
Melatonin-Mediated Circadian Rhythm Signaling Exhibits Bidirectional Regulatory Effects on the State of Hair Follicle Stem Cells

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Drug Treatment
2.2. Immunofluorescence
2.3. Real-Time qPCR
2.4. Western Blotting
2.5. Droplet Digital PCR
2.6. Cleavage Under Targets and Tagmentation (CUT&Tag)
2.7. Cleavage Under Targets and Release Using Nuclease (CUT&RUN)
2.8. DNA Pull-Down
2.9. RNA Interference
2.10. Dual-Luciferase Reporter
2.11. Statistical Analysis
3. Results
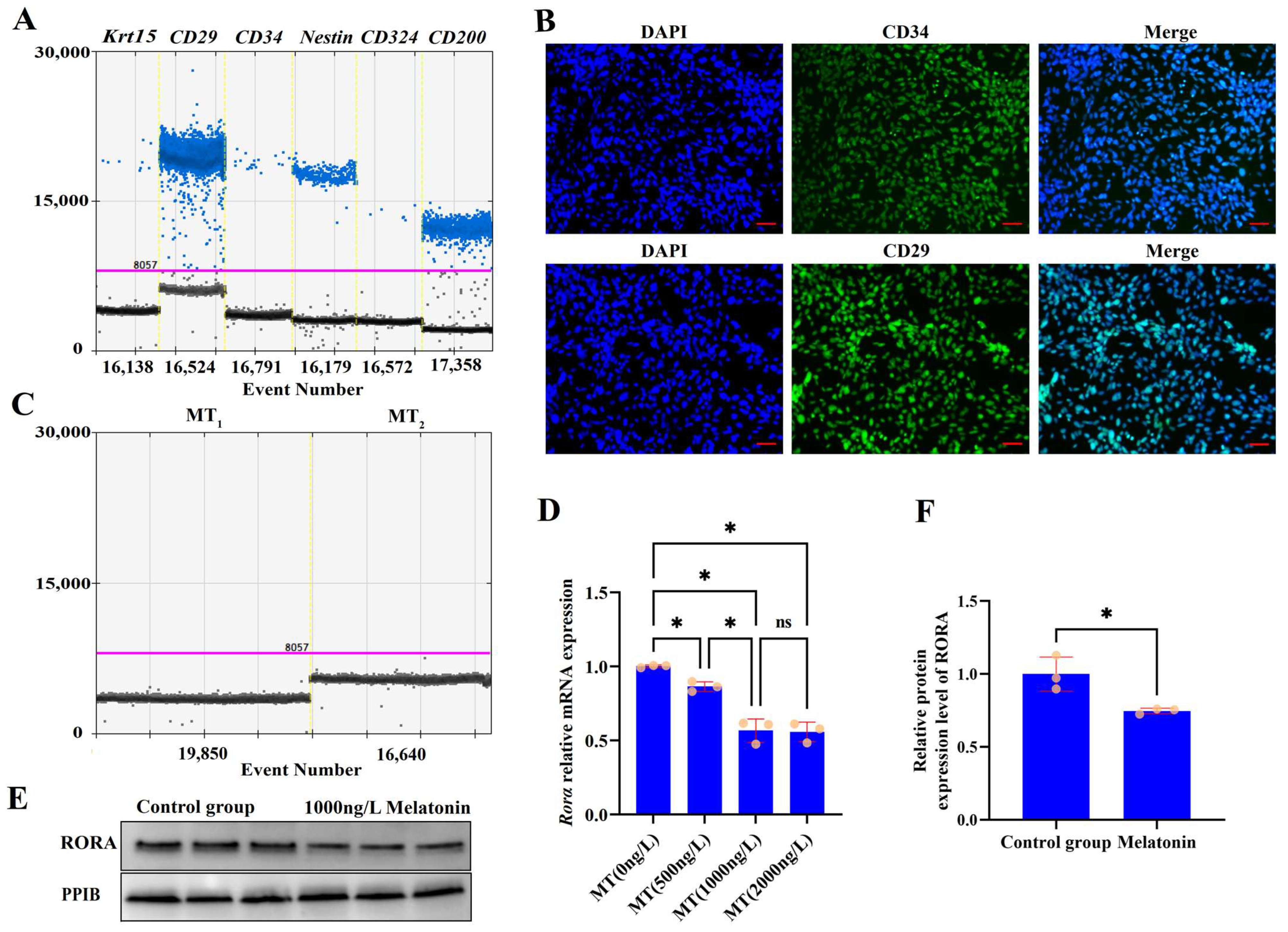
3.1. Characterization and Analysis of Melatonin Receptor Expression in HFSCs
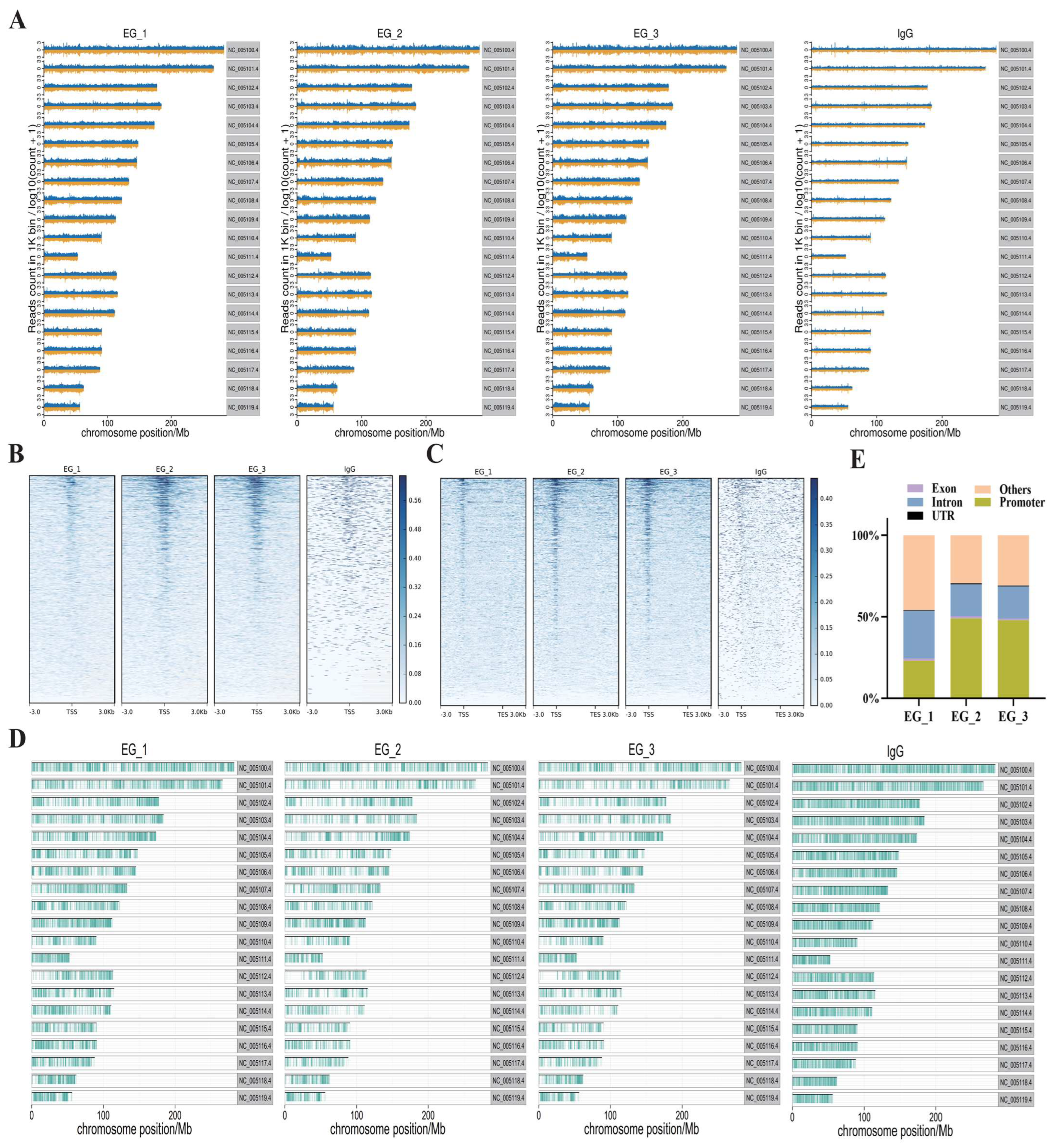
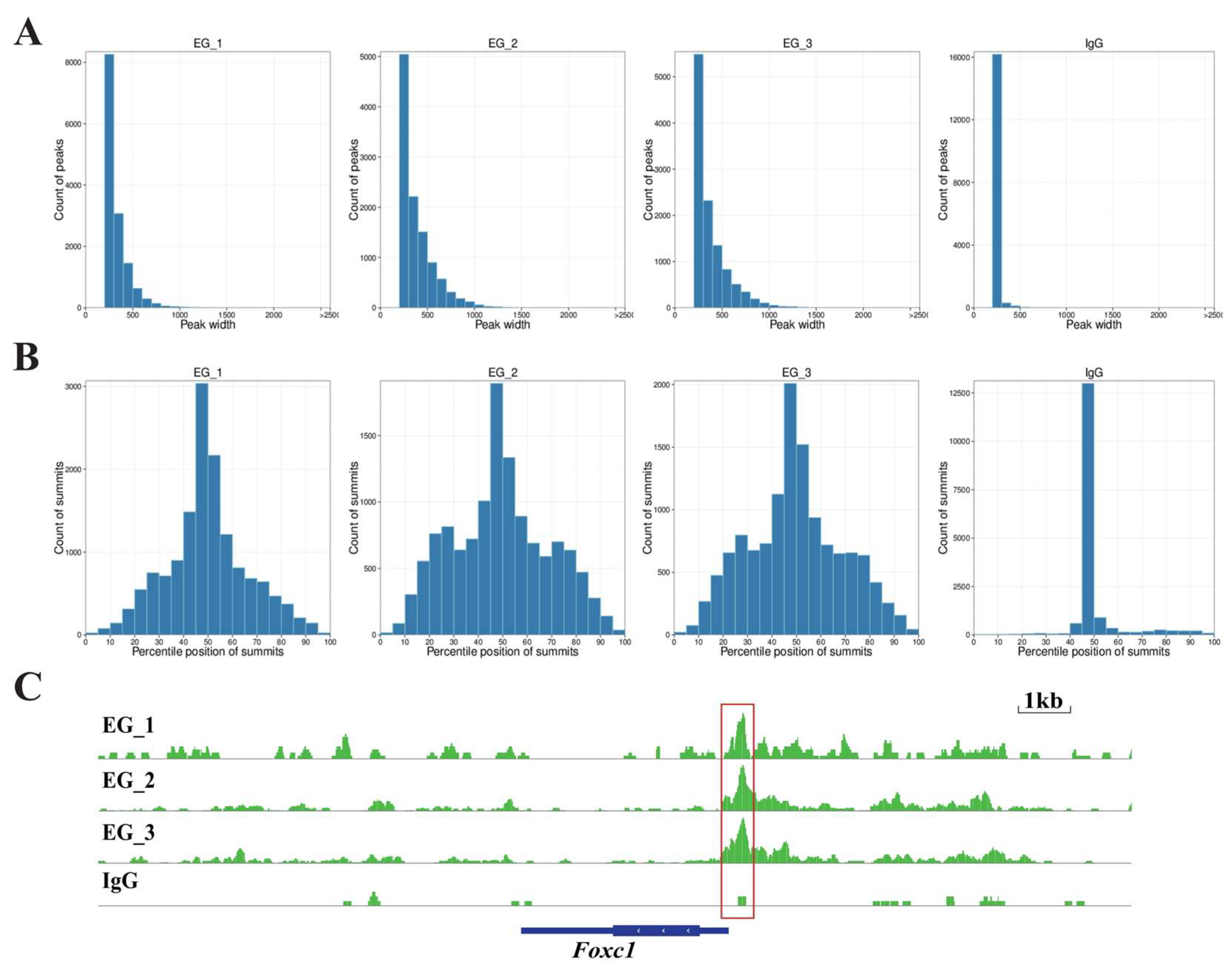
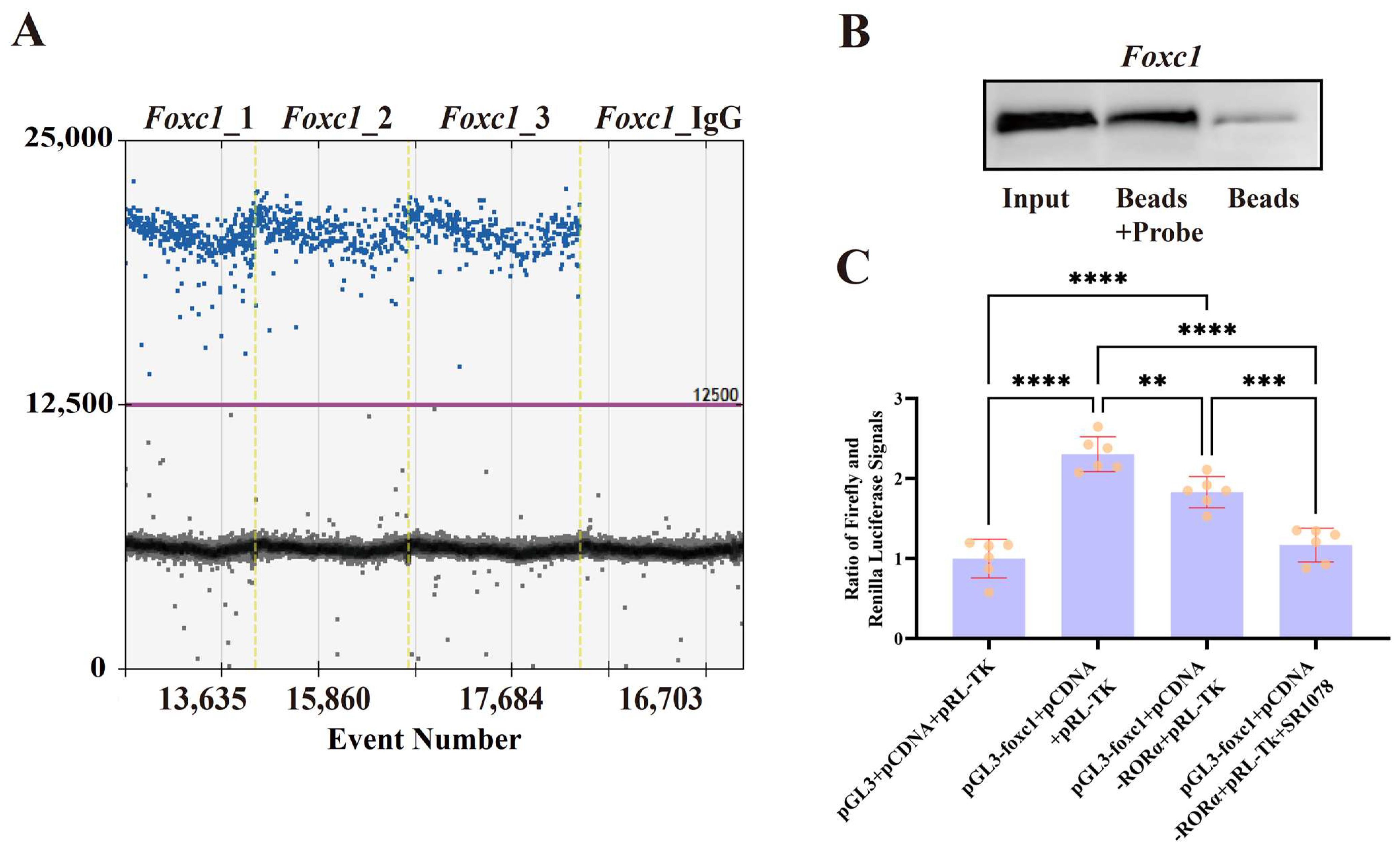
3.2. CUT&Tag
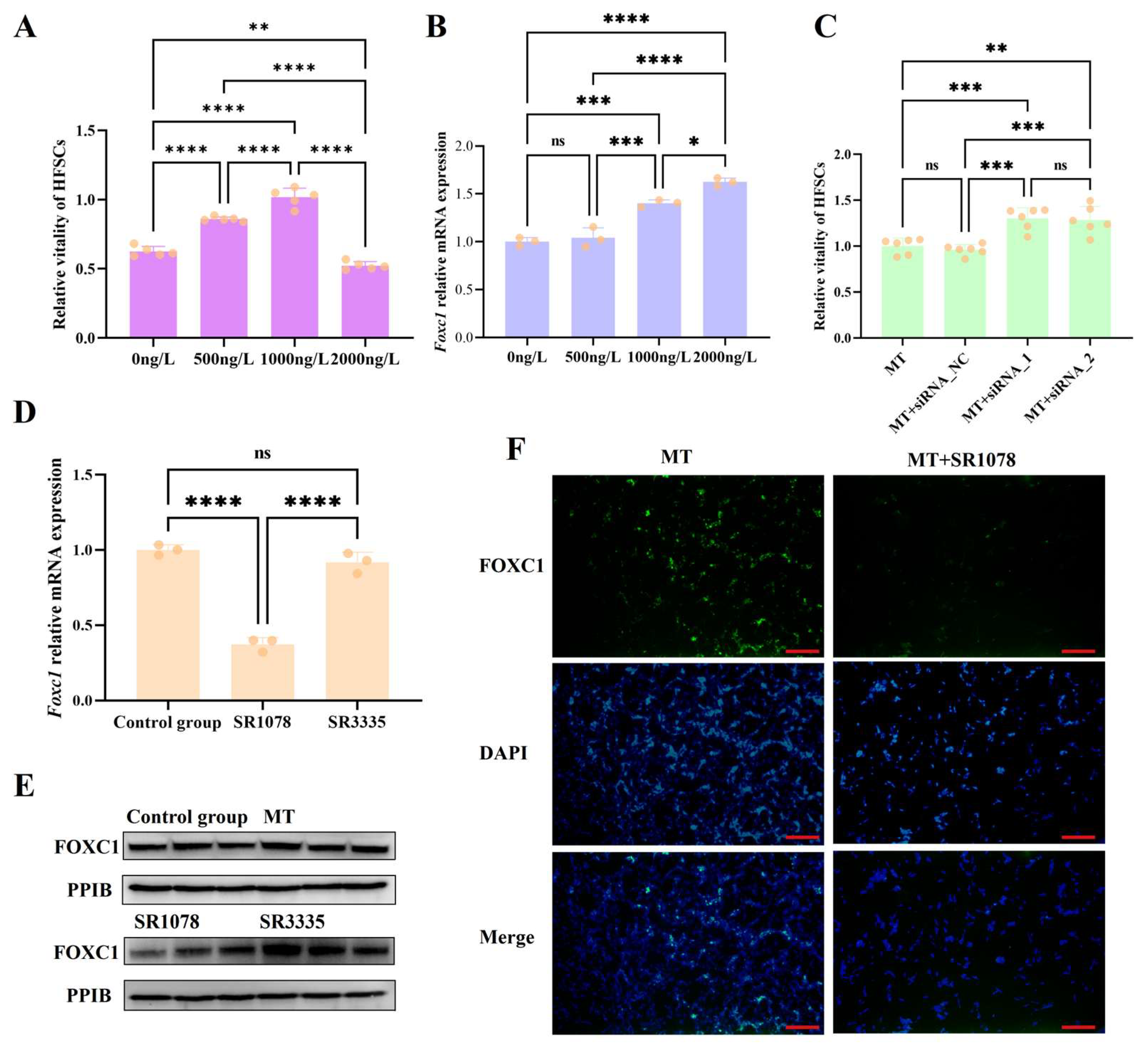
3.3. Melatonin May Regulate Foxc1 Expression Through RORA to Prevent Excessive Activation of HFSCs
3.4. Verification of Interactions Between RORA and Foxc1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Welle, M.M. Basic principles of hair follicle structure, morphogenesis, and regeneration. Vet. Pathol. 2023, 60, 732–747. [Google Scholar] [CrossRef]
- Schneider, M.R.; Schmidt-Ullrich, R.; Paus, R. The Hair Follicle as a Dynamic Miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef] [PubMed]
- Harland, D.P. Introduction to Hair Development. In Hair Fibre: Proteins, Structure and Development; Plowman, J.E., Harland, D.P., DebChoudhury, S., Eds.; Advances in Experimental Medicine and Biology; Springer Nature: London, UK, 2018; Volume 1054, pp. 89–96. [Google Scholar]
