
Molecular Profiling of Axial Spondyloarthritis Patients Reveals an Association between Innate and Adaptive Cell Populations and Therapeutic Response to Tumor Necrosis Factor Inhibitors

 , , , , , ,
, , , , , ,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Samples Collection
2.2. Data Analysis
3. Results
3.1. TNFi (Adalimumab) Leads to a Decrease in Disease Activity in the Majority of axSpA Patients
3.2. Treatment with Adalimumab Had a Significant Impact on the Expression of Blood Cell Transcripts and Plasma Proteins of axSpa Patients
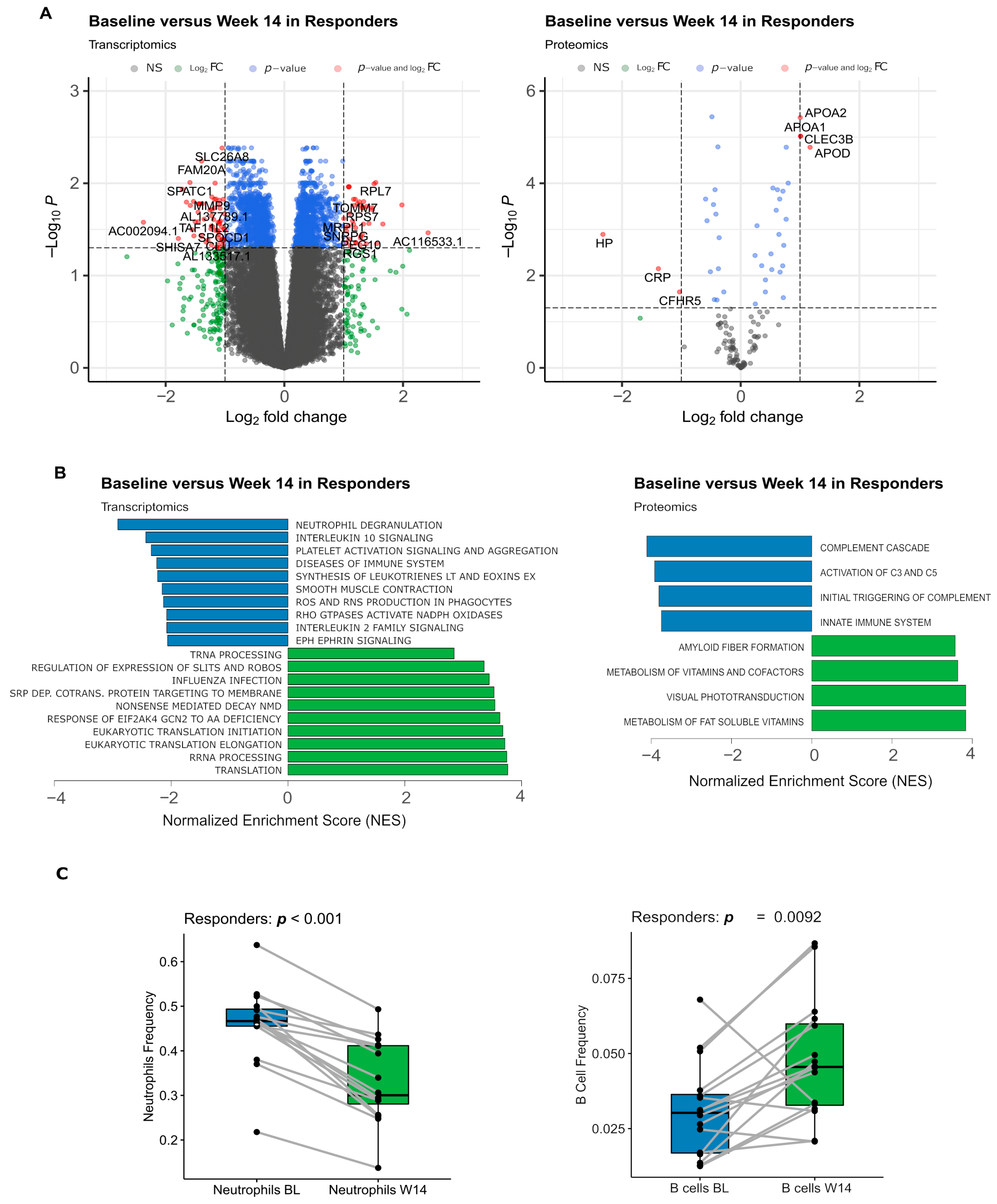
3.3. Transcripts and Proteins Varying between Baseline and Week 14 Were Associated with a Decrease in Innate Immune Activity
3.4. Markers of Inflammation Exhibited a Decrease in Plasma Levels as Early as 3 Days after Adalimumab Treatment, Observed in Both Responders and Non-Responders
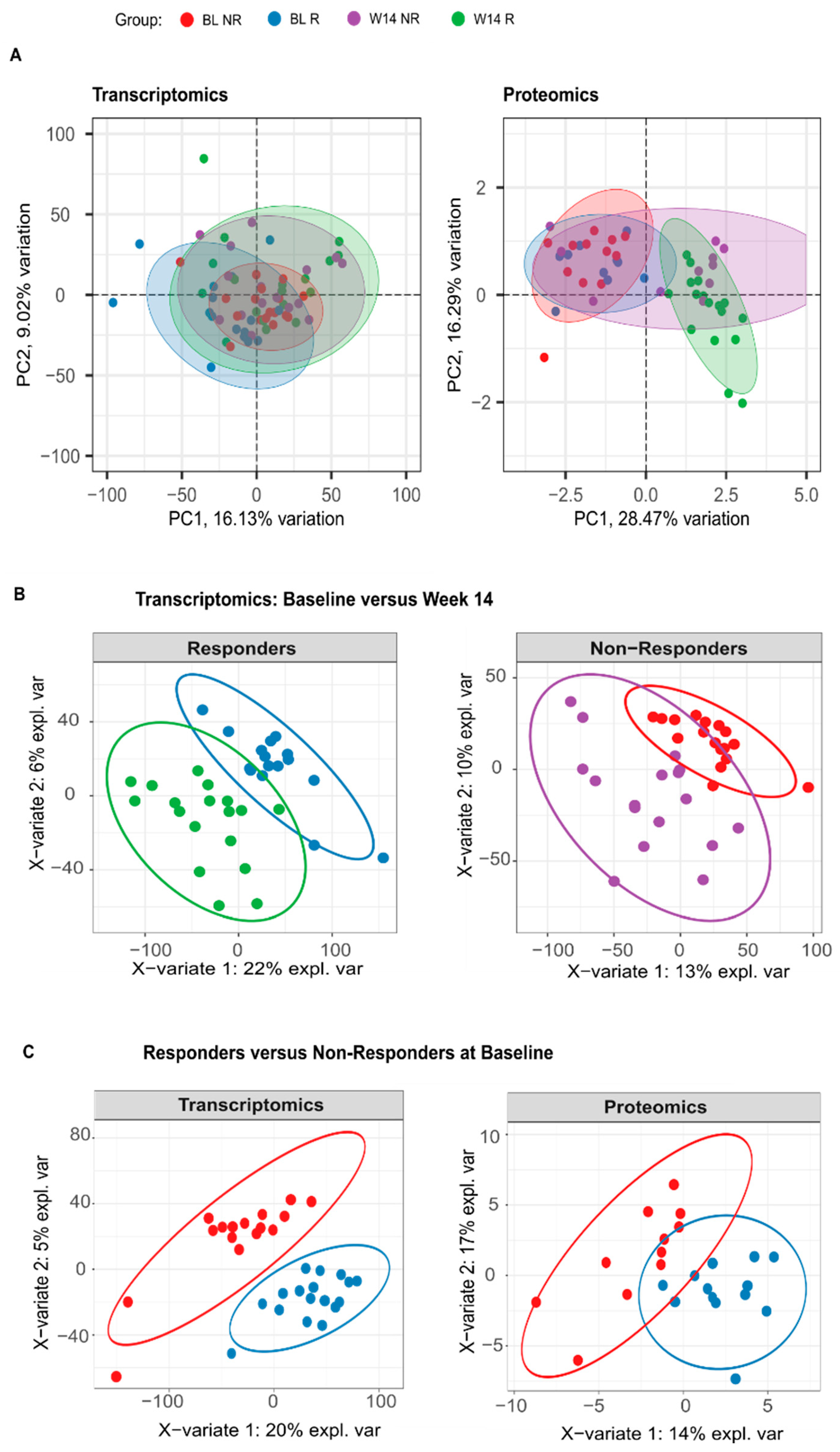
3.5. Blood Transcriptome Data at Baseline Suggest That Response to Adalimumab Stems from an Interplay between Innate and Adaptive Immunity
3.6. The Blood Transcriptome Data Enhance the Ability to Differentiate between Responders and Non-Responders at Baseline
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rudwaleit, M. New approaches to diagnosis and classification of axial and peripheral spondyloarthritis. Curr. Opin. Rheumatol. 2010, 22, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Pimentel-Santos, F.M.; CORPOREA Study Group; Mourão, A.F.; Ribeiro, C.; Costa, J.; Santos, H.; Barcelos, A.; Pinto, P.; Godinho, F.; Cruz, M.; et al. Spectrum of ankylosing spondylitis in Portugal. Development of BASDAI, BASFI, BASMI and mSASSS reference centile charts. Clin. Rheumatol. 2012, 31, 447–454. [Google Scholar] [CrossRef]
- Van Der Heijde, D.; Ramiro, S.; Landewé, R.; Baraliakos, X.; van den Bosch, F.; Sepriano, A.; Regel, A.; Ciurea, A.; Dagfinrud, H.; Dougados, M.; et al. 2016 update of the ASAS-EULAR management recommendations for axial spondyloarthritis. Ann. Rheum. Dis. 2017, 76, 978–991. [Google Scholar] [CrossRef]
- Callhoff, J.; Sieper, J.; Weiß, A.; Zink, A.; Listing, J. Efficacy of TNFα blockers in patients with ankylosing spondylitis and non-radiographic axial spondyloarthritis: A meta-analysis. Ann. Rheum. Dis. 2015, 74, 1241–1248. [Google Scholar] [CrossRef]
