
C/EBPβ Regulates HIF-1α-Driven Invasion of Non-Small-Cell Lung Cancer Cells

Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture, Hypoxic Condition, and Transfection
2.2. Cell Proliferation Assay
2.3. Migration and Invasion Assay
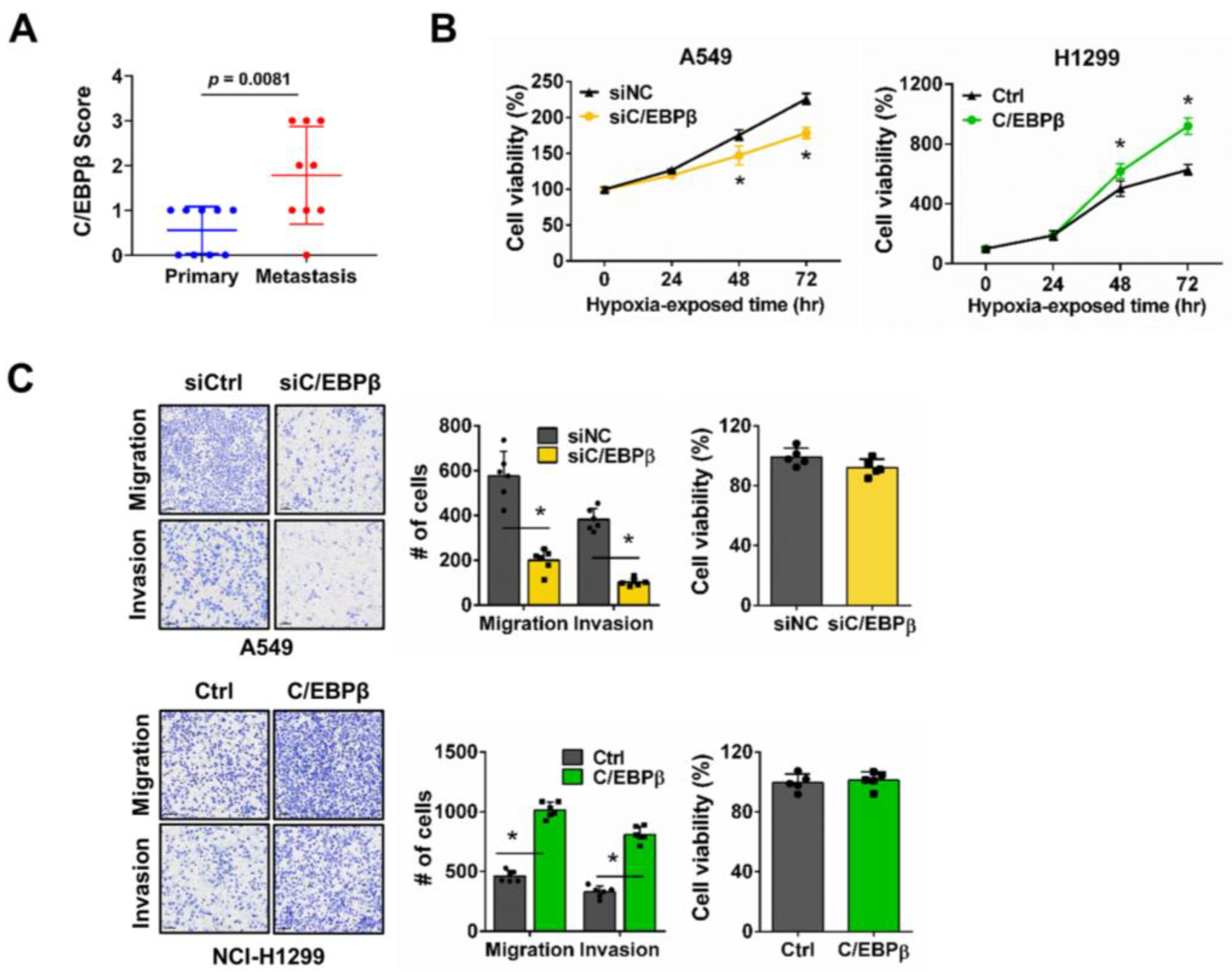
2.4. Tissue Microarray (TMA) Analysis
2.5. Microarray and Bioinformatics Analysis
2.6. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
2.7. Western Blot Analysis
2.8. Chromatin Immunoprecipitation Analysis
2.9. Statistical Analysis
3. Results
3.1. C/EBPβ Increases Proliferation, Migration, and Invasion in NSCLC Cells Under Hypoxic Conditions
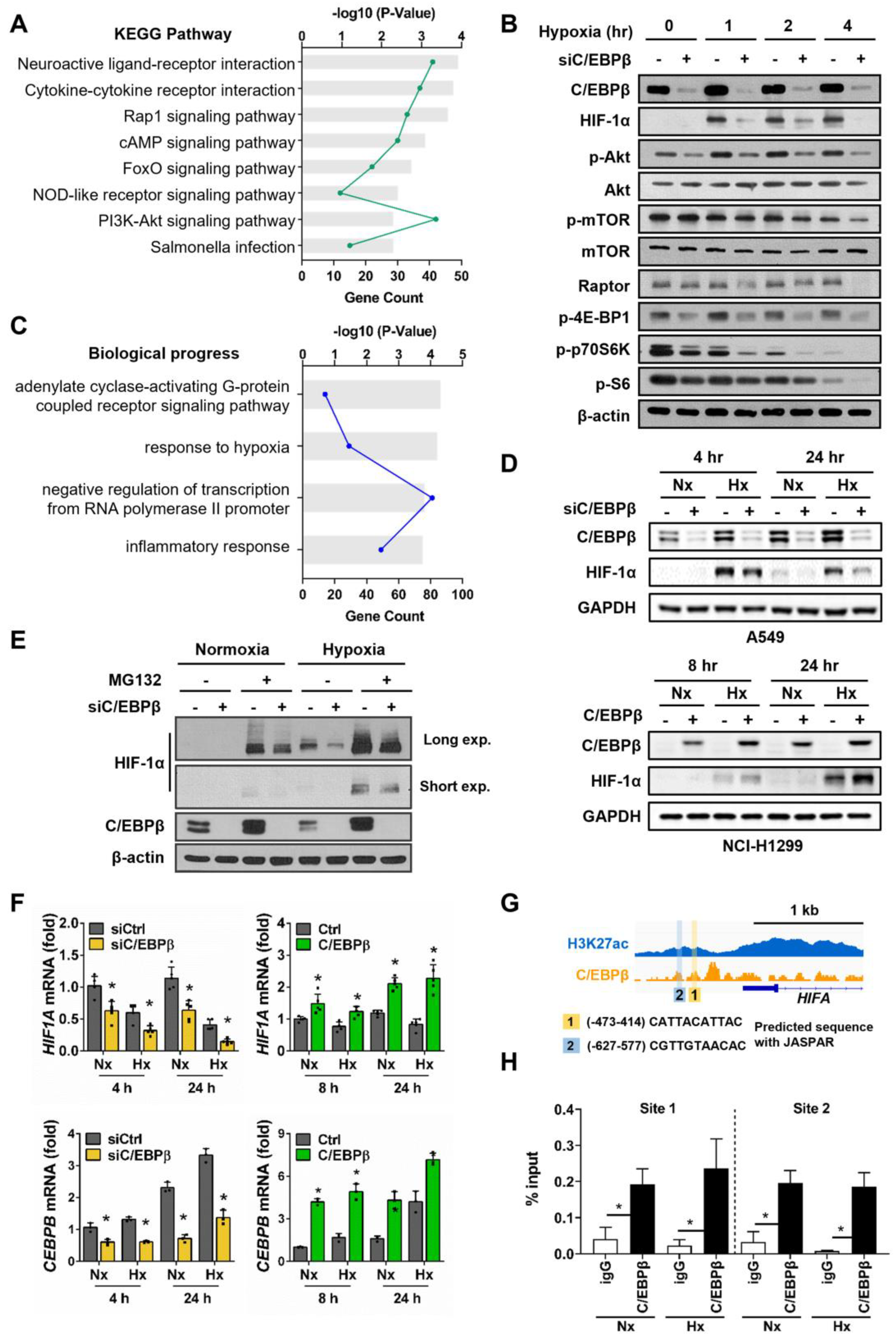
3.2. C/EBPβ Regulates HIF1A Expression by Directly Binding to Its Promoter
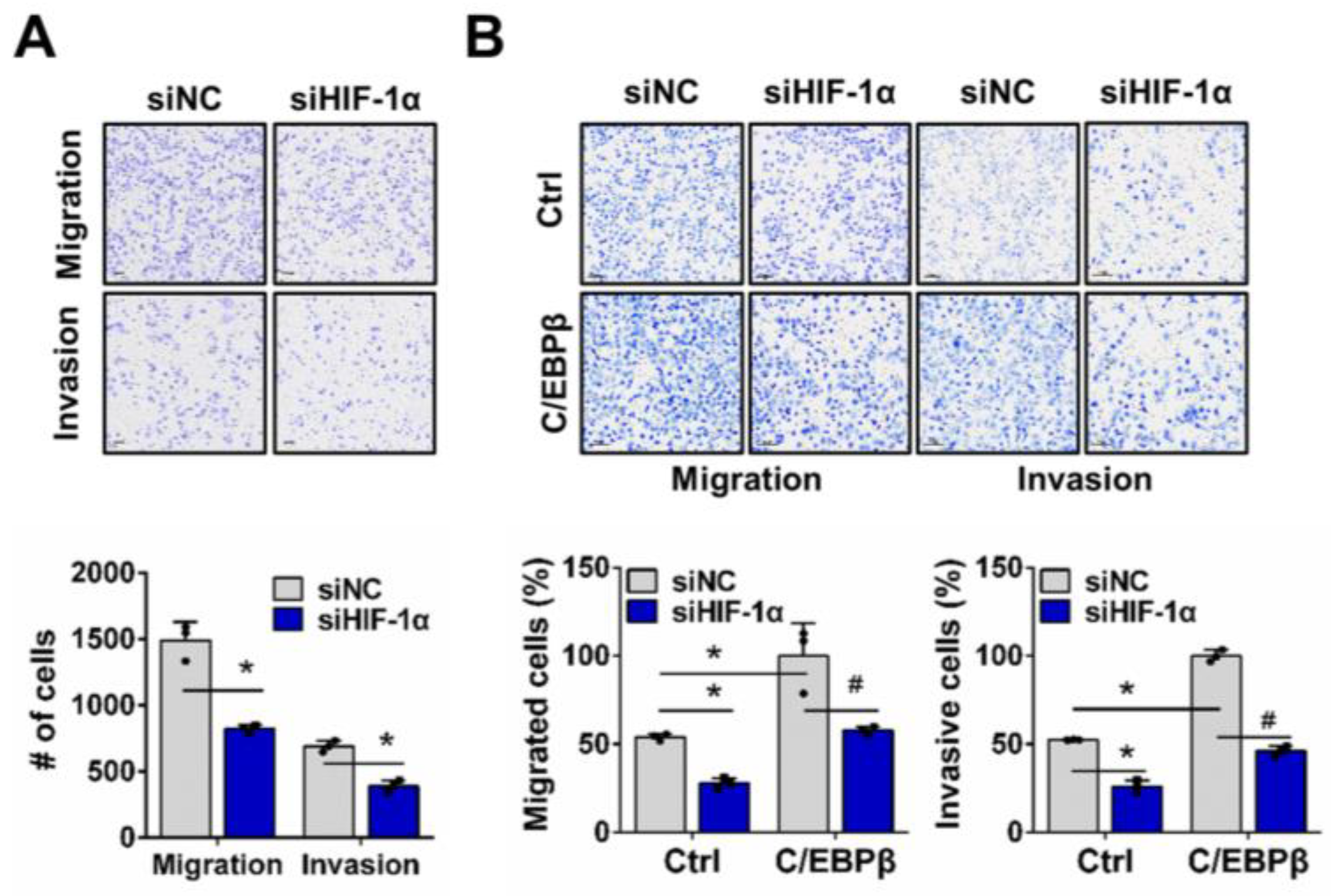
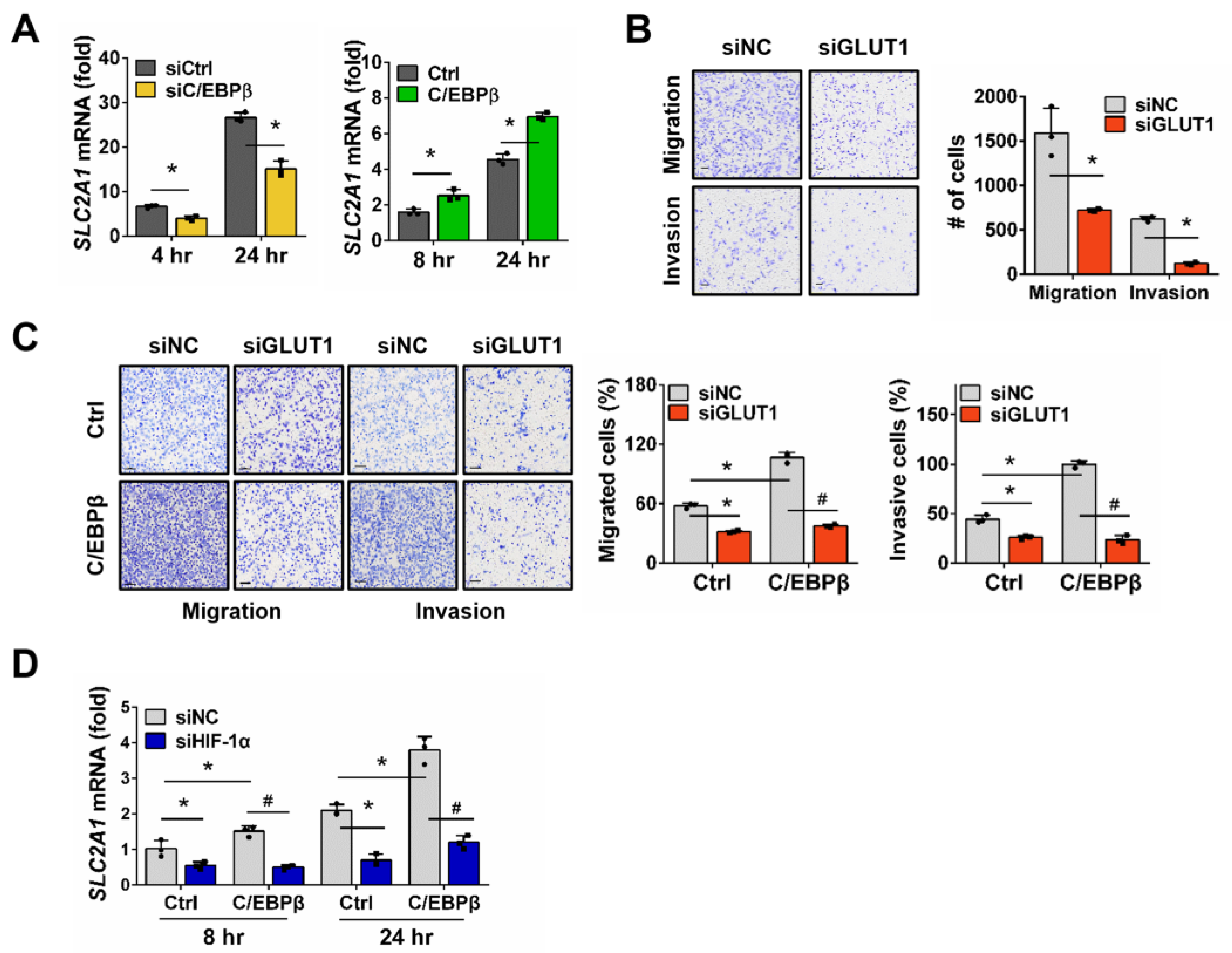
3.3. SLC2A1 Regulates Migration and Invasion of NSCLC Cells Under Hypoxic Conditions, Downstream of C/EBPβ-HIF-1α Axis
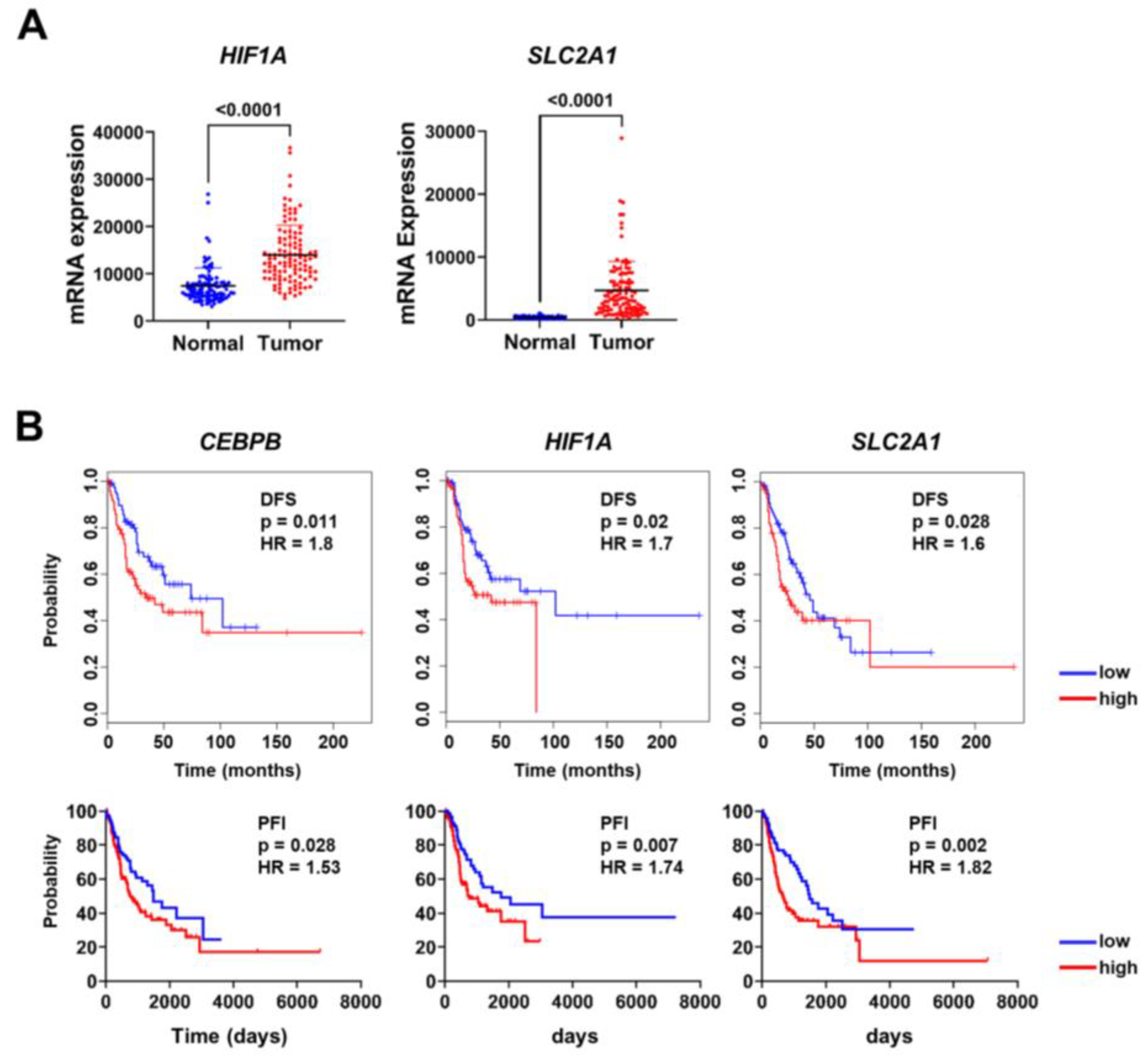
3.4. CEBPB, HIF1A, and SLC2A1 Are Associated with Prognosis of Lung Adenocarcinoma Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Thai, A.A.; Solomon, B.J.; Sequist, L.V.; Gainor, J.F.; Heist, R.S. Lung cancer. Lancet 2021, 398, 535–554. [Google Scholar] [CrossRef]
- Daly, M.E.; Singh, N.; Ismaila, N.; Antonoff, M.B.; Arenberg, D.A.; Bradley, J.; David, E.; Detterbeck, F.; Früh, M.; Gubens, M.A.; et al. Management of Stage III Non-Small-Cell Lung Cancer: ASCO Guideline. J. Clin. Oncol. 2022, 40, 1356–1384. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Temin, S.; Baker, S., Jr.; Blanchard, E.; Brahmer, J.R.; Celano, P.; Duma, N.; Ellis, P.M.; Elkins, I.B.; Haddad, R.Y.; et al. Therapy for Stage IV Non-Small-Cell Lung Cancer Without Driver Alterations: ASCO Living Guideline. J. Clin. Oncol. 2022, 40, 3323–3343. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Temin, S.; Baker, S., Jr.; Blanchard, E.; Brahmer, J.R.; Celano, P.; Duma, N.; Ellis, P.M.; Elkins, I.B.; Haddad, R.Y.; et al. Therapy for Stage IV Non-Small-Cell Lung Cancer With Driver Alterations: ASCO Living Guideline. J. Clin. Oncol. 2022, 40, 3310–3322. [Google Scholar] [CrossRef]
