
SOX2 and SOX9 Expression in Developing Postnatal Opossum (Monodelphis domestica) Cortex

 , ,
, ,  , and
, and 
Abstract
1. Introduction
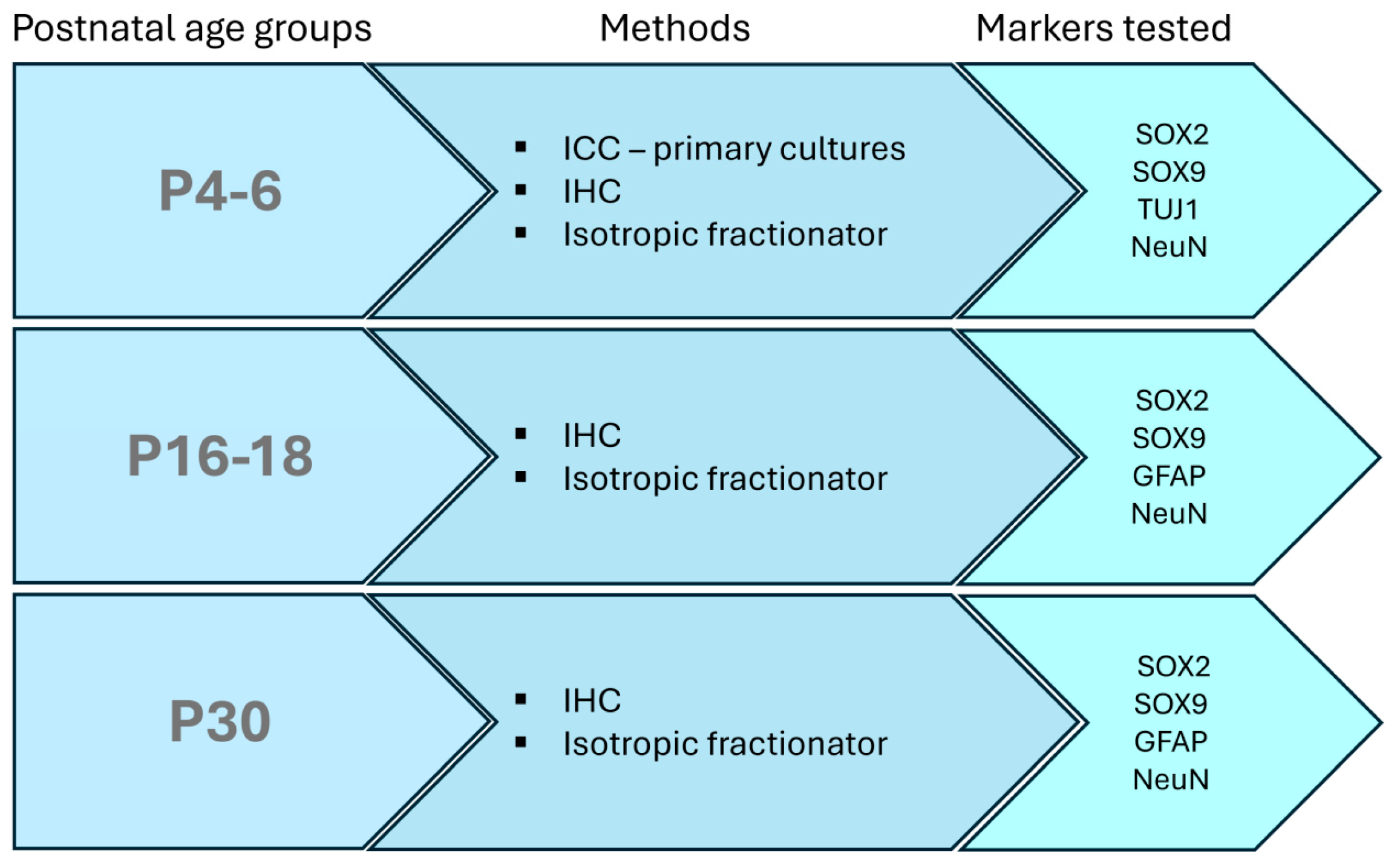
2. Materials and Methods
2.1. Animals
2.2. Primary Neuronal Cultures
2.3. Immunocytochemistry
2.4. Immunohistochemistry
2.5. Isotropic Fractionator
2.6. Imaging
2.7. Statistics
3. Results
3.1. SOX2 Expression in Developing Opossum Cortex
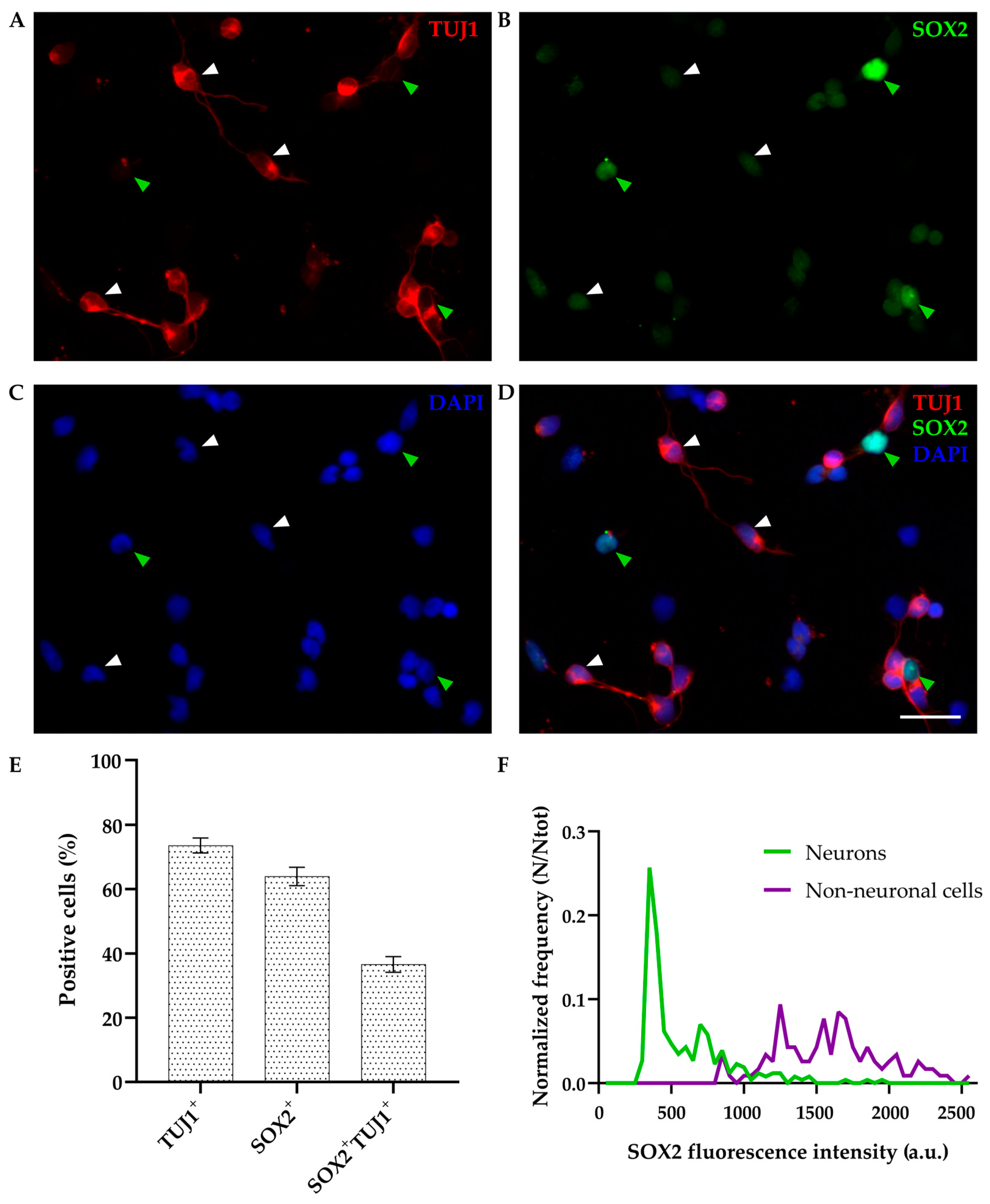
3.1.1. SOX2 Expression in Primary Dissociated Cultures of P4-6 Cortex at DIV1
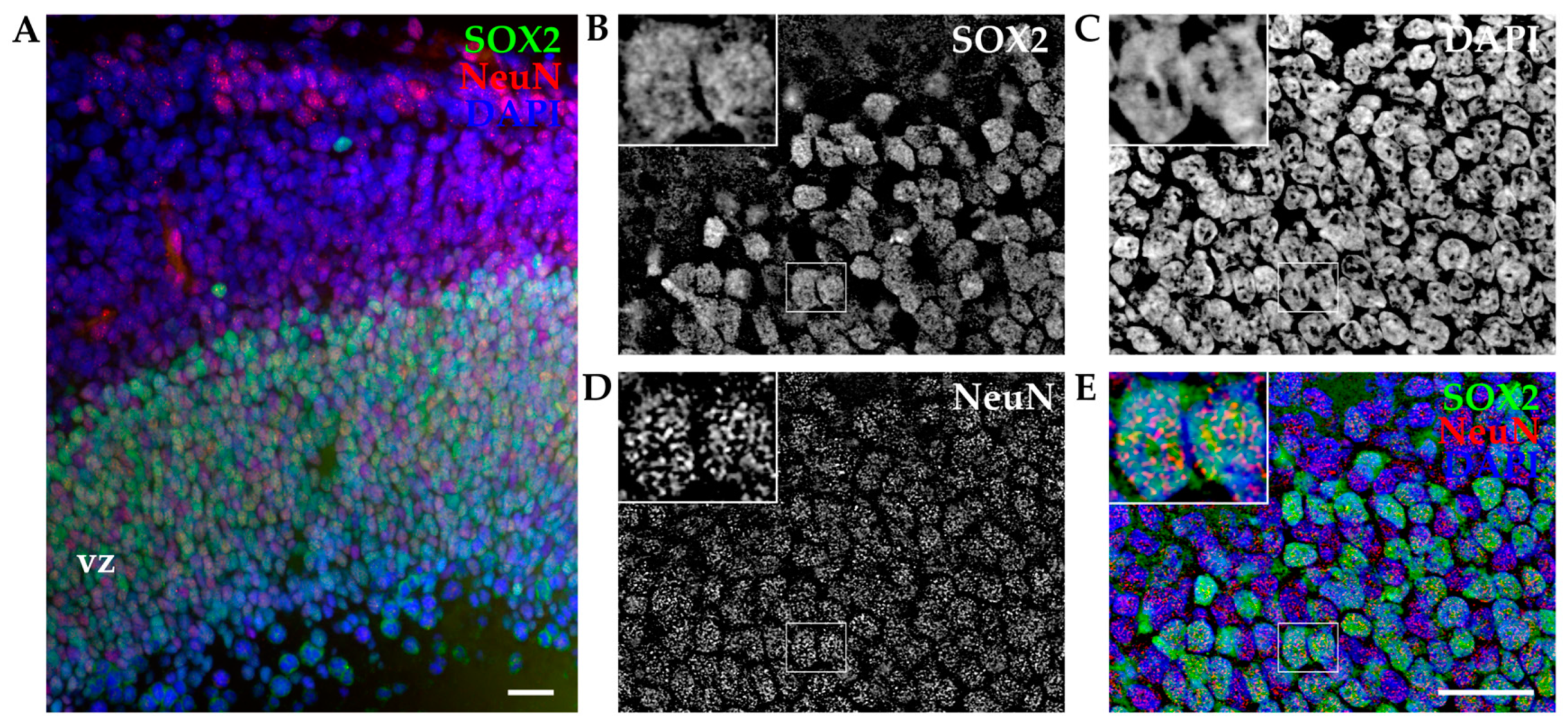
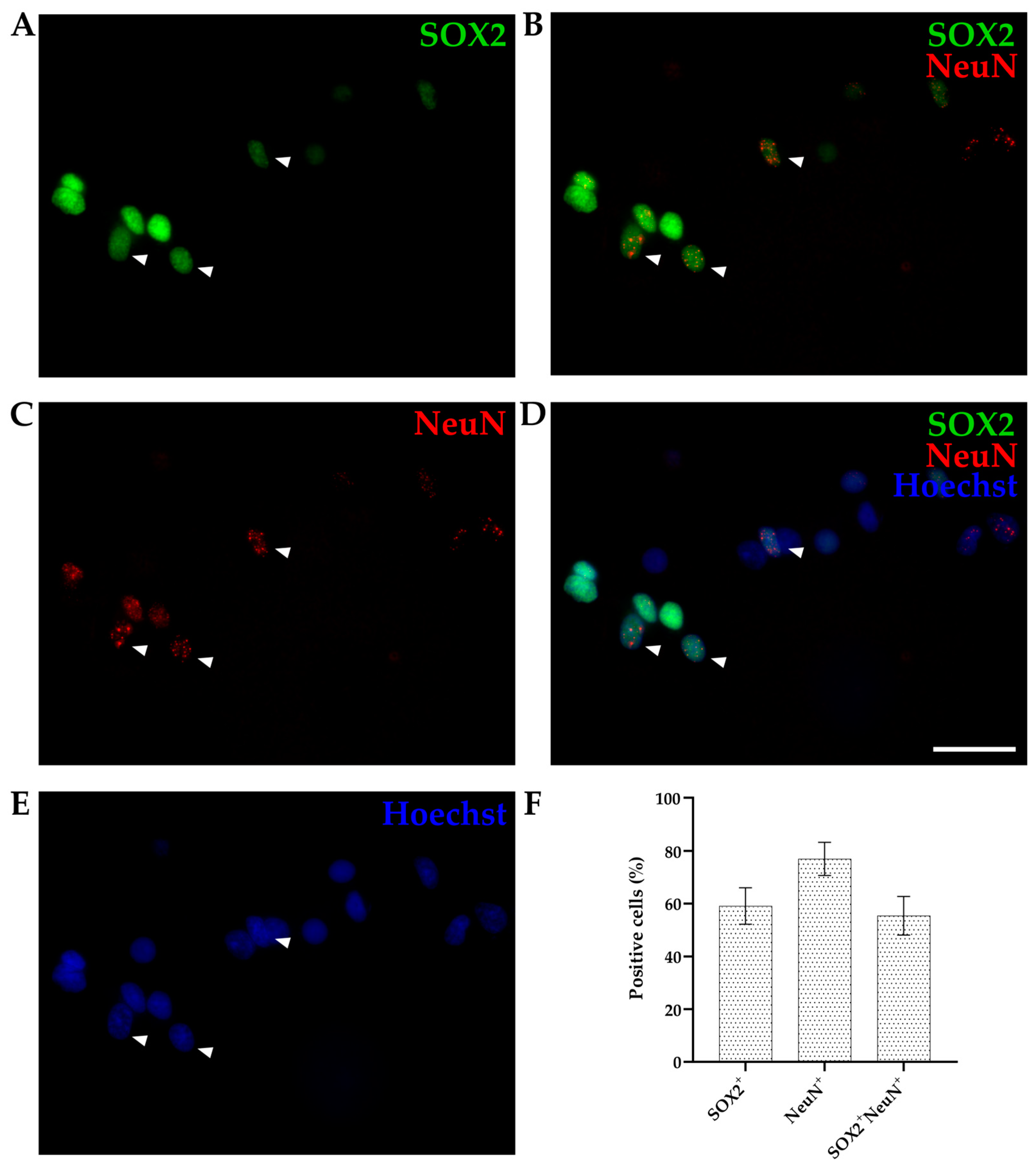
3.1.2. SOX2 Expression by IHC
3.1.3. Isotropic Fractionator
3.2. SOX9 Expression in Developing Opossum Cortex
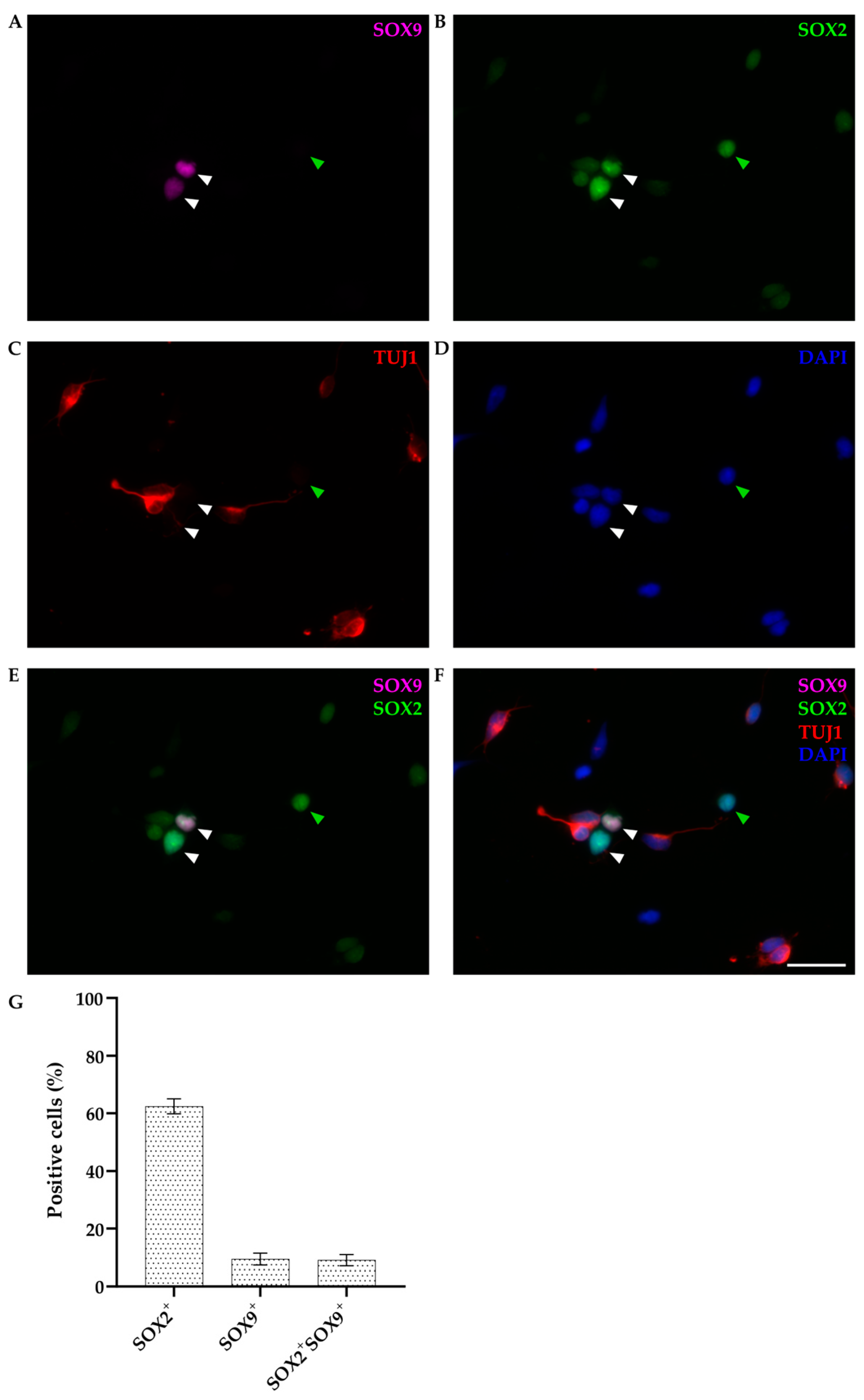
3.2.1. SOX9 Expression in Primary Dissociated Cultures of P4-6 Cortex at DIV1
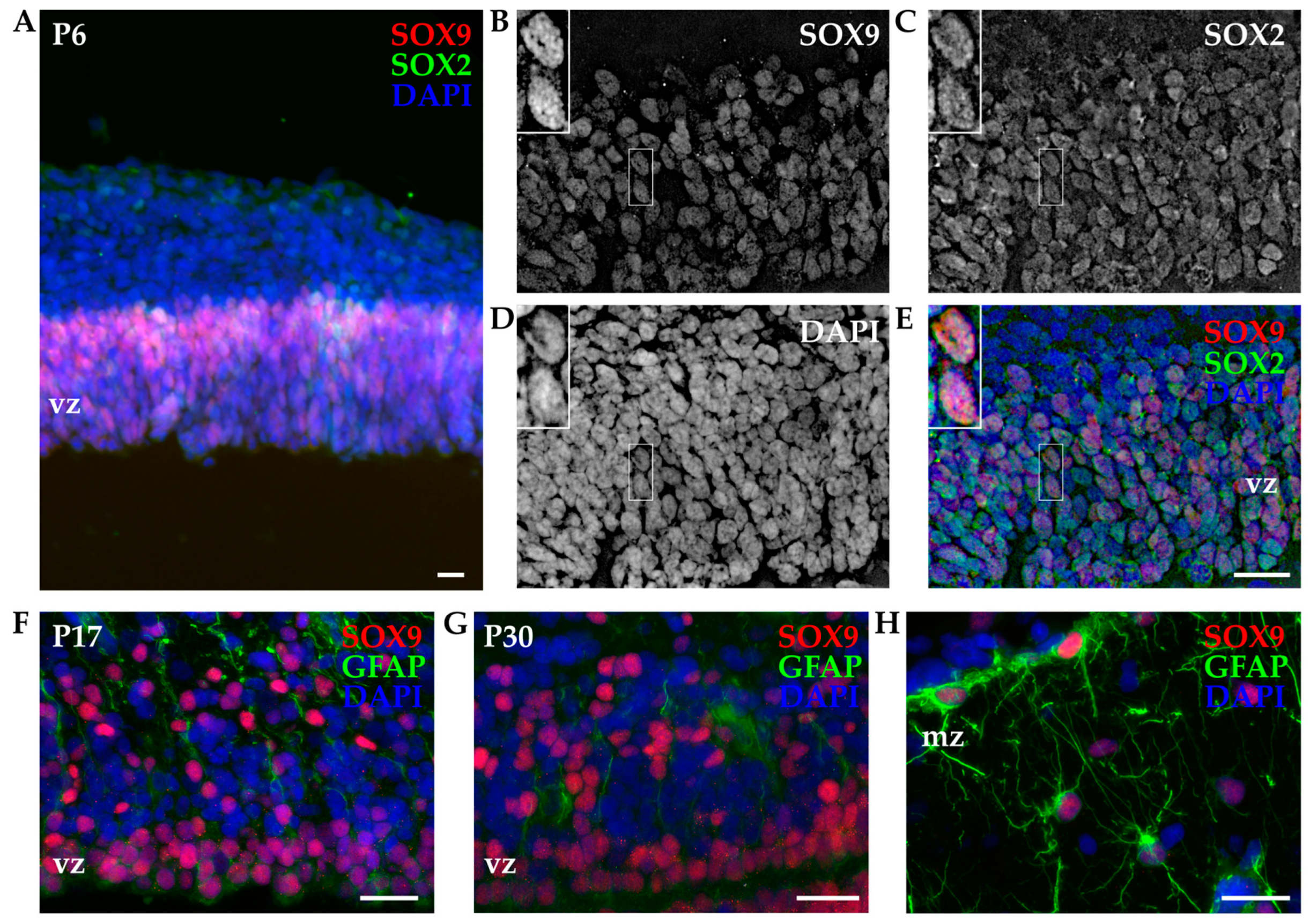
3.2.2. SOX9 Expression by IHC
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paridaen, J.T.M.L.; Huttner, W.B. Neurogenesis during Development of the Vertebrate Central Nervous System. EMBO Rep. 2014, 15, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Yanai, I. A Periodic Table of Cell Types. Development 2019, 146, dev169854. [Google Scholar] [CrossRef] [PubMed]
- Shabani, K.; Hassan, B.A. The Brain on Time: Links between Development and Neurodegeneration. Development 2023, 150, dev200397. [Google Scholar] [CrossRef]
- Schaefers, A.T.U.; Teuchert-Noodt, G. Developmental Neuroplasticity and the Origin of Neurodegenerative Diseases. World J. Biol. Psychiatry 2016, 17, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.J.; Barker, R.A. Neurodegeneration: A Failure of Neuroregeneration? Lancet 2001, 358, 1174–1176. [Google Scholar] [CrossRef] [PubMed]