- Paus, R.; Foitzik, K. In search of the “hair cycle clock”: A guided tour. Differentiation 2004, 72, 489–511. [Google Scholar] [CrossRef]
- Taylor, G.; Lehrer, M.S.; Jensen, P.J.; Sun, T.T.; Lavker, R.M. Involvement of follicular stem cells in forming not only the follicle but also the epidermis. Cell 2000, 102, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, R.M. The pluripotency of hair follicle stem cells. Cell Cycle 2006, 5, 232–233. [Google Scholar] [CrossRef]
- Joachimiak, R.; Bajek, A.; Drewa, T. Hair follicle as a novel source of stem cells. Postep. Hig. I Med. Dosw. 2012, 66, 181–186. [Google Scholar] [CrossRef]
- Ma, D.R.; Yang, E.N.; Lee, S.T. A review: The location, molecular characterisation and multipotency of hair follicle epidermal stem cells. Ann. Acad. Med. Singap. 2004, 33, 784–788. [Google Scholar] [CrossRef]
- Jaks, V.; Barker, N.; Kasper, M.; Van Es, J.H.; Snippert, H.J.; Clevers, H.; Toftgard, R. Lgr5 marks cycling, yet long-lived, hair follicle stem cells. Nat. Genet. 2008, 40, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, D.; Wang, J.; Wang, L.; Qiu, W.; Kume, T.; Dowell, R.; Yi, R. Escape of hair follicle stem cells causes stem cell exhaustion during aging. Nat. Aging 2021, 1, 889–903. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Siegenthaler, J.A.; Dowell, R.D.; Yi, R. Foxc1 reinforces quiescence in self-renewing hair follicle stem cells. Science 2016, 351, 613–617. [Google Scholar] [CrossRef]
- Chang, Y.; Bao, P.; Chu, M.; Wu, X.; Liang, C.; Yan, P. Research Progress on the Regulation of LncRNA in the Development of Mammalian Hair Follicle. Biotechnol. Bull. 2019, 35, 205–212. [Google Scholar]
