
Activity Evaluation and Mode of Action of ICA Against Toxoplasma gondii In Vitro

 ,
,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
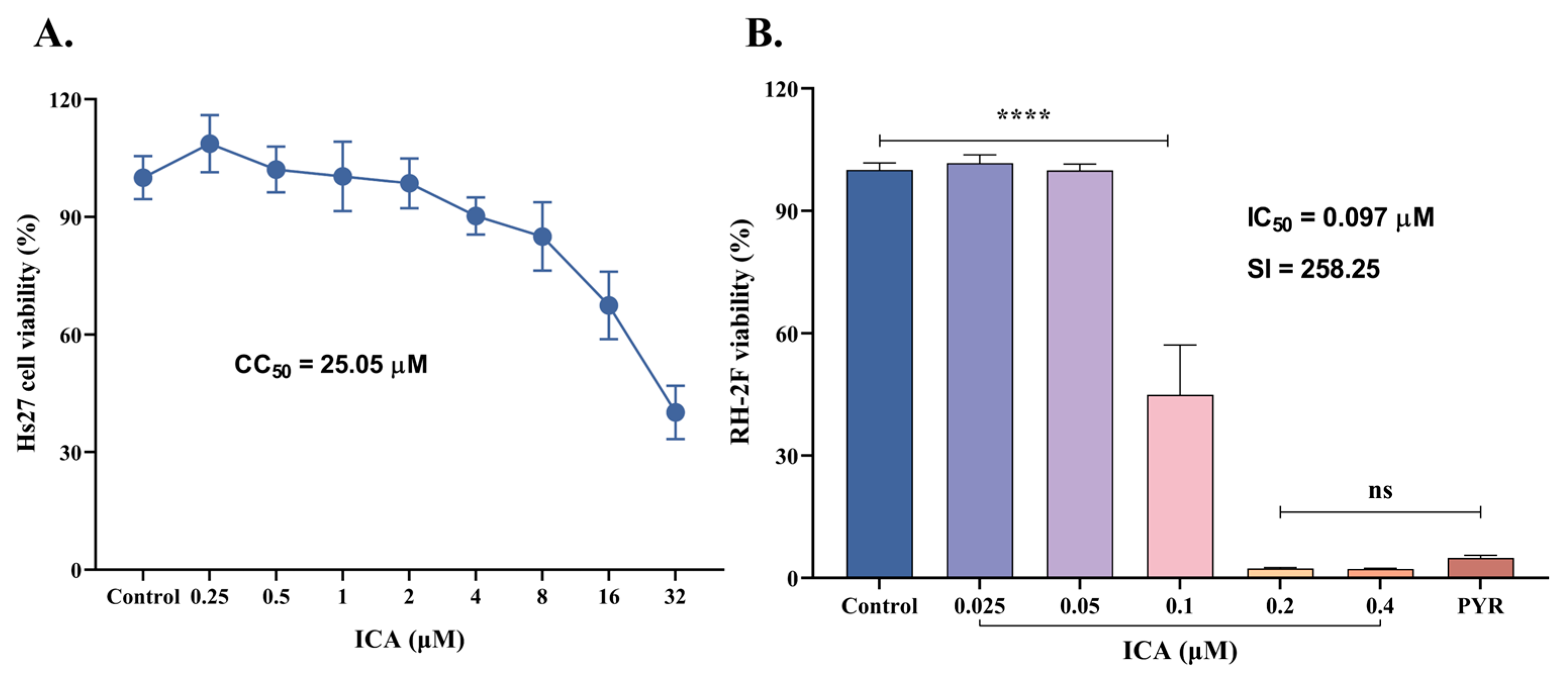
2.1. Cytotoxicity Assay
2.2. Inhibition Assay
2.3. Antiproliferation Assay
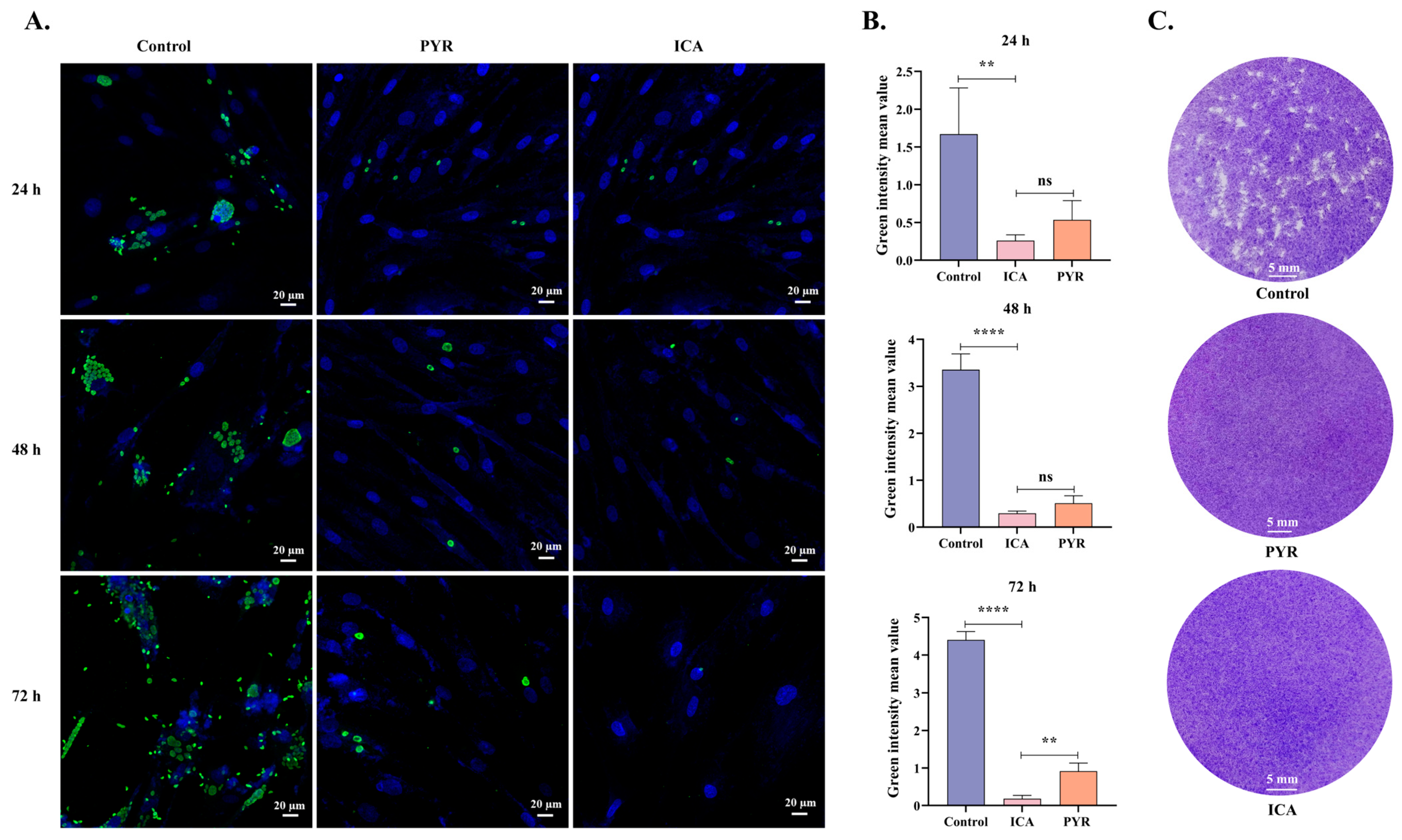
2.4. Plaque Assay
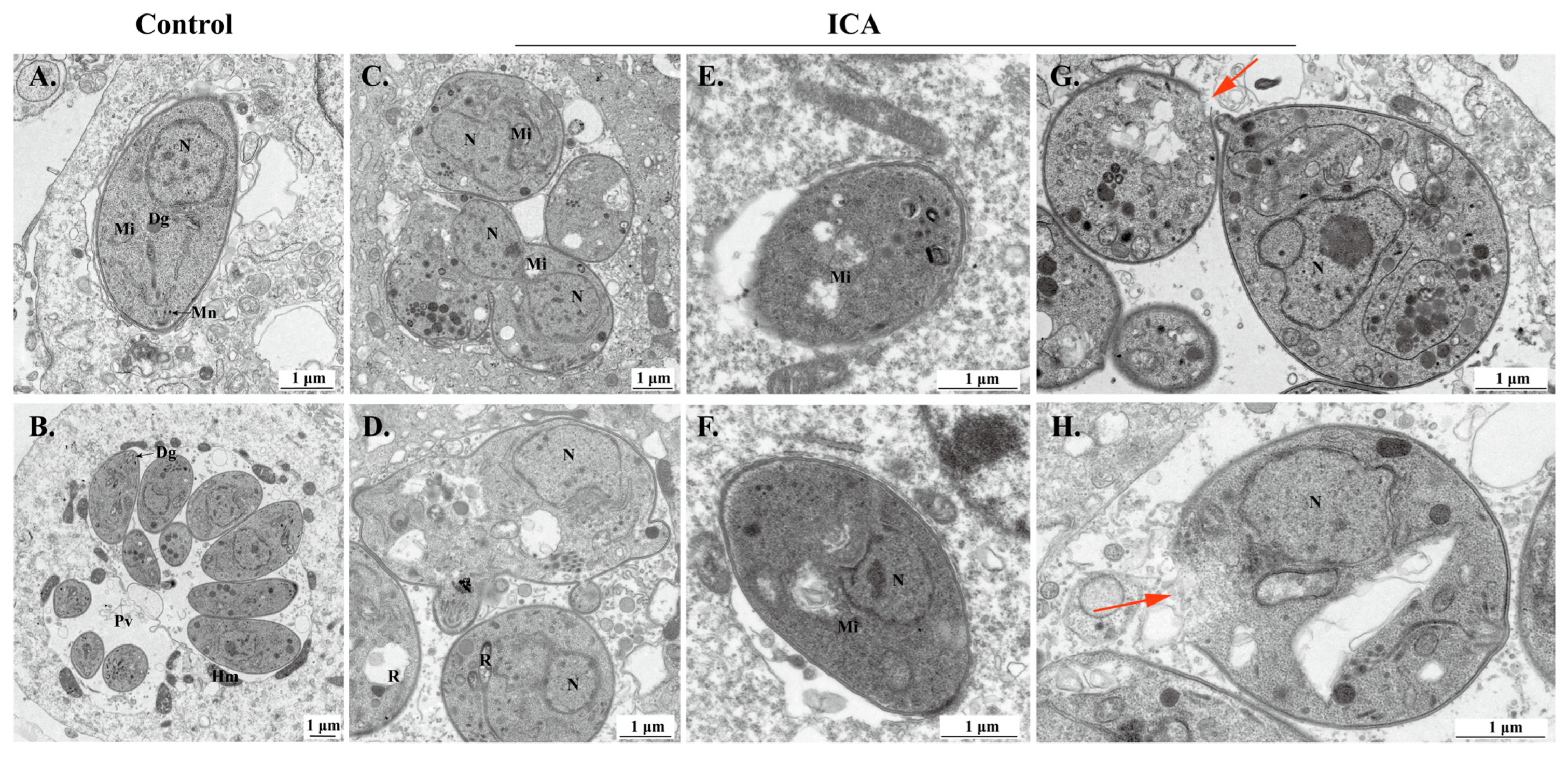
2.5. Transmission Electron Microscopy Assay
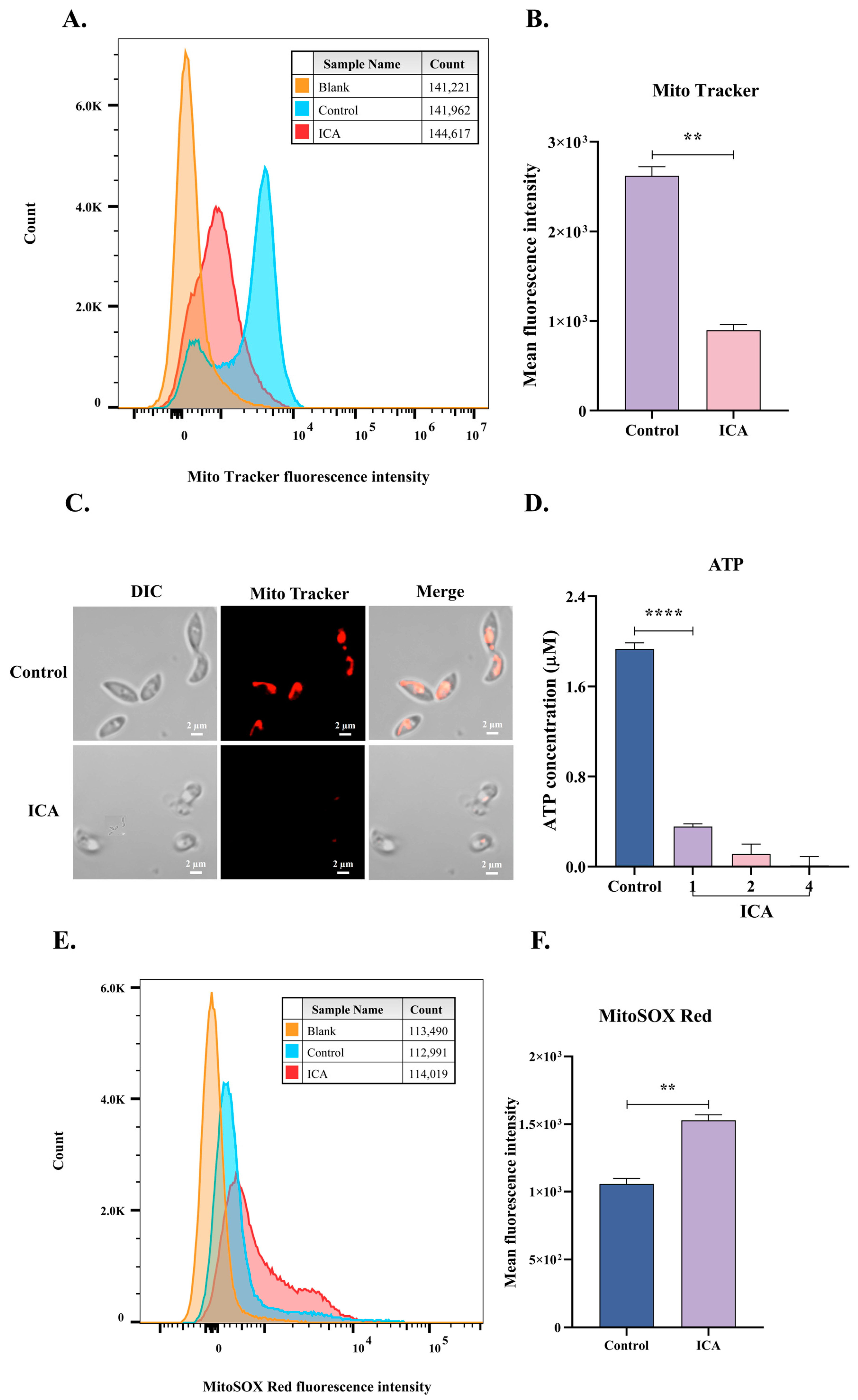
2.6. Mitochondrial Membrane Potential Assay of Extracellular T. gondii
2.7. Mitochondrial Superoxide Assay
2.8. ATP Content Assay
2.9. Mitochondrial Membrane Potential Assay of Hs27 Cells
3. Results
3.1. Safety and Efficacy of ICA Against T. gondii Infection
3.2. ICA Suppresses T. gondii Proliferation
3.3. ICA Alters T. gondii Morphology
3.4. ICA Causes Mitochondrial Dysfunction in T. gondii
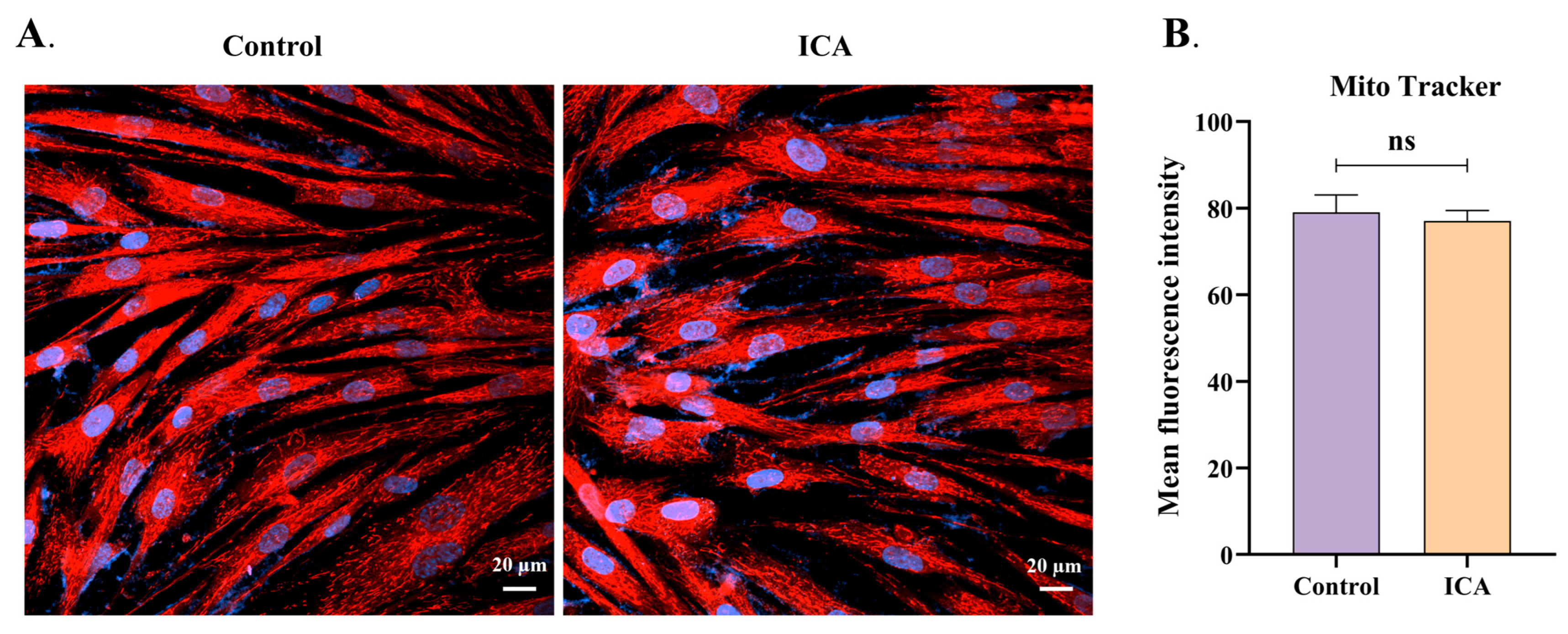
3.5. ICA Does Not Alter the Mitochondrial Membrane Potential of Host Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lourido, S. Toxoplasma gondii. Trends Parasitol. 2019, 35, 944–945. [Google Scholar] [CrossRef] [PubMed]
- Kochanowsky, J.A.; Koshy, A.A. Toxoplasma gondii. Curr. Biol. 2018, 28, R770–R771. [Google Scholar] [CrossRef]