- Merryweather, D.; Roach, P. The Need for Advanced Three-Dimensional Neural Models and Developing Enabling Technologies. MRS Commun. 2017, 7, 309–319. [Google Scholar] [CrossRef]
- Bonfanti, L.; Peretto, P. Adult Neurogenesis in Mammals—A Theme with Many Variations. Eur. J. Neurosci. 2011, 34, 930–950. [Google Scholar] [CrossRef]
- Herculano-Houzel, S. Mammalian Neurobiology: The Elephant (Brain) in the Room. Curr. Biol. 2022, 32, R176–R178. [Google Scholar] [CrossRef]
- Vandeberg, J.L.; Williams-Blangero, S. The Laboratory Opossum. In The UFAW Handbook on the Care and Management of Laboratory and Other Research Animals; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; pp. 246–261. ISBN 978-1-4443-1877-7. [Google Scholar]
- Kumar, S.; De Leon, E.M.; Granados, J.; Whitworth, D.J.; VandeBerg, J.L. Monodelphis domestica Induced Pluripotent Stem Cells Reveal Metatherian Pluripotency Architecture. Int. J. Mol. Sci. 2022, 23, 12623. [Google Scholar] [CrossRef]
- Samollow, P.B. The Opossum Genome: Insights and Opportunities from an Alternative Mammal. Genome Res. 2008, 18, 1199–1215. [Google Scholar] [CrossRef][Green Version]
- Nicholls, J.G.; Stewart, R.R.; Erulkar, S.D.; Saunders, N.R. Reflexes, Fictive Respiration and Cell Division in the Brain and Spinal Cord of the Newborn Opossum, Monodelphis domestica, Isolated and Maintained in Vitro. J. Exp. Biol. 1990, 152, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mladinic, M.; Muller, K.J.; Nicholls, J.G. Central Nervous System Regeneration: From Leech to Opossum. J. Physiol. 2009, 587, 2775–2782. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z.M.; Bandtlow, C.E.; Erulkar, S.D.; Schwab, M.E.; Nicholls, J.G. The Critical Period for Repair of CNS of Neonatal Opossum (Monodelphis domestica) in Culture: Correlation with Development of Glial Cells, Myelin and Growth-Inhibitory Molecules. Eur. J. Neurosci. 