- Baraliakos, X.; Braun, J.; Deodhar, A.; Poddubnyy, D.; Kivitz, A.; Tahir, H.; Bosch, F.V.D.; Delicha, E.-M.; Talloczy, Z.; Fierlinger, A. Long-term efficacy and safety of secukinumab 150 mg in ankylosing spondylitis: 5-year results from the phase III MEASURE 1 extension study. RMD Open 2019, 5, e001005. [Google Scholar] [CrossRef]
- Zochling, J.; Braun, J. Management and treatment of ankylosing spondylitis. Curr. Opin. Rheumatol. 2005, 17, 418–425. [Google Scholar] [CrossRef]
- Kiltz, U.; Braun, J. Aktuelle Therapie der axialen Spondyloarthritis: Klinische Wirksamkeit. Z. Für Rheumatol. 2020, 79, 13–22. [Google Scholar] [CrossRef]
- Gentileschi, S.; Rigante, D.; Sota, J.; Lopalco, G.; Giannotta, M.G.; Emmi, G.; Di Scala, G.; Iannone, F.; Fabiani, C.; Frediani, B.; et al. Long-Term Effectiveness of Secukinumab in Patients with Axial Spondyloarthritis. Mediat. Inflamm. 2020, 2020, 6983272. [Google Scholar] [CrossRef]
- Alazmi, M.; Sari, I.; Krishnan, B.; Inman, R.D.; Haroon, N. Profiling Response to Tumor Necrosis Factor Inhibitor Treatment in Axial Spondyloarthritis. Arthritis Care Res. 2018, 70, 1393–1399. [Google Scholar] [CrossRef]
- Chen, K.; Wei, X.-Z.; Zhu, X.-D.; Bai, Y.-S.; Chen, Y.; Wang, C.-F.; Chen, Z.-Q.; Li, M. Whole-blood gene expression profiling in ankylosing spondylitis identifies novel candidate genes that may contribute to the inflammatory and tissue-destructive disease aspects. Cell. Immunol. 2013, 286, 59–64. [Google Scholar] [CrossRef]
- Blair, H.A. Secukinumab: A Review in Ankylosing Spondylitis. Drugs 2019, 79, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Dougados, M. Treat to target in axial spondyloarthritis: From its concept to its implementation. J. Autoimmun. 2020, 110, 102398. [Google Scholar] [CrossRef]
- Nell, V.P.K.; Machold, K.P.; Eberl, G.; Stamm, T.A.; Uffmann, M.; Smolen, J.S. Benefit of very early referral and very early therapy with disease-modifying anti-rheumatic drugs in patients with early rheumatoid arthritis. Rheumatology 2004, 43, 906–914. [Google Scholar] [CrossRef]
- Robinson, P.C.; Brown, M.A. The window of opportunity: A relevant concept for axial spondyloarthritis. Arthritis Res. Ther. 2014, 16, 109. [Google Scholar] [CrossRef]
