


Boosting Fitness Costs Associated with Antibiotic Resistance in the Gut: On the Way to Biorestoration of Susceptible Populations

 ,
, 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Stress as a Relative Concept: The Case of Intestinal Microbiota
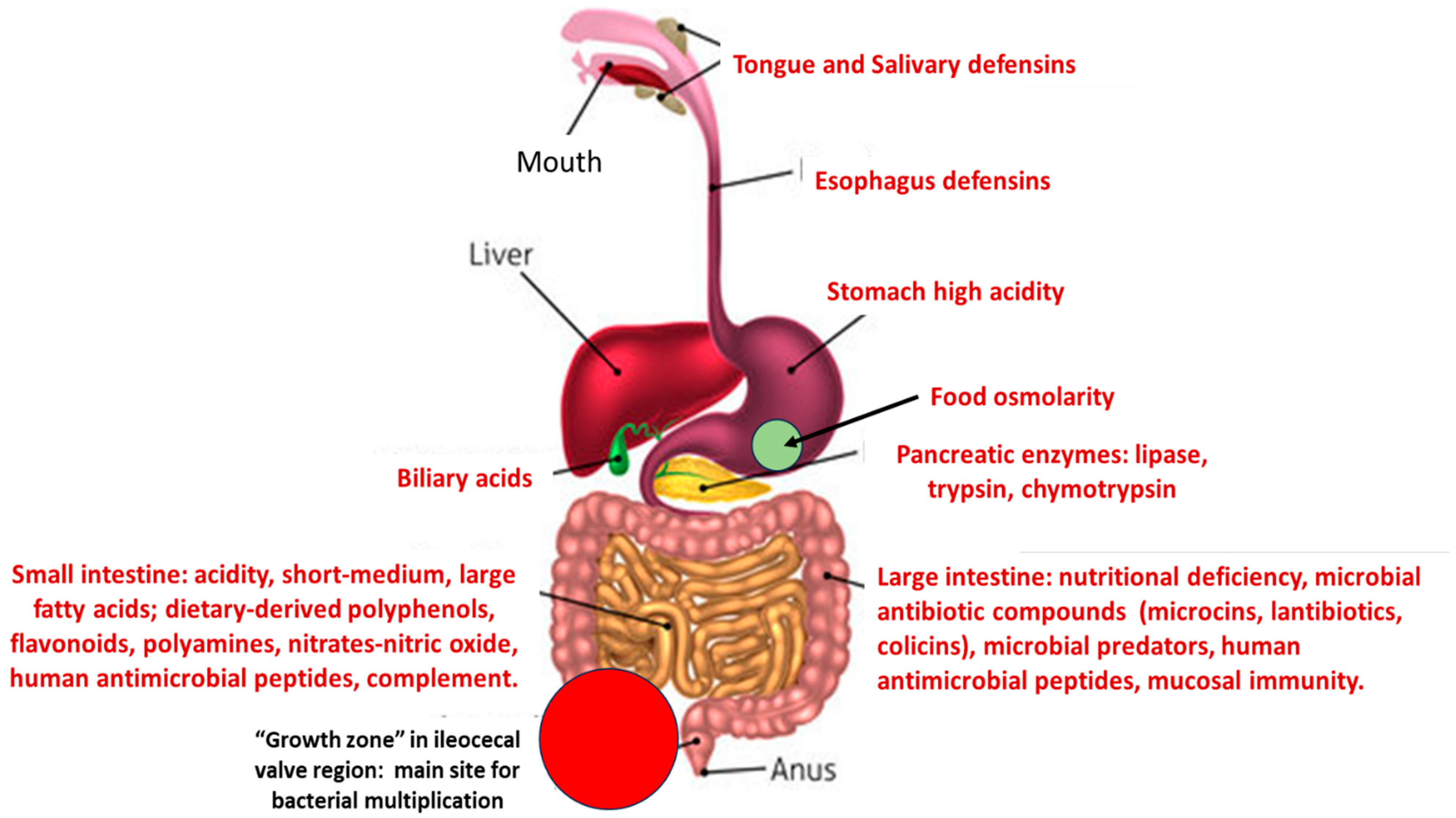
3. The Main Sources of Bacterial Stress in the Intestinal Microbiota
3.1. Acid Stress
3.2. Bile Stress
3.3. Stress by Pancreatic Enzymes
3.4. Stress by Short- and Long-Chain Fatty Acids
3.5. Stress by Dietary Compounds
3.5.1. Stress by Polyphenols
3.5.2. Stress by Polyamines
3.5.3. Nitric Oxide Stress, Osmolar Stress
3.6. Stress by Nutritional Deficiency
3.7. Stress Resulting from Microbial Interactions
3.7.1. Stress by Bacterial Antimicrobial Peptides: Microcins, Lantibiotics, Colicins
3.7.2. Stress by Bacteriophages and Microbial Predators
3.8. Stress by Inflammation and Immunity
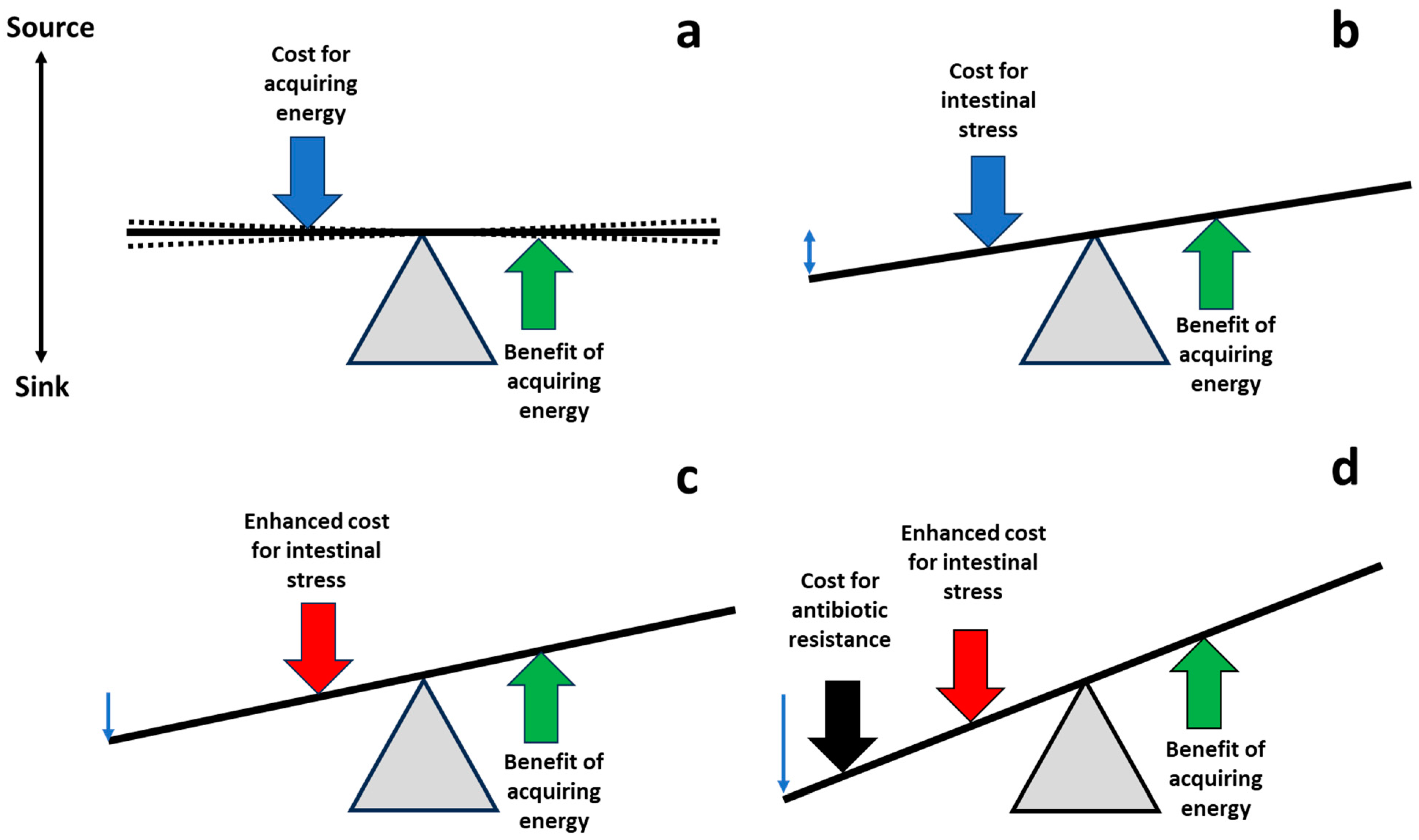
4. Modulating Intestinal Stress to Select for Antibiotic Susceptibility
5. Boosting Fitness Costs of Antibiotic-Resistant Organisms in the Gut: A Testable Hypothesis
Funding
Conflicts of Interest
References
- Davies, K.J. Adaptive homeostasis. Mol. Asp. Med. 2016, 49, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, G.E. Concluding remarks. Cold Spring Harb. Symp. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Baquero, F.; Coque, T.M.; Galán, J.C.; Martinez, J.L. The origin of niches and species in the bacterial world. Front. Microbiol. 2021, 12, 657986. [Google Scholar] [CrossRef] [PubMed]
- Tansley, A.G. The use and abuse of vegetational concepts and terms. In The Future of Nature: Documents of Global Change; Robin, L., Sörlin, S., Warde, P., Eds.; Yale University Press: New Haven, CT, USA, 2013; pp. 220–232. [Google Scholar]
- Willis, A.J. The ecosystem: An evolving concept viewed historically. Funct. Ecol. 1997, 11, 268–271. [Google Scholar] [CrossRef]
- Wollmuth, E.M.; Angert, E.R. Microbial circadian clocks: Host-microbe interplay in diel cycles. BMC Microbiol. 2023, 23, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Shahab, M.; Shahab, N. Coevolution of the human host and gut microbiome: Metagenomics of microbiota. Cureus 2022, 14, 6. [Google Scholar] [CrossRef]
- Gonzalez-Alba, J.M.; Baquero, F.; Cantón, R.; Galán, J.C. Stratified reconstruction of ancestral Escherichia coli diversification. BMC Genom. 2019, 20, 1–15. [Google Scholar] [CrossRef]
- Gordon, D.M.; Cowling, A. The distribution and genetic structure of Escherichia coli in Australian vertebrates: Host and geographic effects. Microbiology 2003, 149, 3575–3586. [Google Scholar] [CrossRef]
- Rodríguez, I.; Figueiredo, A.S.; Sousa, M.; Aracil-Gisbert, S.; Fernández-de-Bobadilla, M.D.; Lanza, V.F.; Rodríguez, C.; Zamora, J.; Loza, E.; Mingo, P.; et al. A 21-year survey of Escherichia coli from bloodstream infections [BSI] in a tertiary hospital reveals how community-hospital dynamics of B2 phylogroup clones influence local BSI rates. mSphere 2021, 6, e0086821. [Google Scholar] [CrossRef]
- Tedim, A.P.; Ruiz-Garbajosa, P.; Corander, J.; Rodríguez, C.M.; Cantón, R.; Willems, R.J.; Baquero, F.; Coque, T.M. Population biology of intestinal Enterococcus isolates from hospitalized and non-hospitalized individuals in different age groups. Appl. Environ. Microbiol. 2015, 81, 1820–1831. [Google Scholar] [CrossRef]