1995, 7, 2119–2129. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, J.; Saunders, N. Regeneration of Immature Mammalian Spinal Cord after Injury. Trends Neurosci. 1996, 19, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Puzzolo, E.; Mallamaci, A. Cortico-Cerebral Histogenesis in the Opossum Monodelphis domestica: Generation of a Hexalaminar Neocortex in the Absence of a Basal Proliferative Compartment. Neural Dev. 2010, 5, 8. [Google Scholar] [CrossRef]
- Cheung, A.F.P.; Kondo, S.; Abdel-Mannan, O.; Chodroff, R.A.; Sirey, T.M.; Bluy, L.E.; Webber, N.; De Proto, J.; Karlen, S.J.; Krubitzer, L.; et al. The Subventricular Zone Is the Developmental Milestone of a 6-Layered Neocortex: Comparisons in Metatherian and Eutherian Mammals. Cereb. Cortex 2010, 20, 1071–1081. [Google Scholar] [CrossRef]
- Molnár, Z.; Knott, G.W.; Blakemore, C.; Saunders, N.R. Development of Thalamocortical Projections in the South American Gray Short-Tailed Opossum (Monodelphis domestica). J. Comp. Neurol. 1998, 398, 491–514. [Google Scholar] [CrossRef]
- Seelke, A.M.H.; Dooley, J.C.; Krubitzer, L.A. Differential Changes in the Cellular Composition of the Developing Marsupial Brain. J. Comp. Neurol. 2013, 521, 2602–2620. [Google Scholar] [CrossRef]
- Seelke, A.M.H.; Dooley, J.C.; Krubitzer, L.A. The Cellular Composition of the Marsupial Neocortex. J. Comp. Neurol. 2014, 522, 2286–2298. [Google Scholar] [CrossRef]
- Bartkowska, K.; Gajerska, M.; Turlejski, K.; Djavadian, R.L. Expression of TrkC Receptors in the Developing Brain of the Monodelphis Opossum and Its Effect on the Development of Cortical Cells. PLoS ONE 2013, 8, e74346. [Google Scholar] [CrossRef]
