveils a weak correlation with respect to prostate cancer aggressiveness [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kafeel, S.; Palmiero, G.; Salzillo, A.; Ragone, A.; Naviglio, S.; Sapio, L. Unravelling the Adiponectin Hallmark and Exploring the Therapeutic Potential of Its Receptor Agonists in Cancer Metabolic Reprogramming. Biomolecules 2025, 15, 820. https://doi.org/10.3390/biom15060820
Kafeel S, Palmiero G, Salzillo A, Ragone A, Naviglio S, Sapio L. Unravelling the Adiponectin Hallmark and Exploring the Therapeutic Potential of Its Receptor Agonists in Cancer Metabolic Reprogramming. Biomolecules. 2025; 15(6):820. https://doi.org/10.3390/biom15060820
Chicago/Turabian StyleKafeel, Sanober, Giuseppina Palmiero, Alessia Salzillo, Angela Ragone, Silvio Naviglio, and Luigi Sapio. 2025. "Unravelling the Adiponectin Hallmark and Exploring the Therapeutic Potential of Its Receptor Agonists in Cancer Metabolic Reprogramming" Biomolecules 15, no. 6: 820. https://doi.org/10.3390/biom15060820
APA StyleKafeel, S., Palmiero, G., Salzillo, A., Ragone, A., Naviglio, S., & Sapio, L. (2025). Unravelling the Adiponectin Hallmark and Exploring the Therapeutic Potential of Its Receptor Agonists in Cancer Metabolic Reprogramming. Biomolecules, 15(6), 820. https://doi.org/10.3390/biom15060820