- Galbraith, H. Nutritional and hormonal regulation of hair follicle growth and development. Proc. Nutr. Soc. 1998, 57, 195–205. [Google Scholar] [CrossRef]
- Zhou, G.; Wang, X.; Chen, Y.; Kang, D. Potential Involvement of miR-144 in the Regulation of Hair Follicle Development and Cycle Through Interaction with Lhx2. Genes 2024, 15, 1454. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, H.; Jing, J.; Yu, L.; Wu, X.; Lu, Z. Regulation of hair follicle development by exosomes derived from dermal papilla cells. Biochem. Biophys. Res. Commun. 2018, 500, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Randall, V.A. Hormonal regulation of hair follicles exhibits a biological paradox. Semin. Cell Dev. Biol. 2007, 18, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Conrad, F.; Ohnemus, U.; Bodo, E.; Biro, T.; Tychsen, B.; Gerstmayer, B.; Bosio, A.; Schmidt-Rose, T.; Altgilbers, S.; Bettermann, A.; et al. Substantial sex-dependent differences in the response of human scalp hair follicles to estrogen stimulation in vitro advocate gender-tailored management of female versus male pattern balding. J. Investig. Dermatol. Symp. Proc. 2005, 10, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Gao, F.; Guo, J.; Wu, D.; Hao, B.; Li, Y.; Zhao, C. A Microarray-Based Analysis Reveals that a Short Photoperiod Promotes Hair Growth in the Arbas Cashmere Goat. PLoS ONE 2016, 11, e0147124. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Song, S.; Dong, K.; Chen, X.; Liu, X.; Rouzi, M.; Zhao, Q.; He, X.; Pu, Y.; Guan, W.; et al. Skin transcriptome reveals the intrinsic molecular mechanisms underlying hair follicle cycling in Cashmere goats under natural and shortened photoperiod conditions. Sci. Rep. 2017, 7, 13502. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z.; Sun, H.Z.; Zhang, C.H.; Jin, L.; Sang, D.; Li, S.L. Effects of photoperiod on circadian clock genes in skin contribute to the regulation of hair follicle cycling of Inner Mongolia white cashmere goats. Anim. Sci. J. 2020, 91, e13320. [Google Scholar] [CrossRef] [PubMed]
- Emet, M.; Ozcan, H.; Ozel, L.; Yayla, M.; Halici, Z.; Hacimuftuoglu, A. A Review of Melatonin, Its Receptors and Drugs. Eurasian J. Med. 2016, 48, 135–141. [Google Scholar] [CrossRef]
- Jaworek, J.; Leja-Szpak, A.; Nawrot-Porabka, K.; Szklarczyk, J.; Kot, M.; Pierzchalski, P.; Goralska, M.; Ceranowicz, P.; Warzecha, Z.; Dembinski, A.; et al. Effects of Melatonin and Its Analogues on Pancreatic Inflammation, Enzyme Secretion, and Tumorigenesis. Int. J. Mol. Sci. 2017, 18, 1014. [Google Scholar] [CrossRef] [PubMed]
- Kokkola, T.; Laitinen, J.T. Melatonin receptor genes. Ann. Med. 1998, 30, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.F.; Pang, C.S.; Poon, A.M.; Lee, P.P.; Liu, Z.M.; Shiu, S.Y. Melatonin: A chemical photoperiodic signal with clinical significance in humans. Chin. Med. J. 1998, 111, 197–203. [Google Scholar]
- Reiter, R.J.; Tan, D.-X.; Fuentes-Broto, L. Melatonin: A multitasking molecule. In Neuroendocrinology: The Normal Neuroendocrine System; Martini, L., Chrousos, G.P., Labrie, F., Pacak, K., Pfaff, D.W., Eds.; Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2010; Volume 181, pp. 127–151. [Google Scholar]