- Bartkowska, K.; Aniszewska, A.; Turlejski, K.; Djavadian, R.L. Distribution and Function of TrkB Receptors in the Developing Brain of the Opossum Monodelphis domestica. Dev. Neurobiol. 2014, 74, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Tepper, B.; Bartkowska, K.; Okrasa, M.; Ngati, S.; Braszak, M.; Turlejski, K.; Djavadian, R. Downregulation of TrkC Receptors Increases Dendritic Arborization of Purkinje Cells in the Developing Cerebellum of the Opossum, Monodelphis domestica. Front. Neuroanat. 2020, 14, 56. [Google Scholar] [CrossRef] [PubMed]
- Petrović, A.; Ban, J.; Tomljanović, I.; Pongrac, M.; Ivaničić, M.; Mikašinović, S.; Mladinic, M. Establishment of Long-Term Primary Cortical Neuronal Cultures From Neonatal Opossum Monodelphis domestica. Front. Cell Neurosci. 2021, 15, 661492. [Google Scholar] [CrossRef] [PubMed]
- Ban, J.; Mladinic, M. Monodelphis domestica: A New Source of Mammalian Primary Neurons in Vitro. Neural Regen. Res. 2022, 17, 1726–1727. [Google Scholar] [CrossRef] [PubMed]
- Petrović, A.; Ban, J.; Ivaničić, M.; Tomljanović, I.; Mladinic, M. The Role of ATF3 in Neuronal Differentiation and Development of Neuronal Networks in Opossum Postnatal Cortical Cultures. Int. J. Mol. Sci. 2022, 23, 4964. [Google Scholar] [CrossRef] [PubMed]
- Chew, L.-J.; Gallo, V. The Yin and Yang of Sox Proteins: Activation and Repression in Development and Disease. J. Neurosci. Res. 2009, 87, 3277–3287. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.; Fagan, B.M.; Magness, S.T.; Hutton, S.; Taranova, O.; Hayashi, S.; McMahon, A.; Rao, M.; Pevny, L. SOX2, a Persistent Marker for Multipotential Neural Stem Cells Derived from Embryonic Stem Cells, the Embryo or the Adult. Dev. Neurosci. 2005, 26, 148–165. [Google Scholar] [CrossRef]
- Zhang, S.; Cui, W. Sox2, a Key Factor in the Regulation of Pluripotency and Neural Differentiation. World J. Stem Cells 2014, 6, 305–311. [Google Scholar] [CrossRef]