- Rudwaleit, M.; Schwarzlose, S.; Hilgert, E.S.; Listing, J.; Braun, J.; Sieper, J. MRI in predicting a major clinical response to anti-tumour necrosis factor treatment in ankylosing spondylitis. Ann. Rheum. Dis. 2007, 67, 1276–1281. [Google Scholar] [CrossRef] [PubMed]
- Rudwaleit, M.; Listing, J.; Brandt, J.; Braun, J.; Sieper, J. Prediction of a major clinical response (BASDAI 50) to tumour necrosis factor blockers in ankylosing spondylitis. Ann. Rheum. Dis. 2004, 63, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Tanino, M.; Matoba, R.; Nakamura, S.; Kameda, H.; Amano, K.; Okayama, T.; Nagasawa, H.; Suzuki, K.; Matsubara, K.; Takeuchi, T. Prediction of efficacy of anti-TNF biologic agent, infliximab, for rheumatoid arthritis patients using a comprehensive transcriptome analysis of white blood cells. Biochem. Biophys. Res. Commun. 2009, 387, 261–265. [Google Scholar] [CrossRef]
- Stuhlmüller, B.; Häupl, T.; Hernandez, M.M.; Grützkau, A.; Kuban, R.-J.; Tandon, N.; Voss, J.W.; Salfeld, J.; Kinne, R.W.; Burmester, G.R. CD11c as a Transcriptional Biomarker to Predict Response to Anti-TNF Monotherapy With Adalimumab in Patients with Rheumatoid Arthritis. Clin. Pharmacol. Ther. 2010, 87, 311–321. [Google Scholar] [CrossRef]
- Thomson, T.M.; Lescarbeau, R.M.; Drubin, D.A.; Laifenfeld, D.; de Graaf, D.; Fryburg, D.A.; Littman, B.; Deehan, R.; Van Hooser, A. Blood-based identification of non-responders to anti-TNF therapy in rheumatoid arthritis. BMC Med. Genom. 2015, 8, 26. [Google Scholar] [CrossRef]
- Oswald, M.; Curran, M.E.; Lamberth, S.L.; Townsend, R.M.; Hamilton, J.D.; Chernoff, D.N.; Carulli, J.; Townsend, M.J.; Weinblatt, M.E.; Kern, M.; et al. Modular Analysis of Peripheral Blood Gene Expression in Rheumatoid Arthritis Captures Reproducible Gene Expression Changes in Tumor Necrosis Factor Responders. Arthritis Rheumatol. 2015, 67, 344–351. [Google Scholar] [CrossRef]
- Farutin, V.; Prod’homme, T.; McConnell, K.; Washburn, N.; Halvey, P.; Etzel, C.J.; Guess, J.; Duffner, J.; Getchell, K.; Meccariello, R.; et al. Molecular profiling of rheumatoid arthritis patients reveals an association between innate and adaptive cell populations and response to anti-tumor necrosis factor. Arthritis Res. Ther. 2019, 21, 216. [Google Scholar] [CrossRef]