- Pehrsson, E.C.; Tsukayama, P.; Patel, S.; Mejía-Bautista, M.; Sosa-Soto, G.; Navarrete, K.M.; Calderon, M.; Cabrera, L.; Hoyos-Arango, W.; Bertoli, M.T.; et al. Interconnected microbiomes and resistomes in low-income human habitats. Nature 2016, 533, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Lanza, V.F.; Baquero, M.R.; del Campo, R.; Bravo-Vázquez, D.A. Microcins in Enterobacteriaceae: Peptide antimicrobials in the eco-active intestinal chemosphere. Front. Microbiol. 2019, 10, 2261. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, S.; Morosini, M.I.; Cobo, M.; Foruny, J.R.; López-Sanromán, A.; Cobo, J.; Romero, J.; Cantón, R.; del Campo, R. Gut eradication of VIM-1 producing ST9 Klebsiella oxytoca after fecal microbiota transplantation for diarrhea caused by a Clostridium difficile hypervirulent R027 strain. Diagn. Microbiol. Infect. Dis. 2016, 86, 470–471. [Google Scholar] [CrossRef] [PubMed]
- Nhu, N.T.Q.; Young, V.B. The relationship between the microbiome and antimicrobial resistance. Clin. Infect. Dis. 2023, 77 (Suppl. S6), S479–S486. [Google Scholar] [CrossRef] [PubMed]
- Woodworth, M.H.; Conrad, R.E.; Haldopoulos, M.; Pouch, S.M.; Babiker, A.; Mehta, A.K.; Sitchenko, K.L.; Wang, C.H.; Strudwick, A.; Ingersoll, J.M.; et al. Fecal microbiota transplantation promotes reduction of antimicrobial resistance by strain replacement. Sci. Transl. Med. 2023, 15, eabo2750. [Google Scholar] [CrossRef] [PubMed]
- Fallingborg, J. Intraluminal pH of the human gastrointestinal tract. Dan. Med. Bull. 1999, 46, 183–196. [Google Scholar] [PubMed]
- Dittoe, D.K.; Ricke, S.C.; Kiess, A.S. Organic acids and potential for modifying the avian gastrointestinal tract and reducing pathogens and disease. Front. Vet. Sci. 2018, 5, 216. [Google Scholar] [CrossRef]
- Foster, J.W. Escherichia coli acid resistance: Tales of an amateur acidophile. Nat. Rev. Microbiol. 2004, 2, 898–907. [Google Scholar] [CrossRef]
- Richard, H.T.; Foster, J.W. Acid resistance in Escherichia coli. Adv. Appl. Microbiol. 2003, 52, 167–186. [Google Scholar]
- Kanjee, U.; Houry, W.A. Mechanisms of acid resistance in Escherichia coli. Ann. Rev. Microbiol. 2013, 67, 65–81. [Google Scholar] [CrossRef]
- Baquero, F.; Martínez, J.L.; Sánchez, A.; Fernández-de-Bobadilla, M.D.; San-Millán, A.; Rodríguez-Beltrán, J. Bacterial subcellular architecture, structural epistasis, and antibiotic resistance. Biology 2023, 12, 640. [Google Scholar] [CrossRef] [PubMed]
- Castanie-Cornet, M.P.; Penfound, T.A.; Smith, D.; Elliott, J.F.; Foster, J.W. Control of acid resistance in Escherichia coli. J. Bacteriol. 1999, 181, 3525–3535. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Lee, I.S.; Frey, J.; Slonczewski, J.L.; Foster, J.W. Comparative analysis of extreme acid survival in Salmonella typhimurium, Shigella flexneri, and Escherichia coli. J. Bacteriol. 1995, 177, 4097–4104. [Google Scholar] [CrossRef] [PubMed]