- Wegner, M. All Purpose Sox: The Many Roles of Sox Proteins in Gene Expression. Int. J. Biochem. Cell Biol. 2010, 42, 381–390. [Google Scholar] [CrossRef]
- Sarkar, A.; Hochedlinger, K. The Sox Family of Transcription Factors: Versatile Regulators of Stem and Progenitor Cell Fate. Cell Stem Cell 2013, 12, 15–30. [Google Scholar] [CrossRef]
- Graham, V.; Khudyakov, J.; Ellis, P.; Pevny, L. SOX2 Functions to Maintain Neural Progenitor Identity. Neuron 2003, 39, 749–765. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.E.; Wynn, S.L.; Sesay, A.; Cruz, C.; Cheung, M.; Gomez Gaviro, M.-V.; Booth, S.; Gao, B.; Cheah, K.S.E.; Lovell-Badge, R.; et al. SOX9 Induces and Maintains Neural Stem Cells. Nat. Neurosci. 2010, 13, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Lee, H.K.; Glasgow, S.M.; Finley, M.; Donti, T.; Gaber, Z.B.; Graham, B.H.; Foster, A.E.; Novitch, B.G.; Gronostajski, R.M.; et al. Sox9 and NFIA Coordinate a Transcriptional Regulatory Cascade during the Initiation of Gliogenesis. Neuron 2012, 74, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Cornwell, A.; Li, J.; Peng, S.; Osorio, M.J.; Aalling, N.; Wang, S.; Benraiss, A.; Lou, N.; Goldman, S.A.; et al. SOX9 Is an Astrocyte-Specific Nuclear Marker in the Adult Brain Outside the Neurogenic Regions. J. Neurosci. 2017, 37, 4493–4507. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Moreira, M.; Halbert, J.; Valloton, D.; Velten, B.; Chen, C.; Shao, Y.; Liechti, A.; Ascenção, K.; Rummel, C.; Ovchinnikova, S.; et al. Gene Expression across Mammalian Organ Development. Nature 2019, 571, 505–509. [Google Scholar] [CrossRef]
- Saunders, N.R.; Adam, E.; Reader, M.; Møllgård, K. Monodelphis domestica (Grey Short-Tailed Opossum): An Accessible Model for Studies of Early Neocortical Development. Anat. Embryol. 1989, 180, 227–236. [Google Scholar] [CrossRef]