- Steeg, P.S. Tumor metastasis: Mechanistic insights and clinical challenges. Nat. Med. 2006, 12, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Ell, B.; Kang, Y. Transcriptional control of cancer metastasis. Trends Cell Biol. 2013, 23, 603–611. [Google Scholar] [CrossRef]
- Chen, Z.; Han, F.; Du, Y.; Shi, H.; Zhou, W. Hypoxic microenvironment in cancer: Molecular mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2023, 8, 70. [Google Scholar] [CrossRef] [PubMed]
- Frede, S.; Stockmann, C.; Freitag, P.; Fandrey, J. Bacterial lipopolysaccharide induces HIF-1 activation in human monocytes via p44/42 MAPK and NF-kappaB. Biochem. J. 2006, 396, 517–527. [Google Scholar] [CrossRef]
- Rankin, E.B.; Giaccia, A.J. Hypoxic control of metastasis. Science 2016, 352, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, L.; Li, X.F. Hypoxia and the Tumor Microenvironment. Technol. Cancer Res. Treat. 2021, 20, 15330338211036304. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia-inducible factors in physiology and medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar] [CrossRef]
- Koyasu, S.; Kobayashi, M.; Goto, Y.; Hiraoka, M.; Harada, H. Regulatory mechanisms of hypoxia-inducible factor 1 activity: Two decades of knowledge. Cancer Sci. 2018, 109, 560–571. [Google Scholar] [CrossRef]
- Kaelin, W.G., Jr.; Ratcliffe, P.J. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-inducible factors and the response to hypoxic stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Schofield, C.J.; Ratcliffe, P.J. Oxygen sensing by HIF hydroxylases. Nat. Rev. Mol. Cell Biol. 2004, 5, 343–354. [Google Scholar] [CrossRef]