- Castro, V.S.; Fang, Y.; Yang, X.; Stanford, K. Association of resistance to quaternary ammonium compounds and organic acids with genetic markers and their relationship to Escherichia coli serogroup. Food Microbiol. 2023, 113, 104267. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Olson, C.A.; Sastry, A.V.; Fang, X.; Phaneuf, P.V.; Chen, K.; Wu, M.; Szubin, R.; Xu, S.; Gao, Y.; et al. Adaptive laboratory evolution of Escherichia coli under acid stress. Microbiology 2020, 166, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.; Dressman, J.B. Composition and physicochemical properties of fasted-state human duodenal and jejunal fluid: A critical evaluation of the available data. J. Pharm. Sci. 2014, 103, 3398–3411. [Google Scholar] [CrossRef]
- Kurdi, P.; Kawanishi, K.; Mizutani, K.; Yokota, A. Mechanism of growth inhibition by free bile acids in lactobacilli and bifidobacteria. J. Bacteriol. 2006, 188, 1979–1986. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.; Hill, C. The interaction between bacteria and bile. FEMS Microb. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef]
- Flahaut, S.; Hartke, A.; Giard, J.C.; Auffray, Y. Alkaline stress response in Enterococcus faecalis: Adaptation, cross-protection and changes in protein synthesis. Appl. Environ. Microbiol. 1997, 63, 812–814. [Google Scholar] [CrossRef]
- Sistrunk, J.R.; Nickerson, K.P.; Chanin, R.B.; Rasko, D.A.; Faherty, C.S. Survival of the fittest: How bacterial pathogens utilize bile to enhance infection. Clin. Microbiol. Rev. 2016, 29, 819–836. [Google Scholar] [CrossRef]
- Terzi, H.A.; Kardes, H.; Atasoy, A.R.; Aykan, S.B.; Karakece, E.; Ustundag, G.H. The antibacterial effects of bilirubin on gram-negative bacterial agents of sepsis. Biomed. Res. 2016, 27, 207–209. [Google Scholar]
- Schaffner, S.H.; Lee, A.V.; Pham, M.T.; Kassaye, B.B.; Li, H.; Tallada, S.; Lis, C.; Lang, M.; Liu, Y.; Ahmed, N.; et al. Extreme acid modulates fitness trade-offs of multidrug efflux pumps MdtEF-TolC and AcrAB-TolC in Escherichia coli K-12. Appl. Environ. Microbiol. 2021, 87, e00724-21. [Google Scholar] [CrossRef] [PubMed]
- Bustos, A.Y.; de Valdez, G.F.; Fadda, S.; Taranto, M.P. New insights into bacterial bile resistance mechanisms: The role of bile salt hydrolase and its impact on human health. Food Res. Intern. 2018, 112, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Beltrán, J.; Rodríguez-Rojas, A.; Guelfo, J.R.; Couce, A.; Blázquez, J. Oxidative stress and bile salts. PLoS ONE 2012, 7, e34791. [Google Scholar]
- Isaacs, C.E.; Litov, R.E.; Marie, P.; Thormar, H. Addition of lipases to infant formulas produces antiviral and antibacterial activity. J. Nutr. Biochem. 1992, 3, 304–308. [Google Scholar] [CrossRef]
- Zhang, X.H.; Wang, Z.; Yin, B.; Wu, H.; Tang, S.; Wu, L.L.; Su, Y.N.; Lin, Y.; Liu, X.Q.; Pang, B.; et al. A complex of trypsin and chymotrypsin effectively inhibited growth of pathogenic bacteria inducing cow mastitis and showed synergistic antibacterial activity with antibiotics. Livestock Sci. 2016, 188, 25–36. [Google Scholar] [CrossRef]
- Li, Y.; Watanabe, E.; Kawashima, Y.; Plichta, D.R.; Wang, Z.; Ujike, M.; Ang, Q.Y.; Wu, R.; Furuichi, M.; Takeshita, K.; et al. Identification of trypsin-degrading commensals in the large intestine. Nature 2022, 609, 582–589. [Google Scholar] [CrossRef]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lip. Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef]