- Herculano-Houzel, S.; Lent, R. Isotropic Fractionator: A Simple, Rapid Method for the Quantification of Total Cell and Neuron Numbers in the Brain. J. Neurosci. 2005, 25, 2518–2521. [Google Scholar] [CrossRef]
- Bahney, J.; von Bartheld, C.S. Validation of the Isotropic Fractionator: Comparison with Unbiased Stereology and DNA Extraction for Quantification of Glial Cells. J. Neurosci. Methods 2014, 222, 165–174. [Google Scholar] [CrossRef]
- Azevedo, F.A.C.; Carvalho, L.R.B.; Grinberg, L.T.; Farfel, J.M.; Ferretti, R.E.L.; Leite, R.E.P.; Jacob Filho, W.; Lent, R.; Herculano-Houzel, S. Equal Numbers of Neuronal and Nonneuronal Cells Make the Human Brain an Isometrically Scaled-up Primate Brain. J. Comp. Neurol. 2009, 513, 532–541. [Google Scholar] [CrossRef]
- Herculano-Houzel, S. The Human Brain in Numbers: A Linearly Scaled-up Primate Brain. Front. Hum. Neurosci. 2009, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, S.E.; Porfirio, J.; da Cunha, F.B.; Manger, P.R.; Tavares, W.; Pessoa, L.; Raghanti, M.A.; Sherwood, C.C.; Herculano-Houzel, S. Cellular Scaling Rules for the Brains of Marsupials: Not as “Primitive” as Expected. Brain Behav. Evol. 2017, 89, 48–63. [Google Scholar] [CrossRef] [PubMed]
- Petrović, A.; Kaur, J.; Tomljanović, I.; Nistri, A.; Mladinic, M. Pharmacological Induction of Heat Shock Protein 70 by Celastrol Protects Motoneurons from Excitotoxicity in Rat Spinal Cord in Vitro. Eur. J. Neurosci. 2019, 49, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Andreotti, J.P.; Silva, W.N.; Costa, A.C.; Picoli, C.C.; Bitencourt, F.C.O.; Coimbra-Campos, L.M.C.; Resende, R.R.; Magno, L.A.V.; Romano-Silva, M.A.; Mintz, A.; et al. Neural Stem Cell Niche Heterogeneity. Semin. Cell Dev. Biol. 2019, 95, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Malatesta, P.; Appolloni, I.; Calzolari, F. Radial Glia and Neural Stem Cells. Cell Tissue Res. 2008, 331, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Kriegstein, A.; Alvarez-Buylla, A. The Glial Nature of Embryonic and Adult Neural Stem Cells. Annu. Rev. Neurosci. 