- Wen, Y.; Zhou, X.; Lu, M.; He, M.; Tian, Y.; Liu, L.; Wang, M.; Tan, W.; Deng, Y.; Yang, X.; et al. Bclaf1 promotes angiogenesis by regulating HIF-1α transcription in hepatocellular carcinoma. Oncogene 2019, 38, 1845–1859. [Google Scholar] [CrossRef] [PubMed]
- Schmid, T.; Zhou, J.; Köhl, R.; Brüne, B. p300 relieves p53-evoked transcriptional repression of hypoxia-inducible factor-1 (HIF-1). Biochem. J. 2004, 380, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Carro, M.S.; Lim, W.K.; Alvarez, M.J.; Bollo, R.J.; Zhao, X.; Snyder, E.Y.; Sulman, E.P.; Anne, S.L.; Doetsch, F.; Colman, H.; et al. The transcriptional network for mesenchymal transformation of brain tumours. Nature 2010, 463, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chen, Y.; Zhan, L.; Dong, Q.; Wang, Y.; Li, X.; He, L.; Zhang, J. CEBPB-mediated upregulation of SERPINA1 promotes colorectal cancer progression by enhancing STAT3 signaling. Cell Death Discov. 2024, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Sui, Q.; Jin, X.; Shan, G.; Huang, Y.; Yi, Y.; Zeng, D.; Zhao, M.; Zhan, C.; Wang, Q. IL6-STAT3-C/EBPβ-IL6 positive feedback loop in tumor-associated macrophages promotes the EMT and metastasis of lung adenocarcinoma. J. Exp. Clin. Cancer Res. 2024, 43, 63. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Guo, W.; Qiao, B. Abnormal expression of CEBPB promotes the progression of renal cell carcinoma through regulating the generation of IL-6. Heliyon 2023, 9, e20175. [Google Scholar] [CrossRef]
- Lei, K.; Kang, S.S.; Ahn, E.H.; Chen, C.; Liao, J.; Liu, X.; Li, H.; Edgington-Mitchell, L.E.; Jin, L.; Ye, K. C/EBPβ/AEP Signaling Regulates the Oxidative Stress in Malignant Cancers, Stimulating the Metastasis. Mol. Cancer Ther. 2021, 20, 1640–1652. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.F.; Zou, Y.S.; Mendelsohn, M.; Gao, Y.; Naka, Y.; Du Yan, S.; Pinsky, D.; Stern, D. Nuclear factor interleukin 6 motifs mediate tissue-specific gene transcription in hypoxia. J. Biol. Chem. 1997, 272, 4287–4294. [Google Scholar] [CrossRef]