- Ackermann, K.; Stehle, J.H. Melatonin synthesis in the human pineal gland: Advantages, implications, and difficulties. Chronobiol. Int. 2006, 23, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Chattoraj, A.; Liu, T.; Zhang, L.S.; Huang, Z.; Borjigin, J. Melatonin formation in mammals: In vivo perspectives. Rev. Endocr. Metab. Disord. 2009, 10, 237–243. [Google Scholar] [CrossRef] [PubMed]
- do Amaral, F.G.; Cipolla-Neto, J. A brief review about melatonin, a pineal hormone. Arch. Endocrinol. Metab. 2018, 62, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Tosini, G. Melatonin circadian rhythm in the retina of mammals. Chronobiol. Int. 2000, 17, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Tosini, G.; Fukuhara, C. Photic and circadian regulation of retinal melatonin in mammals. J. Neuroendocrinol. 2003, 15, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Hu, V.W. Is retinoic acid-related orphan receptor-alpha (RORA) a target for gene-environment interactions contributing to autism? Neurotoxicology 2012, 33, 1434–1435. [Google Scholar] [CrossRef]
- Jetten, A.M. Retinoid-related orphan receptors (RORs): Critical roles in development, immunity, circadian rhythm, and cellular metabolism. Nucl. Recept. Signal. 2009, 7, nrs-07003. [Google Scholar] [CrossRef]
- Emery, P.; Reppert, S.M. A rhythmic ror. Neuron 2004, 43, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Boukhtouche, F.; Mariani, J.; Tedgui, A. The “CholesteROR” protective pathway in the vascular system. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 637–643. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.-K.; Takeda, Y.; Janjetovic, Z.; Brozyna, A.A.; Skobowiat, C.; Wang, J.; Postlethwaite, A.; Li, W.; Tuckey, R.C.; et al. RORα and RORγ are expressed in human skin and serve as receptors for endogenously produced noncalcemic 20-hydroxy- and 20,23-dihydroxyvitamin D. Faseb J. 2014, 28, 2775–2789. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Jetten, A.M. RORα is not a receptor for melatonin. Bioessays 2016, 38, 1193–1194. [Google Scholar] [CrossRef] [PubMed]