2009, 32, 149–184. [Google Scholar] [CrossRef]
- Pevny, L.H.; Nicolis, S.K. Sox2 Roles in Neural Stem Cells. Int. J. Biochem. Cell Biol. 2010, 42, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Bylund, M.; Andersson, E.; Novitch, B.G.; Muhr, J. Vertebrate Neurogenesis Is Counteracted by Sox1-3 Activity. Nat. Neurosci. 2003, 6, 1162–1168. [Google Scholar] [CrossRef]
- Hoffmann, S.A.; Hos, D.; Küspert, M.; Lang, R.A.; Lovell-Badge, R.; Wegner, M.; Reiprich, S. Stem Cell Factor Sox2 and Its Close Relative Sox3 Have Differentiation Functions in Oligodendrocytes. Development 2014, 141, 39–50. [Google Scholar] [CrossRef]
- Mercurio, S.; Serra, L.; Motta, A.; Gesuita, L.; Sanchez-Arrones, L.; Inverardi, F.; Foglio, B.; Barone, C.; Kaimakis, P.; Martynoga, B.; et al. Sox2 Acts in Thalamic Neurons to Control the Development of Retina-Thalamus-Cortex Connectivity. iScience 2019, 15, 257–273. [Google Scholar] [CrossRef]
- Cavallaro, M.; Mariani, J.; Lancini, C.; Latorre, E.; Caccia, R.; Gullo, F.; Valotta, M.; De Biasi, S.; Spinardi, L.; Ronchi, A.; et al. Impaired Generation of Mature Neurons by Neural Stem Cells from Hypomorphic Sox2 Mutants. Development 2008, 135, 541–557. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, S.; Serra, L.; Nicolis, S.K. More than Just Stem Cells: Functional Roles of the Transcription Factor Sox2 in Differentiated Glia and Neurons. Int. J. Mol. Sci. 2019, 20, 4540. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, S.; Serra, L.; Pagin, M.; Nicolis, S.K. Deconstructing Sox2 Function in Brain Development and Disease. Cells 2022, 11, 1604. [Google Scholar] [CrossRef] [PubMed]