- Cooper, L.A.; Gutman, D.A.; Chisolm, C.; Appin, C.; Kong, J.; Rong, Y.; Kurc, T.; Van Meir, E.G.; Saltz, J.H.; Moreno, C.S.; et al. The tumor microenvironment strongly impacts master transcriptional regulators and gene expression class of glioblastoma. Am. J. Pathol. 2012, 180, 2108–2119. [Google Scholar] [CrossRef] [PubMed]
- Cassel, T.N.; Nord, M. C/EBP transcription factors in the lung epithelium. Am. J. Physiol. Lung Cell Mol. Physiol. 2003, 285, L773–L781. [Google Scholar] [CrossRef]
- Ramji, D.P.; Foka, P. CCAAT/enhancer-binding proteins: Structure, function and regulation. Biochem. J. 2002, 365, 561–575. [Google Scholar] [CrossRef]
- Lee, J.H.; Sung, J.Y.; Choi, E.K.; Yoon, H.-K.; Kang, B.R.; Hong, E.K.; Park, B.-K.; Kim, Y.-N.; Rho, S.B.; Yoon, K. C/EBPβ is a transcriptional regulator of Wee1 at the G2/M phase of the cell cycle. Cells 2019, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Elkin, M.; Vlodavsky, I. Tail vein assay of cancer metastasis. Curr. Protoc. Cell Biol. 2001, 19, 19.12.11–19.12.17. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhang, J.J.; Huang, X.Y. Mouse models for tumor metastasis. Methods Mol. Biol. 2012, 928, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Panwar, V.; Singh, A.; Bhatt, M.; Tonk, R.K.; Azizov, S.; Raza, A.S.; Sengupta, S.; Kumar, D.; Garg, M. Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal Transduct. Target. Ther. 2023, 8, 375. [Google Scholar] [CrossRef]
- Weidemann, A.; Johnson, R. Biology of HIF-1α. Cell Death Differ. 2008, 15, 621–627. [Google Scholar] [CrossRef]
- Shi, J.; Wang, H.; Feng, W.; Huang, S.; An, J.; Qiu, Y.; Wu, K. MicroRNA-130a targeting hypoxia-inducible factor 1 alpha suppresses cell metastasis and Warburg effect of NSCLC cells under hypoxia. Life Sci. 2020, 255, 117826. [Google Scholar] [CrossRef]