- Dlugonska, H. Are poikilothermic animals real hosts for Toxoplasma gondii? Ann. Parasitol. 2017, 63, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, T.; Sarvi, S.; Daryani, A. Toxoplasma gondii in mollusks and cold-blooded animals: A systematic review. Parasitology 2021, 148, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, Z.; Wang, S.; Tao, L.F.; Xu, L.X.; Yan, R.F.; Song, X.K.; Li, X.R. Detection of Toxoplasma gondii in shellfish and fish in parts of China. Vet. Parasitol. 2014, 200, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Attias, M.; Teixeira, D.E.; Benchimol, M.; Vommaro, R.C.; Crepaldi, P.H.; De Souza, W. The life-cycle of Toxoplasma gondii reviewed using animations. Parasit. Vectors 2020, 13, 588. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumètre, A.; de Wit, L.A.; VanWormer, E.; Villena, I. Environmental transmission of Toxoplasma gondii: Oocysts in water, soil and food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Shapiro, K.; VanWormer, E. Dynamics and epidemiology of Toxoplasma gondii oocyst shedding in domestic and wild felids. Transbound. Emerg. Dis. 2022, 69, 2412–2423. [Google Scholar] [CrossRef] [PubMed]
- Flegr, J.; Prandota, J.; Sovičková, M.; Israili, Z.H. Toxoplasmosis—A global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries. PLoS ONE 2014, 9, e90203. [Google Scholar] [CrossRef] [PubMed]