- Gimple, R.C.; Bhargava, S.; Dixit, D.; Rich, J.N. Glioblastoma Stem Cells: Lessons from the Tumor Hierarchy in a Lethal Cancer. Genes Dev. 2019, 33, 591–609. [Google Scholar] [CrossRef]
- Wuebben, E.L.; Rizzino, A. The Dark Side of SOX2: Cancer—A Comprehensive Overview. Oncotarget 2017, 8, 44917–44943. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AGE GROUP | SOX2 | NeuN | SOX2+/NeuN+ |
|---|---|---|---|
| P16-18 | 41.38 ± 5.03% | 46.50 ± 8.25% | 33.88 ± 5.93% |
| P30 | 40.79 ± 5.09% | 49.35 ± 4.94% | 34.68 ± 6.13% 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baričević, Z.; Pongrac, M.; Ivaničić, M.; Hreščak, H.; Tomljanović, I.; Petrović, A.; Cojoc, D.; Mladinic, M.; Ban, J. SOX2 and SOX9 Expression in Developing Postnatal Opossum (Monodelphis domestica) Cortex. Biomolecules 2024, 14, 70. https://doi.org/10.3390/biom14010070
Baričević Z, Pongrac M, Ivaničić M, Hreščak H, Tomljanović I, Petrović A, Cojoc D, Mladinic M, Ban J. SOX2 and SOX9 Expression in Developing Postnatal Opossum (Monodelphis domestica) Cortex. Biomolecules. 2024; 14(1):70. https://doi.org/10.3390/biom14010070
Chicago/Turabian StyleBaričević, Zrinko, Marta Pongrac, Matea Ivaničić, Helena Hreščak, Ivana Tomljanović, Antonela Petrović, Dan Cojoc, Miranda Mladinic, and Jelena Ban. 2024. "SOX2 and SOX9 Expression in Developing Postnatal Opossum (Monodelphis domestica) Cortex" Biomolecules 14, no. 1: 70. https://doi.org/10.3390/biom14010070
APA StyleBaričević, Z., Pongrac, M., Ivaničić, M., Hreščak, H., Tomljanović, I., Petrović, A., Cojoc, D., Mladinic, M., & Ban, J. (2024). SOX2 and SOX9 Expression in Developing Postnatal Opossum (Monodelphis domestica) Cortex. Biomolecules, 14(1), 70. https://doi.org/10.3390/biom14010070