- Byun, Y.; Choi, Y.C.; Jeong, Y.; Lee, G.; Yoon, S.; Jeong, Y.; Yoon, J.; Baek, K. MiR-200c downregulates HIF-1α and inhibits migration of lung cancer cells. Cell Mol. Biol. Lett. 2019, 24, 28. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, Y.Q.; Shen, Y.B.; Shu, H.M.; Wang, X.J.; Zhao, C.L.; Chen, C.J. HIF-1α knockdown by miRNA decreases survivin expression and inhibits A549 cell growth in vitro and in vivo. Int. J. Mol. Med. 2013, 32, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.A.; Scafoglio, C. Heterogeneity of glucose transport in lung cancer. Biomolecules 2020, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Sun, J.; Fang, K.; Xu, J.; Yang, J.; Chunlei, D.; Gong, Y.; Ma, H. SLC2A1 boosts the resistance of non-small cell lung cancer to taxanes by stimulating the formation of EPCAM(+) cancer stem-like cells via glycolysis. Transl. Oncol. 2024, 49, 102082. [Google Scholar] [CrossRef]
- Ma, Y.; Li, J.; Xiong, C.; Sun, X.; Shen, T. Development of a prognostic model for NSCLC based on differential genes in tumour stem cells. Sci. Rep. 2024, 14, 20938. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, H.; Sun, D. SLC2A1 plays a significant prognostic role in lung adenocarcinoma and is associated with tumor immunity based on bioinformatics analysis. Ann. Transl. Med. 2022, 10, 519. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Sun, S.; Guo, L.; Song, P.; Xue, X.; Zhang, H.; Zhang, G.; Li, R.; Gao, Y.; Qiu, B.; et al. Elevated SLC2A1 Expression Correlates with Poor Prognosis in Patients with Surgically Resected Lung Adenocarcinoma: A Study Based on Immunohistochemical Analysis and Bioinformatics. DNA Cell Biol. 2020, 39, 631–644. [Google Scholar] [CrossRef]