- Demar, M.; Hommel, D.; Djossou, F.; Peneau, C.; Boukhari, R.; Louvel, D.; Bourbigot, A.-M.; Nasser, V.; Ajzenberg, D.; Darde, M.-L.; et al. Acute toxoplasmoses in immunocompetent patients hospitalized in an intensive care unit in French Guiana. Clin. Microbiol. Infect. 2012, 18, E221–E231. [Google Scholar] [CrossRef]
- Muller, J.; Hemphill, A. Toxoplasma gondii infection: Novel emerging therapeutic targets. Expert Opin. Ther. Targets 2023, 27, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Antczak, M.; Dzitko, K.; Długońska, H. Human toxoplasmosis–Searching for novel chemotherapeutics. Biomed. Pharmacother. 2016, 82, 677–684. [Google Scholar] [CrossRef]
- Alday, P.H.; Doggett, J.S. Drugs in development for toxoplasmosis: Advances, challenges, and current status. Drug Des. Devel Ther. 2017, 11, 273–293. [Google Scholar] [CrossRef] [PubMed]
- Mccabe, R.E.; Oster, S. Current recommendations and future prospects in the treatment of toxoplasmosis. Drugs 1989, 38, 973–987. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of toxoplasmosis: Historical perspective, animal models, and current clinical practice. Clin. Microbiol. Rev. 2018, 31, e00017–e00057. [Google Scholar] [CrossRef]