- Rudwaleit, M.; van der Heijde, D.; Landewé, R.; Listing, J.; Akkoc, N.; Brandt, J.; Braun, J.; Chou, C.T.; Collantes-Estevez, E.; Dougados, M.; et al. The development of Assessment of SpondyloArthritis international Society classification criteria for axial spondyloarthritis (part II): Validation and final selection. Ann. Rheum. Dis. 2009, 68, 777–783, Erratum in Ann. Rheum. Dis. 2019, 78, e59. [Google Scholar] [CrossRef]
- Machado, P.M.; Cerqueira, M.; Ávila-Ribeiro, P.; Aguiar, R.; Bernardo, A.; Sepriano, A.; Águeda, A.; Cordeiro, A.; Raposo, A.; Rodrigues, A.M.; et al. Portuguese recommendations for the use of biological therapies in patients with axial spondyloarthritis—2016 update. Acta Reumatol. Port. 2017, 42, 209–218. [Google Scholar] [PubMed]
- Anderson, J.J.; Baron, G.; Van Der Heijde, D.; Felson, D.T.; Dougados, M. Ankylosing spondylitis assessment group preliminary definition of short-term improvement in ankylosing spondylitis. Arthritis Rheum. 2001, 44, 1876–1886. [Google Scholar] [CrossRef] [PubMed]
- Brandt, J.; Listing, J.; Sieper, J.; Rudwaleit, M.; van der Heijde, D.; Braun, J. Development and preselection of criteria for short term improvement after anti-TNF treatment in ankylosing spondylitis. Ann. Rheum. Dis. 2004, 63, 1438–1444. [Google Scholar] [CrossRef] [PubMed]
- Haroon, N.; Tsui, F.W.L.; O’shea, F.D.; Chiu, B.; Tsui, H.W.; Zhang, H.; Marshall, K.W.; Inman, R.D. From gene expression to serum proteins: Biomarker discovery in ankylosing spondylitis. Ann. Rheum. Dis. 2010, 69, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Ellis, J.J.; Pennisi, D.J.; Song, X.; Batra, J.; Hollis, K.; Bradbury, L.A.; Li, Z.; Kenna, T.J.; Brown, M.A. Transcriptome analysis of ankylosing spondylitis patients before and after TNF-α inhibitor therapy reveals the pathways affected. Genes Immun. 2017, 18, 184–190. [Google Scholar] [CrossRef] [PubMed]
- The Australo-Anglo-American Spondyloarthritis Consortium (TASC) Genome-wide association study of ankylosing spondylitis identifies non-MHC susceptibility loci. Nat. Genet. 2010, 42, 123–127. [CrossRef]
- Hacker, H.; Karin, M. Regulation and Function of IKK and IKK-Related Kinases. Sci. STKE 2006, 2006, re13. [Google Scholar] [CrossRef]
- White, C.R.; Garber, D.W.; Anantharamaiah, G.M. Anti-inflammatory and cholesterol-reducing properties of apolipoprotein mimetics: A review. J. Lipid Res. 2014, 55, 2007–2021. [Google Scholar] [CrossRef]