- Mu, M.; Zhang, Q.; Li, J.; Zhao, C.; Li, X.; Chen, Z.; Sun, X.; Yu, J. USP51 facilitates colorectal cancer stemness and chemoresistance by forming a positive feed-forward loop with HIF1A. Cell Death Differ. 2023, 30, 2393–2407. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Gao, J.; Liu, R.; Liu, W.; Gao, T.; Yang, Y.; Zhang, D.; Yang, T.; Yin, X.; Yu, H. CARM1 drives triple-negative breast cancer progression by coordinating with HIF1A. Protein Cell 2024, 15, 744–765. [Google Scholar] [CrossRef]
- Ebright, R.Y.; Zachariah, M.A.; Micalizzi, D.S.; Wittner, B.S.; Niederhoffer, K.L.; Nieman, L.T.; Chirn, B.; Wiley, D.F.; Wesley, B.; Shaw, B. HIF1A signaling selectively supports proliferation of breast cancer in the brain. Nat. Commun. 2020, 11, 6311. [Google Scholar] [CrossRef]
- Sethuraman, A.; Brown, M.; Seagroves, T.N.; Wu, Z.-H.; Pfeffer, L.M.; Fan, M. SMARCE1 regulates metastatic potential of breast cancer cells through the HIF1A/PTK2 pathway. Breast Cancer Res. 2016, 18, 81. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Lu, C.-C.; Yang, L.-Y.; Wang, J.-J.; Wang, B.-S.; Cai, H.-Q.; Hao, J.-J.; Xu, X.; Cai, Y.; Zhang, Y. ANXA2 promotes esophageal cancer progression by activating MYC-HIF1A-VEGF axis. J. Exp. Clin. Cancer Res. 2018, 37, 183. [Google Scholar] [CrossRef] [PubMed]
- Tirpe, A.A.; Gulei, D.; Ciortea, S.M.; Crivii, C.; Berindan-Neagoe, I. Hypoxia: Overview on hypoxia-mediated mechanisms with a focus on the role of HIF genes. Int. J. Mol. Sci. 2019, 20, 6140. [Google Scholar] [CrossRef] [PubMed]
- Yohena, T.; Yoshino, I.; Takenaka, T.; Kameyama, T.; Ohba, T.; Kuniyoshi, Y.; Maehara, Y. Upregulation of hypoxia-inducible factor-1α mRNA and its clinical significance in non-small cell lung cancer. J. Thorac. Oncol. 2009, 4, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Hu, D.-F.; Rui, Y.; Jiang, A.-B.; Liu, Z.-L.; Huang, L.-N. Prognosis value of HIF-1α expression in patients with non-small cell lung cancer. Gene 2014, 541, 69–74. [Google Scholar] [CrossRef]