- Kortagere, S. Screening for small molecule inhibitors of Toxoplasma gondii. Expert Opin. Drug Discov. 2012, 7, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Tassone, G.; Landi, G.; Linciano, P.; Francesconi, V.; Tonelli, M.; Tagliazucchi, L.; Costi, M.P.; Mangani, S.; Pozzi, C. Evidence of pyrimethamine and cycloguanil analogues as dual inhibitors of Trypanosoma brucei pteridine reductase and dihydrofolate reductase. Pharmaceuticals 2021, 14, 636. [Google Scholar] [CrossRef] [PubMed]
- Paquet, T.; Gordon, R.; Waterson, D.; Witty, M.J.; Chibale, K. Antimalarial aminothiazoles and aminopyridines from phenotypic whole-cell screening of a SoftFocus((R)) library. Future Med. Chem. 2012, 4, 2265–2277. [Google Scholar] [CrossRef]
- Ghaemmaghami, S.; Russo, M.; Renslo, A.R. Successes and challenges in phenotype-based lead discovery for prion diseases. J. Med. Chem. 2014, 57, 6919–6929. [Google Scholar] [CrossRef]
- Relave, E.T.; Hedna, R.; Di Maio, A.; Devred, F.; Kovacic, H.; Robin, M.; Breuzard, G. Therapeutic contribution of Tau-binding thiazoloflavonoid hybrid derivatives against glioblastoma using pharmacological approach in 3D spheroids. Int. J. Mol. Sci. 2024, 25, 11785. [Google Scholar] [CrossRef] [PubMed]