- Mitchell, M.K.; Ellermann, M. Long chain fatty acids and virulence repression in intestinal bacterial pathogens. Front. Cell. Infect. Microbiol. 2022, 12, 928503. [Google Scholar] [CrossRef] [PubMed]
- Prasun, K.; Jin-Hyung, L.; Haluk, B.; Jintae, L. Fatty acids as antibiofilm and antivirulence agents. Trends Microbiol. 2020, 28, 753–768. [Google Scholar]
- Berding, K.; Kaulmann, D.; Cryan, J.F. Food-gut microbiota interactions. In Probiotics, Advanced Food and Health Applications; Brandelli, A., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 233–256. [Google Scholar]
- Freitas, A.R.; Tedim, A.P.; Novais, C.; Ruiz-Garbajosa, P.; Werner, G.; Laverde-Gomez, J.A.; Cantón, R.; Peixe, L.; Baquero, F.; Coque, T.M. Global spread of the hyl [Efm] colonization-virulence gene in megaplasmids of the Enterococcus faecium CC17 polyclonal subcluster. Antimicrob. Agents Chemother. 2010, 54, 2660–2665. [Google Scholar] [CrossRef] [PubMed]
- Coppo, E.; Marchese, A. Antibacterial activity of polyphenols. Curr. Pharm. Biotechnol. 2014, 15, 380–390. [Google Scholar] [CrossRef]
- Manso, T.; Lores, M.; de Miguel, T. Antimicrobial activity of polyphenols and natural polyphenolic extracts on clinical isolates. Antibiotics 2021, 11, 46. [Google Scholar] [CrossRef]
- Ivanov, M.; Novović, K.; Malešević, M.; Dinić, M.; Stojković, D.; Jovčić, B.; Soković, M. Polyphenols as inhibitors of antibiotic resistant bacteria–mechanisms underlying rutin interference with bacterial virulence. Pharmaceuticals 2022, 15, 385. [Google Scholar] [CrossRef]
- Cushnie, T.T.; Lamb, A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef]
- Chambers, C.S.; Viktorová, J.; Řehořová; Biedermann, D.; Turková, L.; Macek, T.; Křen, V.; Valentová, K. Defying multidrug resistance! Modulation of related transporters by flavonoids and flavonolignans. J. Agric. Food. Chem. 2019, 68, 1763–1779. [Google Scholar] [CrossRef]
- Kemperman, R.A.; Bolca, S.; Roger, L.C.; Vaughan, E.E. Novel approaches for analysing gut microbes and dietary polyphenols: Challenges and opportunities. Microbiology 2010, 156, 3224–3231. [Google Scholar] [CrossRef]
- Zhong, W.; Tang, M.; Xie, Y.; Huang, X.; Liu, Y. Tea polyphenols inhibit the activity and toxicity of Staphylococcus aureus by destroying cell membranes and accumulating reactive oxygen species. Foodb. Pathog. Dis. 2023, 20, 294–302. [Google Scholar] [CrossRef]
- Bachrach, U.; Weinstein, A. Effect of aliphatic polyamines on growth and macromolecular syntheses in bacteria. Microbiology 1970, 60, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, M.; Borselli, D.; Brunel, J.M. Polyamine derivatives: A revival of an old neglected scaffold to fight resistant Gram-negative bacteria? Future Med. Chem. 2016, 8, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Cadelis, M.M.; Edmeades, L.R.; Chen, D.; Gill, E.S.; Fraser, K.; Rouvier, F.; Bourguet-Kondracki, M.L.; Brunel, J.M.; Copp, B.R. Investigation of naphthyl–polyamine conjugates as antimicrobials and antibiotic enhancers. Antibiotics 2023, 12, 1014. [Google Scholar] [CrossRef]