- Yee Koh, M.; Spivak-Kroizman, T.R.; Powis, G. HIF-1 regulation: Not so easy come, easy go. Trends Biochem. Sci. 2008, 33, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Schepens, B.; Tinton, S.A.; Bruynooghe, Y.; Beyaert, R.; Cornelis, S. The polypyrimidine tract-binding protein stimulates HIF-1alpha IRES-mediated translation during hypoxia. Nucleic Acids Res. 2005, 33, 6884–6894. [Google Scholar] [CrossRef] [PubMed]
- Galbán, S.; Kuwano, Y.; Pullmann, R., Jr.; Martindale, J.L.; Kim, H.H.; Lal, A.; Abdelmohsen, K.; Yang, X.; Dang, Y.; Liu, J.O.; et al. RNA-binding proteins HuR and PTB promote the translation of hypoxia-inducible factor 1alpha. Mol. Cell Biol. 2008, 28, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Lang, K.J.; Kappel, A.; Goodall, G.J. Hypoxia-inducible factor-1alpha mRNA contains an internal ribosome entry site that allows efficient translation during normoxia and hypoxia. Mol. Biol. Cell 2002, 13, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Qian, Y.; Yu, J.; Wong, C.C. Metabolic rewiring in the promotion of cancer metastasis: Mechanisms and therapeutic implications. Oncogene 2020, 39, 6139–6156. [Google Scholar] [CrossRef] [PubMed]
- Davis-Yadley, A.H.; Abbott, A.M.; Pimiento, J.M.; Chen, D.-T.; Malafa, M.P. Increased expression of the glucose transporter type 1 gene is associated with worse overall survival in resected pancreatic adenocarcinoma. Pancreas 2016, 45, 974–979. [Google Scholar] [CrossRef]
- Cai, K.; Chen, S.; Zhu, C.; Li, L.; Yu, C.; He, Z.; Sun, C. FOXD1 facilitates pancreatic cancer cell proliferation, invasion, and metastasis by regulating GLUT1-mediated aerobic glycolysis. Cell Death Dis. 2022, 13, 765. [Google Scholar] [CrossRef] [PubMed]
- Haber, R.S.; Rathan, A.; Weiser, K.R.; Pritsker, A.; Itzkowitz, S.H.; Bodian, C.; Slater, G.; Weiss, A.; Burstein, D.E. GLUT1 glucose transporter expression in colorectal carcinoma: A marker for poor prognosis. Cancer 1998, 83, 34–40. [Google Scholar] [CrossRef]