- Girardini, M.; Ferlenghi, F.; Annunziato, G.; Degiacomi, G.; Papotti, B.; Marchi, C.; Sammartino, J.C.; Rasheed, S.S.; Contini, A.; Pasca, M.R.; et al. Expanding the knowledge around antitubercular 5-(2-aminothiazol-4-yl) isoxazole-3-carboxamides: Hit-to-lead optimization and release of a novel antitubercular chemotype via scaffold derivatization. Eur. J. Med. Chem. 2023, 245, 114916. [Google Scholar] [CrossRef] [PubMed]
- Gentles, R.G.; Grant-Young, K.; Hu, S.; Huang, Y.; Poss, M.A.; Andres, C.; Fiedler, T.; Knox, R.; Lodge, N.; Weaver, C.D.; et al. Initial SAR studies on apamin-displacing 2-aminothiazole blockers of calcium-activated small conductance potassium channels. Bioorg. Med. Chem. Lett. 2008, 18, 5316–5319. [Google Scholar] [CrossRef]
- Diness, J.G.; Sørensen, U.S.; Nissen, J.D.; Al-Shahib, B.; Jespersen, T.; Grunnet, M.; Hansen, R.S. Inhibition of small-conductance Ca2+-activated K+ channels terminates and protects against atrial fibrillation. Circ. Arrhythm. Electrophysiol. 2010, 3, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Skarsfeldt, M.A.; Diness, J.G.; Bentzen, B.H. Small conductance calcium activated K+ channel inhibitor decreases stretch induced vulnerability to atrial fibrillation. Int. J. Cardiol. Heart Vasc. 2021, 37, 100898. [Google Scholar] [CrossRef] [PubMed]