- Babadjouni, A.; Reddy, M.; Zhang, R.; Raffi, J.; Phong, C.; Mesinkovska, N. Melatonin and the Human Hair Follicle. J. Drugs Dermatol. JDD 2023, 22, 260–264. [Google Scholar] [CrossRef]
- Fischer, T.W.; Burmeister, G.; Schmidt, H.W.; Elsner, P. Melatonin increases anagen hair rate in women with androgenetic alopecia or diffuse alopecia: Results of a pilot randomized controlled trial. Br. J. Dermatol. 2004, 150, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Hardeland, R.; Zmijewski, M.A.; Slominski, R.M.; Reiter, R.J.; Paus, R. Melatonin: A Cutaneous Perspective on its Production, Metabolism, and Functions. J. Investig. Dermatol. 2018, 138, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.-L.; Li, Y.-K.; Gao, C.-X.; Li, W.-W.; Li, L.; Wang, H.; Shen, W.; Ge, W. Melatonin promotes hair regeneration by modulating the Wnt/β-catenin signalling pathway. Cell Prolif. 2024, 57, e13656. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Rochat, A.; Kedzia, C.; Kobayashi, K.; Barrandon, Y. Morphogenesis and renewal of hair follicles from adult multipotent stem cells. Cell 2001, 104, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Rochat, A.; Kobayashi, K.; Barrandon, Y. Location of stem cells of human hair follicles by clonal analysis. Cell 1994, 76, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Shwartz, Y.; Gonzalez-Celeiro, M.; Chen, C.-L.; Pasolli, H.A.; Sheu, S.-H.; Fan, S.M.