- Younes, M.; Lechago, L.V.; Lechago, J. Overexpression of the human erythrocyte glucose transporter occurs as a late event in human colorectal carcinogenesis and is associated with an increased incidence of lymph node metastases. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 1996, 2, 1151–1154. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Diagnosis | Stage | T | N | Metastatic Site |
|---|---|---|---|---|---|
| 1 | squamous cell carcinoma | IIIB | 4 | 2 | Lymph node |
| 2 | squamous cell carcinoma | IIIA | 2 | 2 | Lymph node |
| 3 | squamous cell carcinoma | IIA | 2 | 1 | Lymph node |
| 4 | adenocarcinoma | IB | 2 | 0 | Bone |
| 5 | adenocarcinoma | IIA | 2 | 1 | Lymph node |
| 6 | adenocarcinoma | IIIB | 4 | 2 | Lymph node |
| 7 | mucinous adenocarcinoma | IIIB | 2 | 2 | Lymph node |
| 8 | adenosquamous cell carcinoma | IIIA | 2 | 2 | Bone |
| 9 | mucoepidermoid | IIA | 2 | 0 | Soft tissue |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, S.H.; Lee, J.H.; Choi, E.K.; Rho, S.B.; Yoon, K. C/EBPβ Regulates HIF-1α-Driven Invasion of Non-Small-Cell Lung Cancer Cells. Biomolecules 2025, 15, 36. https://doi.org/10.3390/biom15010036
Seo SH, Lee JH, Choi EK, Rho SB, Yoon K. C/EBPβ Regulates HIF-1α-Driven Invasion of Non-Small-Cell Lung Cancer Cells. Biomolecules. 2025; 15(1):36. https://doi.org/10.3390/biom15010036
Chicago/Turabian StyleSeo, Seung Hee, Ji Hae Lee, Eun Kyung Choi, Seung Bae Rho, and Kyungsil Yoon. 2025. "C/EBPβ Regulates HIF-1α-Driven Invasion of Non-Small-Cell Lung Cancer Cells" Biomolecules 15, no. 1: 36. https://doi.org/10.3390/biom15010036
APA StyleSeo, S. H., Lee, J. H., Choi, E. K., Rho, S. B., & Yoon, K. (2025). C/EBPβ Regulates HIF-1α-Driven Invasion of Non-Small-Cell Lung Cancer Cells. Biomolecules, 15(1), 36. https://doi.org/10.3390/biom15010036