- Shah, P.; Swiatlo, E. A multifaceted role for polyamines in bacterial pathogens. Mol. Microbiol. 2008, 68, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Porrini, C.; Ramarao, N.; Tran, S.L. Dr. NO and Mr. Toxic—The versatile role of nitric oxide. Biol. Chem. 2020, 401, 547–572. [Google Scholar] [CrossRef]
- Leclerc, M.; Bedu-Ferrari, C.; Etienne-Mesmin, L.; Mariadassou, M.; Lebreuilly, L.; Tran, S.L.; Brazeau, L.; Mayeur, C.; Delmas, J.; Rué, O.; et al. Nitric oxide impacts human gut microbiota diversity and functionalities. Msystems 2021, 6, e00558-21. [Google Scholar] [CrossRef]
- Dykhuizen, R.S.; Frazer, R.; Duncan, C.; Smith, C.C.; Golden, M.; Benjamin, N.; Leifert, C. Antimicrobial effect of acidified nitrite on gut pathogens: Importance of dietary nitrate in host defense. Antimicrob. Agents Chemother. 1996, 40, 1422–1425. [Google Scholar] [CrossRef]
- Poole, R.K. Nitric oxide and nitrosative stress tolerance in bacteria. Biochem. Soc. Transact. 2005, 33, 176–180. [Google Scholar] [CrossRef]
- Tropini, C.; Moss, E.L.; Merrill, B.D.; Ng, K.M.; Higginbottom, S.K.; Casavant, E.P.; Gonzalez, C.G.; Fremin, B.; Bouley, D.M.; Elias, J.E.; et al. Transient osmotic perturbation causes long-term alteration to the gut microbiota. Cell 2018, 173, 1742–1754.e17. [Google Scholar] [CrossRef]
- Csonka, L.N. Physiological and genetic responses of bacteria to osmotic stress. Microbiol. Rev. 1989, 53, 121–147. [Google Scholar] [CrossRef]
- Lim, J.J.; Diener, C.; Wilson, J.; Valenzuela, J.J.; Baliga, N.S.; Gibbons, S.M. Growth phase estimation for abundant bacterial populations sampled longitudinally from human stool metagenomes. Nat. Commun. 2023, 14, 5682. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.; Wilson, D.J.; Feil, E.; Eyre-Walker, A. The distribution of bacterial doubling times in the wild. Proc. Biol. Sci. 2018, 285, 20180789. [Google Scholar] [CrossRef]
- Arnoldini, M.; Cremer, J.; Hwa, T. Bacterial growth, flow, and mixing shape human gut microbiota density and composition. Gut Microb. 2018, 9, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Glowacki, R.W.P.; Martens, E.C. If you eat it, or secrete it, they will grow: The expanding list of nutrients utilized by human gut bacteria. J. Bacteriol. 2020, 203, e00481-20. [Google Scholar] [CrossRef] [PubMed]
- Reese, A.T.; Pereira, F.C.; Schintlmeister, A.; Berry, D.; Wagner, M.; Hale, L.P.; Wu, A.; Jiang, S.; Durand, H.K.; Zhou, X.; et al. Microbial nitrogen limitation in the mammalian large intestine. Nat. Microbiol. 2018, 3, 1441–1450. [Google Scholar] [CrossRef]
- Brown, D.; Barton, G.; Pan, Z.; Buck, M.; Wigneshweraraj, S. Nitrogen stress response and stringent response are coupled in Escherichia coli. Nat. Commun. 2014, 5, 4115. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.N.; Mandel, M.J.; Silhavy, T.J. Escherichia coli starvation diets: Essential nutrients weigh in distinctly. J. Bacteriol. 2005, 187, 7549–7553. [Google Scholar] [CrossRef]
- Hansen, E.; Karslake, J.; Woods, R.J.; Read, A.F.; Wood, K.B. Antibiotics can be used to contain drug-resistant bacteria by maintaining sufficiently large sensitive populations. PLoS Biol. 2020, 18, e3000713. [Google Scholar] [CrossRef]