- Martineau, C.; Najyb, O.; Signor, C.; Rassart, É.; Moreau, R. Apolipoprotein D deficiency is associated to high bone turnover, low bone mass and impaired osteoblastic function in aged female mice. Metabolism 2016, 65, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Wewer, U.M.; Ibaraki, K.; Schjørring, P.; Durkin, M.E.; Young, M.F.; Albrechtsen, R. A potential role for tetranectin in mineralization during osteogenesis. J. Cell Biol. 1994, 127, 1767–1775. [Google Scholar] [CrossRef] [PubMed]
- Bruijnen, S.T.G.; Verweij, N.J.F.; van Duivenvoorde, L.M.; Bravenboer, N.; Baeten, D.L.P.; van Denderen, C.J.; E van der Horst-Bruinsma, I.; Voskuyl, A.E.; Custers, M.; van de Ven, P.M.; et al. Bone formation in ankylosing spondylitis during anti-tumour necrosis factor therapy imaged by 18F-fluoride positron emission tomography. Rheumatology 2018, 57, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Gulyás, K.; Horváth, Á.; Végh, E.; Pusztai, A.; Szentpétery, Á.; Pethö, Z.; Váncsa, A.; Bodnár, N.; Csomor, P.; Hamar, A.; et al. Effects of 1-year anti-TNF-α therapies on bone mineral density and bone biomarkers in rheumatoid arthritis and ankylosing spondylitis. Clin. Rheumatol. 2020, 39, 167–175. [Google Scholar] [CrossRef]
- Boers, N.; Michielsens, C.A.J.; Van Der Heijde, D.; Broeder, A.A.D.; Welsing, P.M.J. The effect of tumour necrosis factor inhibitors on radiographic progression in axial spondyloarthritis: A systematic literature review. Rheumatology 2019, 58, 1907–1922. [Google Scholar] [CrossRef]
- Kruhøffer, M.; Dyrskjøt, L.; Voss, T.; Lindberg, R.L.; Wyrich, R.; Thykjaer, T.; Orntoft, T.F. Isolation of Microarray-Grade Total RNA, MicroRNA, and DNA from a Single PAXgene Blood RNA Tube. J. Mol. Diagn. 2007, 9, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Khodadoust, M.S.; Liu, C.L.; Newman, A.M.; Alizadeh, A.A. Profiling Tumor Infiltrating Immune Cells with CIBERSORT. In Cancer Systems Biology; von Stechow, L., Ed.; Springer: New York, NY, USA, 2018; Volume 1711, pp. 243–259. [Google Scholar] [CrossRef]
- Abbas, A.R.; Wolslegel, K.; Seshasayee, D.; Modrusan, Z.; Clark, H.F. Deconvolution of Blood Microarray Data Identifies Cellular Activation Patterns in Systemic Lupus Erythematosus. PLoS ONE 2009, 4, e6098. [Google Scholar] [CrossRef]
- Yaari, G.; Bolen, C.R.; Thakar, J.; Kleinstein, S.H. Quantitative set analysis for gene expression: A method to quantify gene set differential expression including gene-gene correlations. Nucleic Acids Res. 2013, 41, e170. [Google Scholar] [CrossRef]