- Bhuniya, D.; Mukkavilli, R.; Shivahare, R.; Launay, D.; Dere, R.T.; Deshpande, A.; Verma, A.; Vishwakarma, P.; Moger, M.; Pradhan, A.; et al. Aminothiazoles: Hit to lead development to identify antileishmanial agents. Eur. J. Med. Chem. 2015, 102, 582–593. [Google Scholar] [CrossRef]
- Qiu, Y.; Zhai, B.; Bai, Y.; Lin, H.; Wu, L.; Luo, W.; Shi, M.; Chen, S.; Zhang, J. In vitro and in vivo activity evaluation and mode of action of broxaldine on Toxoplasma gondii. Int. J. Parasitol. Drugs Drug Resist. 2024, 25, 100552. [Google Scholar] [CrossRef]
- Dubey, J.P.; Lindsay, D.S.; Speer, C.A. Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clin. Microbiol. Rev. 1998, 11, 267–299. [Google Scholar] [CrossRef] [PubMed]
- Azami, S.J.; Rahimi, H.M.; Mirjalali, H.; Zali, M.R. Unravelling Toxoplasma treatment: Conventional drugs toward nanomedicine. World J. Microbiol. Biotechnol. 2021, 37, 48. [Google Scholar] [CrossRef]
- Brookes, P.S.; Yoon, Y.; Robotham, J.L.; Anders, M.W.; Sheu, S.-S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am. J. Physiol. Cell Physiol. 2004, 287, C817–C833. [Google Scholar] [CrossRef] [PubMed]
- Sakamuru, S.; Attene-Ramos, M.S.; Xia, M. Mitochondrial membrane potential assay. Methods Mol. Biol. 2022, 2474, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Wescott, H.H.; Zuniga, E.S.; Bajpai, A.; Trujillo, C.; Ehrt, S.; Schnappinger, D.; Roberts, D.M.; Parish, T. Identification of enolase as the target of 2-aminothiazoles in Mycobacterium tuberculosis. Front. Microbiol. 2018, 9, 2542. [Google Scholar] [CrossRef]