- Asensio, C.; Pérez-Díaz, J.C.; Martínez, M.C.; Baquero, F. A new family of low molecular weight antibiotics from enterobacteria. Biochem. Biophys. Res. Commun. 1976, 69, 7–14. [Google Scholar] [CrossRef]
- McAuliffe, O.; Ross, R.P.; Hill, C. Lantibiotics: Structure, biosynthesis and mode of action. FEMS Microbiol. Rev. 2001, 25, 285–308. [Google Scholar] [CrossRef]
- Guo, L.; Wambui, J.; Wang, C.; Muchaamba, F.; Fernandez-Cantos, M.V.; Broos, J.; Tasara, T.; Kuipers, O.P.; Stephan, R. Cesin, a short natural variant of nisin, displays potent antimicrobial activity against major pathogens despite lacking two C-terminal macrocycles. Microbiol Spectr. 2023, 11, e0531922. [Google Scholar] [CrossRef] [PubMed]
- Draper, L.A.; Cotter, P.D.; Hill, C.; Ross, R.P. Lantibiotic resistance. Microbiol. Mol. Biol. Rev. 2015, 79, 171–191. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Riley, M.A. The phenotypic and fitness effects of colicin resistance in Escherichia coli K-12. Evolution 1999, 53, 1019–1027. [Google Scholar]
- Sutcliffe, S.G.; Reyes, A.; Maurice, C.F. Bacteriophages playing nice: Lysogenic bacteriophage replication stable in the human gut microbiota. Iscience 2023, 26, 106007. [Google Scholar] [CrossRef]
- Manganelli, R.; Gennaro, M.L. Protecting from envelope stress: Variations on the phage-shock-protein theme. Trends Microbiol. 2017, 25, 205–216. [Google Scholar] [CrossRef] [PubMed]
- De Sordi, L.; Lourenço, M.; Debarbieux, L. The battle within: Interactions of bacteriophages and bacteria in the gastrointestinal tract. Cell Host Microb. 2019, 25, 210–218. [Google Scholar] [CrossRef]
- Chabé, M.; Lokmer, A.; Ségurel, L. Gut protozoa: Friends or foes of the human gut microbiota? Trends Parasitol. 2017, 33, 925–934. [Google Scholar] [CrossRef]
- Sun, S.; Noorian, P.; McDougald, D. Dual role of mechanisms involved in resistance to predation by protozoa and virulence to humans. Front. Microbiol. 2018, 9, 1017. [Google Scholar] [CrossRef]
- Iebba, V.; Santangelo, F.; Totino, V.; Nicoletti, M.; Gagliardi, A.; De Biase, R.V.; Cucchiara, S.; Nencioni, L.; Conte, M.P.; Schippa, S. Higher prevalence and abundance of Bdellovibrio bacteriovorus in the human gut of healthy subjects. PLoS ONE 2013, 8, e61608. [Google Scholar] [CrossRef]
- Reinoso Webb, C.; Koboziev, I.; Furr, K.L.; Grisham, M.B. Protective and pro-inflammatory roles of intestinal bacteria. Pathophysiology 2016, 2, 67–80. [Google Scholar] [CrossRef]
- Adolph, T.E.; Mayr, L.; Grabherr, F.; Tilg, H. Paneth cells and their antimicrobials in intestinal immunity. Curr. Pharmac. Des. 2018, 24, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Karlsson-Sjöberg, J.; Pütsep, K.A. CRS-peptides: Unique defense peptides of mouse Paneth cells. Mucosal Immunol. 2012, 5, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Batista Araujo, J.; Sastre de Souza, G.; Lorenzon, E.N. Indolicidin revisited: Biological activity, potential applications and perspectives of an antimicrobial peptide not yet fully explored. World J. Microbiol. Biotechnol. 2022, 38, 39. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Enée, E.; Andre-Vallee, C.; Falcone, M.; Sun, J.; Diana, J. Intestinal cathelicidin antimicrobial peptide shapes a protective neonatal gut microbiota against pancreatic autoimmunity. Gastroenterology 2022, 162, 1288–1302. [Google Scholar] [CrossRef] [PubMed]