- Lewis, M.J.; Barnes, M.R.; Blighe, K.; Goldmann, K.; Rana, S.; Hackney, J.A.; Ramamoorthi, N.; John, C.R.; Watson, D.S.; Kummerfeld, S.K.; et al. Molecular Portraits of Early Rheumatoid Arthritis Identify Clinical and Treatment Response Phenotypes. Cell Rep. 2019, 28, 2455–2470.e5. [Google Scholar] [CrossRef]
- Hartmann, E.M.; Allain, F.; Gaillard, J.-C.; Pible, O.; Armengaud, J. Taking the Shortcut for High-Throughput Shotgun Proteomic Analysis of Bacteria. Methods Mol. Biol. 2014, 1197, 275–285. [Google Scholar] [CrossRef]
- Klein, G.; Mathé, C.; Biola-Clier, M.; Devineau, S.; Drouineau, E.; Hatem, E.; Marichal, L.; Alonso, B.; Gaillard, J.-C.; Lagniel, G.; et al. RNA-binding proteins are a major target of silica nanoparticles in cell extracts. Nanotoxicology 2016, 10, 1555–1564. [Google Scholar] [CrossRef]
- Washburn, M.P.; Wolters, D.; Yates, J.R.R. Large-scale analysis of the yeast proteome by multidimensional protein identification technology. Nat. Biotechnol. 2001, 19, 242–247. [Google Scholar] [CrossRef]
- Liu, H.; Sadygov, R.G.; Yates, J.R. A Model for Random Sampling and Estimation of Relative Protein Abundance in Shotgun Proteomics. Anal. Chem. 2004, 76, 4193–4201. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NR (N = 17) Mean (sd) | R (N = 18) Mean (sd) | p | |
|---|---|---|---|
| Erythrocyte Sedimentation Rate (mm/h) | |||
| Baseline | 26.1 (20.4) | 33.2 (28.5) | 0.541 |
| Week 14 | 11.9 (10.7) | 10.8 (9.10) | 0.856 |
| BL-W14 | 14.2 (16.3) | 22.3 (23.5) | 0.298 |
| C-Reactive Protein (mg/L) | |||
| Baseline | 11.3 (11.5) | 23.7 (19.7) | 0.011 |
| Week 14 | 7.42 (10.9) | 3.90 (2.78) | 0.754 |
| BL-W14 | 3.89 (4.78) | 19.8 (19.4) | <0.001 |
| BASDAI score | |||
| Baseline | 5.35 (2.63) | 6.53 (1.46) | 0.234 |
| Week 14 | 4.07 (2.18) | 1.93 (1.44) | 0.003 |
| BL-W14 | 1.28 (1.50) | 4.60 (1.81) | <0.001 |
| BASFI score | |||
| Baseline | 5.29 (2.72) | 6.71 (1.91) | 0.156 |
| Week 14 | 3.71 (2.65) | 2.55 (2.10) | 0.176 |
| BL-W14 | 1.58 (1.49) | 4.16 (2.06) | <0.001 |
| ASDAS-ESR score | |||
| Baseline | 3.23 (0.86) | 3.76 (1.08) | 0.203 |
| Week 14 | 2.27 (0.93) | 1.48 (0.50) | 0.008 |
| BL-W14 | 0.96 (0.57) | 2.28 (1.00) | <0.001 |
| ASDAS-CRP score | |||