-Y.; Shamsi, F.; Assaad, S.; Lin, E.T.-Y.; Zhang, B.; et al. Cell Types Promoting Goosebumps Form a Niche to Regulate Hair Follicle Stem Cells. Cell 2020, 182, 578–593.e19. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Aoi, N.; Sato, T.; Yamauchi, Y.; Suga, H.; Eto, H.; Kato, H.; Araki, J.; Yoshimura, K. Differential expression of stem-cell-associated markers in human hair follicle epithelial cells. Lab. Investig. 2009, 89, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Lyle, S.; Yang, Z.X.; Cotsarelis, G. Keratin 15 promoter targets putative epithelial stem cells in the hair follicle bulge. J. Investig. Dermatol. 2003, 121, 963–968. [Google Scholar] [CrossRef]
- Amoh, Y.; Li, L.; Katsuoka, K.; Penman, S.; Hoffman, R.M. Multipotent nestin-positive, keratin-negative hair-follicle bulge stem cells can form neurons. Proc. Natl. Acad. Sci. USA 2005, 102, 5530–5534. [Google Scholar] [CrossRef]
- Ohyama, M.; Terunuma, A.; Tock, C.L.; Radonovich, M.F.; Pise-Masison, C.A.; Hopping, S.B.; Brady, J.N.; Udey, M.C.; Vogel, J.C. Characterization and isolation of stem cell-enriched human hair follicle bulge cells. J. Clin. Investig. 2006, 116, 249–260. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.-K.; Slominski, R.M.; Song, Y.; Qayyum, S.; Placha, W.; Janjetovic, Z.; Kleszczynski, K.; Atigadda, V.; Song, Y.; et al. Melatonin and Its Metabolites Can Serve as Agonists on the Aryl Hydrocarbon Receptor and Peroxisome Proliferator-Activated Receptor Gamma. Int. J. Mol. Sci. 2023, 24, 15496. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Kim, T.-K.; Janjetovic, Z.; Slominski, R.M.; Zmijewski, J.W. Melatonin, mitochondria, and the skin. Cell. Mol. Life Sci. 2017, 74, 3913–3925. [Google Scholar] [CrossRef]