- Sina, C.; Kemper, C.; Derer, S. The intestinal complement system in inflammatory bowel disease: Shaping intestinal barrier function. Semin. Immunol. 2018, 37, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Heesterbeek, D.A.C.; Angelier, M.L.; Harrison, R.A.; Rooijakkers, S.H.M. Complement and bacterial infections: From molecular mechanisms to therapeutic applications. J. Inn. Immun. 2018, 10, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Menendez, A.; Finlay, B.B. Defensins in the immunology of bacterial infections. Curr. Opin. Immunol. 2007, 19, 385–391. [Google Scholar] [CrossRef]
- Ernst, C.M.; Staubitz, P.; Mishra, N.N.; Yang, S.J.; Hornig, G.; Kalbacher, H.; Bayer, A.S.; Kraus, D.; Peschel, A. The bacterial defensin resistance protein MprF consists of separable domains for lipid lysinylation and antimicrobial peptide repulsion. PLoS Path. 2009, 5, e1000660. [Google Scholar] [CrossRef]
- Yang, S.J.; Mishra, N.N.; Rubio, A.; Bayer, A.S. Causal role of single nucleotide polymorphisms within the mprF gene of Staphylococcus aureus in daptomycin resistance. Antimicrob. Agents Chemother. 2013, 57, 5658–5664. [Google Scholar] [CrossRef]
- Diaz, L.; Tran, T.T.; Munita, J.M.; Miller, W.R.; Rincon, S.; Carvajal, L.P.; Arias, C.A. Whole-genome analyses of Enterococcus faecium isolates with diverse daptomycin MICs. Antimicrob. Agents Chemother. 2014, 58, 4527–4534. [Google Scholar] [CrossRef]
- López, C.; Ayala, J.A.; Bonomo, R.A.; González, L.J.; Vila, A.J. Protein determinants of dissemination and host specificity of metallo-β-lactamases. Nat. Commun. 2019, 10, 3617. [Google Scholar] [CrossRef] [PubMed]
- López, C.; Delmonti, J.; Bonomo, R.A.; Vila, A.J. Deciphering the evolution of metallo-β-lactamases: A journey from the test tube to the bacterial periplasm. J. Biol. Chem. 2022, 298, 101665. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I.; Hughes, D. Antibiotic resistance and its cost: Is it possible to reverse resistance? Nat. Rev. Microbiol. 2010, 8, 260–271. [Google Scholar] [CrossRef] [PubMed]
- San Millan, A.; MacLean, R.C. Fitness costs of plasmids: A limit to plasmid transmission. Microbiol. Spectr. 2017, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Le, V.V.H.; Olivera, C.; Spagnuolo, J.; Davies, I.G.; Rakonjac, J. In vitro synergy between sodium deoxycholate and furazolidone against enterobacteria. BMC Microbiol. 2020, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gipson, K.S.; Nickerson, K.P.; Drenkard, E.; Llanos-Chea, A.; Dogiparthi, S.K.; Lanter, B.B.; Hibbler, R.M.; Yonker, L.M.; Hurley, B.P.; Faherty, C.S. The great ESKAPE: Exploring the crossroads of bile and antibiotic resistance in bacterial pathogens. Infect Immun. 2020, 88, e00865-19. [Google Scholar] [CrossRef] [PubMed]
- Perron, G.G.; Gonzalez, A.; Buckling, A. Source–sink dynamics shape the evolution of resistance and its pleiotropic fitness cost. Proc. R. Soc. B Biol. Antib. Sci. 2007, 274, 2351–2356. [Google Scholar] [CrossRef]
- Mavri, A.; Smole Možina, S. Resistance to bile salts and sodium deoxycholate in macrolide-and fluoroquinolone-susceptible and resistant Campylobacter jejuni and Campylobacter coli strains. Microb. Drug Resist. 2013, 19, 168–174. [Google Scholar] [CrossRef]