| Baseline | 3.16 (0.75) | 4.16 (0.76) | <0.001 |
| Week 14 | 2.46 (0.75) | 1.56 (0.57) | <0.001 |
| BL-W14 | 0.70 (0.51) | 2.59 (0.94) | <0.001 |
| Age at Diagnosis (years) | |||
| 37.9 (11.3) | 34.9 (11.6) | 0.301 | |
| Disease duration (years) | |||
| 14.8 (12.7) | 13.7 (7.49) | 0.869 | |
| HLA-B27 Status | 0.010 | ||
| Absent | 10 (58.8%) | 3 (16.7%) | |
| Present | 7 (41.2%) | 15 (83.3%) | |
| Gender | 0.915 | ||
| Female | 5 (29.4%) | 5 (27.8%) | |
| Male | 12 (70.6%) | 13 (72.2%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobral, D.; Fernandes, A.F.; Bernardes, M.; Pinto, P.; Santos, H.; Lagoas-Gomes, J.; Tavares-Costa, J.; Silva, J.A.P.; Dias, J.M.; Bernardo, A.; et al. Molecular Profiling of Axial Spondyloarthritis Patients Reveals an Association between Innate and Adaptive Cell Populations and Therapeutic Response to Tumor Necrosis Factor Inhibitors. Biomolecules 2024, 14, 382. https://doi.org/10.3390/biom14030382
Sobral D, Fernandes AF, Bernardes M, Pinto P, Santos H, Lagoas-Gomes J, Tavares-Costa J, Silva JAP, Dias JM, Bernardo A, et al. Molecular Profiling of Axial Spondyloarthritis Patients Reveals an Association between Innate and Adaptive Cell Populations and Therapeutic Response to Tumor Necrosis Factor Inhibitors. Biomolecules. 2024; 14(3):382. https://doi.org/10.3390/biom14030382
Chicago/Turabian StyleSobral, Daniel, Ana Filipa Fernandes, Miguel Bernardes, Patrícia Pinto, Helena Santos, João Lagoas-Gomes, José Tavares-Costa, José A. P. Silva, João Madruga Dias, Alexandra Bernardo, and et al. 2024. "Molecular Profiling of Axial Spondyloarthritis Patients Reveals an Association between Innate and Adaptive Cell Populations and Therapeutic Response to Tumor Necrosis Factor Inhibitors" Biomolecules 14, no. 3: 382. https://doi.org/10.3390/biom14030382
APA StyleSobral, D., Fernandes, A. F., Bernardes, M., Pinto, P., Santos, H., Lagoas-Gomes, J., Tavares-Costa, J., Silva, J. A. P., Dias, J. M., Bernardo, A., Gaillard, J.-C., Armengaud, J., Benes, V., Domingues, L., Maia, S., Branco, J. C., Coelho, A. V., & Pimentel-Santos, F. M. (2024). Molecular Profiling of Axial Spondyloarthritis Patients Reveals an Association between Innate and Adaptive Cell Populations and Therapeutic Response to Tumor Necrosis Factor Inhibitors. Biomolecules, 14(3), 382. https://doi.org/10.3390/biom14030382