- Samra, T.; Cheret, J.; Gherardini, J.; Verling, S.; Kassir, R.; Paus, R. Melatonin Protects K15+ Human Hair Follicle Stem Cells and Hair Matrix Keratinocytes against Paclitaxel-Induced Damage Ex Vivo. J. Investig. Dermatol. 2024, 144, 697–701.e6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, N.; Zhang, T.; Wang, M.; Ge, W.; Wang, X. Roles of Melatonin in Goat Hair Follicle Stem Cell Proliferation and Pluripotency Through Regulating the Wnt Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 686805. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhao, X.; Li, S.; Xu, Y.; Bai, S.; Zhang, W. Melatonin-Mediated Circadian Rhythm Signaling Exhibits Bidirectional Regulatory Effects on the State of Hair Follicle Stem Cells. Biomolecules 2025, 15, 226. https://doi.org/10.3390/biom15020226
Zhang Y, Zhao X, Li S, Xu Y, Bai S, Zhang W. Melatonin-Mediated Circadian Rhythm Signaling Exhibits Bidirectional Regulatory Effects on the State of Hair Follicle Stem Cells. Biomolecules. 2025; 15(2):226. https://doi.org/10.3390/biom15020226
Chicago/Turabian StyleZhang, Yu, Xuefei Zhao, Shuqi Li, Yanchun Xu, Suying Bai, and Wei Zhang. 2025. "Melatonin-Mediated Circadian Rhythm Signaling Exhibits Bidirectional Regulatory Effects on the State of Hair Follicle Stem Cells" Biomolecules 15, no. 2: 226. https://doi.org/10.3390/biom15020226
APA StyleZhang, Y., Zhao, X., Li, S., Xu, Y., Bai, S., & Zhang, W. (2025). Melatonin-Mediated Circadian Rhythm Signaling Exhibits Bidirectional Regulatory Effects on the State of Hair Follicle Stem Cells. Biomolecules, 15(2), 226. https://doi.org/10.3390/biom15020226