- Capozzi, V.; Fiocco, D.; Amodio, M.L.; Gallone, A.; Spano, G. Bacterial stressors in minimally processed food. Int. J. Mol. Sci. 2009, 10, 3076–3105. [Google Scholar] [CrossRef]
- Arunachalam, E.; Ireland, W.; Yang, X.; Needleman, D. Dissecting flux balances to measure energetic costs in cell biology: Techniques and challenges. Ann. Rev. Cond. Mat. Phys. 2023, 14, 211–235. [Google Scholar] [CrossRef]
- Alonso-Del Valle, A.; León-Sampedro, R.; Rodríguez-Beltrán, J.; DelaFuente, J.; Hernández-García, M.; Ruiz-Garbajosa, P.; Cantón, R.; Peña-Miller, P. Variability of plasmid fitness effects contributes to plasmid persistence in bacterial communities. Nat. Commun. 2021, 12, 2653. [Google Scholar] [CrossRef] [PubMed]
- Reese, A.T.; Kearney, S.M. Incorporating functional trade-offs into studies of the gut microbiota. Curr. Opin. Microbiol. 2019, 50, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Connor, C.H.; Zucoloto, A.Z.; Munnoch, J.T.; Yu, I.L.; Corander, J.; Hoskisson, P.A.; McDonald, B.; McNally, A. Multidrug-resistant E. coli encoding high genetic diversity in carbohydrate metabolism genes displace commensal E. coli from the intestinal tract. PLoS Biol. 2023, 21, e3002329. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.; Jin, M.; Shao, Y.; Yin, J.; Li, H.; Chen, T.; Shi, D.; Zhou, S.; Li, J.; Yang, D. High-sugar, high-fat, and high-protein diets promote antibiotic resistance gene spreading in the mouse intestinal microbiota. Gut Microbes 2022, 14, 2022442. [Google Scholar] [CrossRef]
- Spragge, F.; Bakkeren, E.; Jahn, M.T.; Araujo, E.B.N.; Pearson, C.F.; Wang, X.; Pankhurst, L.; Cunrath, O.; Foster, K.R. Microbiome diversity protects against pathogens by nutrient blocking. Science 2023, 382, eadj3502. [Google Scholar] [CrossRef]
- Stinear, T.P.; Olden, D.C.; Johnson, P.D.; Davies, J.K.; Grayson, M.L. Enterococcal vanB resistance locus in anaerobic bacteria in human faeces. Lancet 2001, 357, 855–856. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baquero, F.; Rodríguez-Beltrán, J.; Coque, T.M.; del Campo, R. Boosting Fitness Costs Associated with Antibiotic Resistance in the Gut: On the Way to Biorestoration of Susceptible Populations. Biomolecules 2024, 14, 76. https://doi.org/10.3390/biom14010076
Baquero F, Rodríguez-Beltrán J, Coque TM, del Campo R. Boosting Fitness Costs Associated with Antibiotic Resistance in the Gut: On the Way to Biorestoration of Susceptible Populations. Biomolecules. 2024; 14(1):76. https://doi.org/10.3390/biom14010076
Chicago/Turabian StyleBaquero, Fernando, Jerónimo Rodríguez-Beltrán, Teresa M. Coque, and Rosa del Campo. 2024. "Boosting Fitness Costs Associated with Antibiotic Resistance in the Gut: On the Way to Biorestoration of Susceptible Populations" Biomolecules 14, no. 1: 76. https://doi.org/10.3390/biom14010076
APA StyleBaquero, F., Rodríguez-Beltrán, J., Coque, T. M., & del Campo, R. (2024). Boosting Fitness Costs Associated with Antibiotic Resistance in the Gut: On the Way to Biorestoration of Susceptible Populations. Biomolecules, 14(1), 76. https://doi.org/10.3390/biom14010076