- Mouveaux, T.; Oria, G.; Werkmeister, E.; Slomianny, C.; Fox, B.A.; Bzik, D.J.; Tomavo, S. Nuclear glycolytic enzyme enolase of Toxoplasma gondii functions as a transcriptional regulator. PLoS ONE 2014, 9, e105820. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Y.; Wang, W.; Wang, Q.; Xu, J.; Dai, G.; Bai, Y.; Zhang, J. Activity Evaluation and Mode of Action of ICA Against Toxoplasma gondii In Vitro. Biomolecules 2025, 15, 202. https://doi.org/10.3390/biom15020202
Qiu Y, Wang W, Wang Q, Xu J, Dai G, Bai Y, Zhang J. Activity Evaluation and Mode of Action of ICA Against Toxoplasma gondii In Vitro. Biomolecules. 2025; 15(2):202. https://doi.org/10.3390/biom15020202
Chicago/Turabian StyleQiu, Yanhua, Weiwei Wang, Qing Wang, Jing Xu, Guonian Dai, Yubin Bai, and Jiyu Zhang. 2025. "Activity Evaluation and Mode of Action of ICA Against Toxoplasma gondii In Vitro" Biomolecules 15, no. 2: 202. https://doi.org/10.3390/biom15020202
APA StyleQiu, Y., Wang, W., Wang, Q., Xu, J., Dai, G., Bai, Y., & Zhang, J. (2025). Activity Evaluation and Mode of Action of ICA Against Toxoplasma gondii In Vitro. Biomolecules, 15(2), 202. https://doi.org/10.3390/biom15020202