
Exploring Omega-3′s Impact on the Expression of Bone-Related Genes in Meagre (Argyrosomus regius)

 ,
,  ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Larval Culture
2.2. Feeding Protocol
2.3. Sampling
2.4. RNA Extraction
2.5. Sequencing
2.6. RNA-Seq Data Processing
2.7. Functional Enrichment Analysis
2.8. Clustering
2.9. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) for Validation
3. Results
3.1. Transcriptome Assembly and Annotation
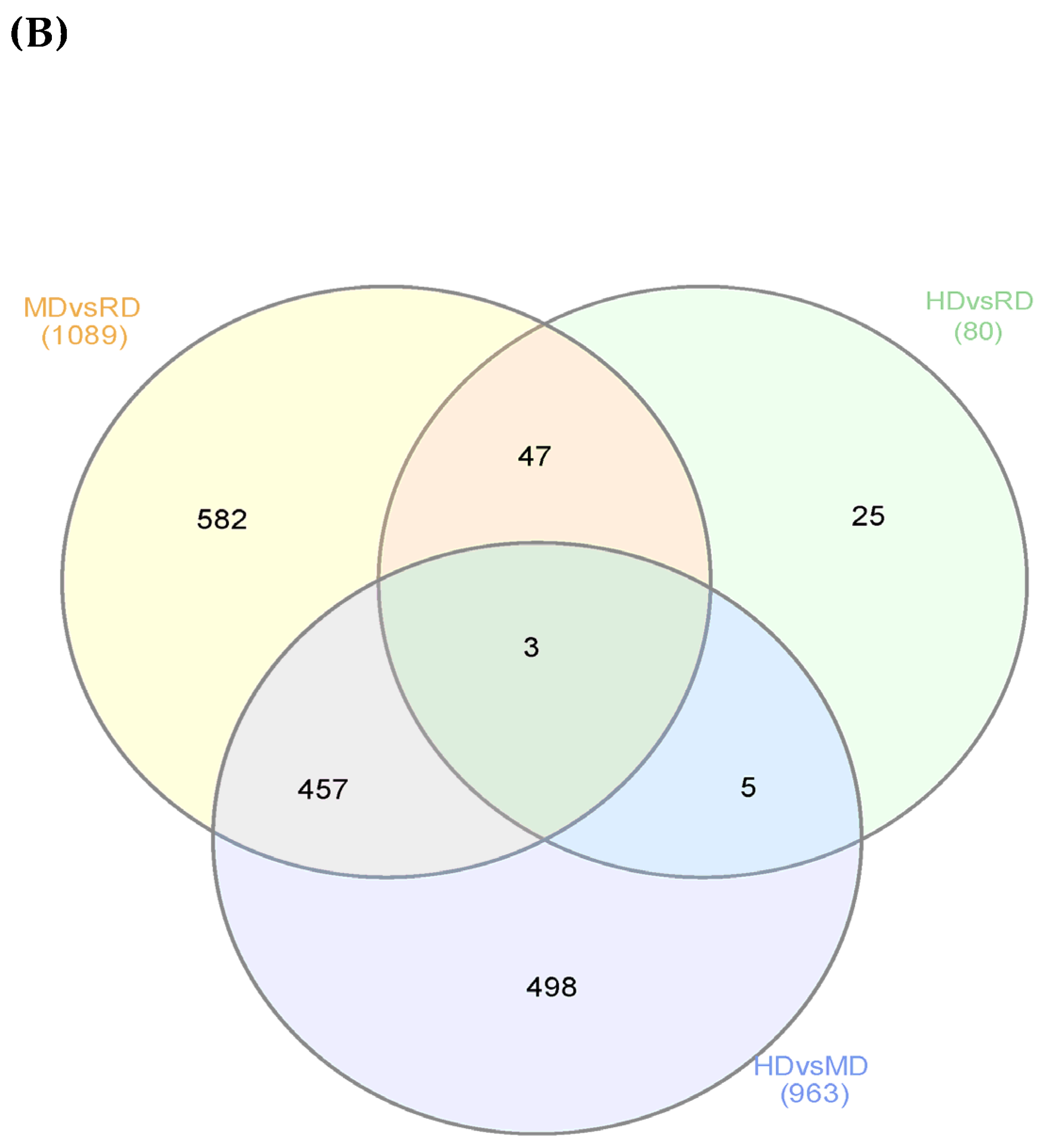
3.2. Differentially Expressed Genes (DEGs)
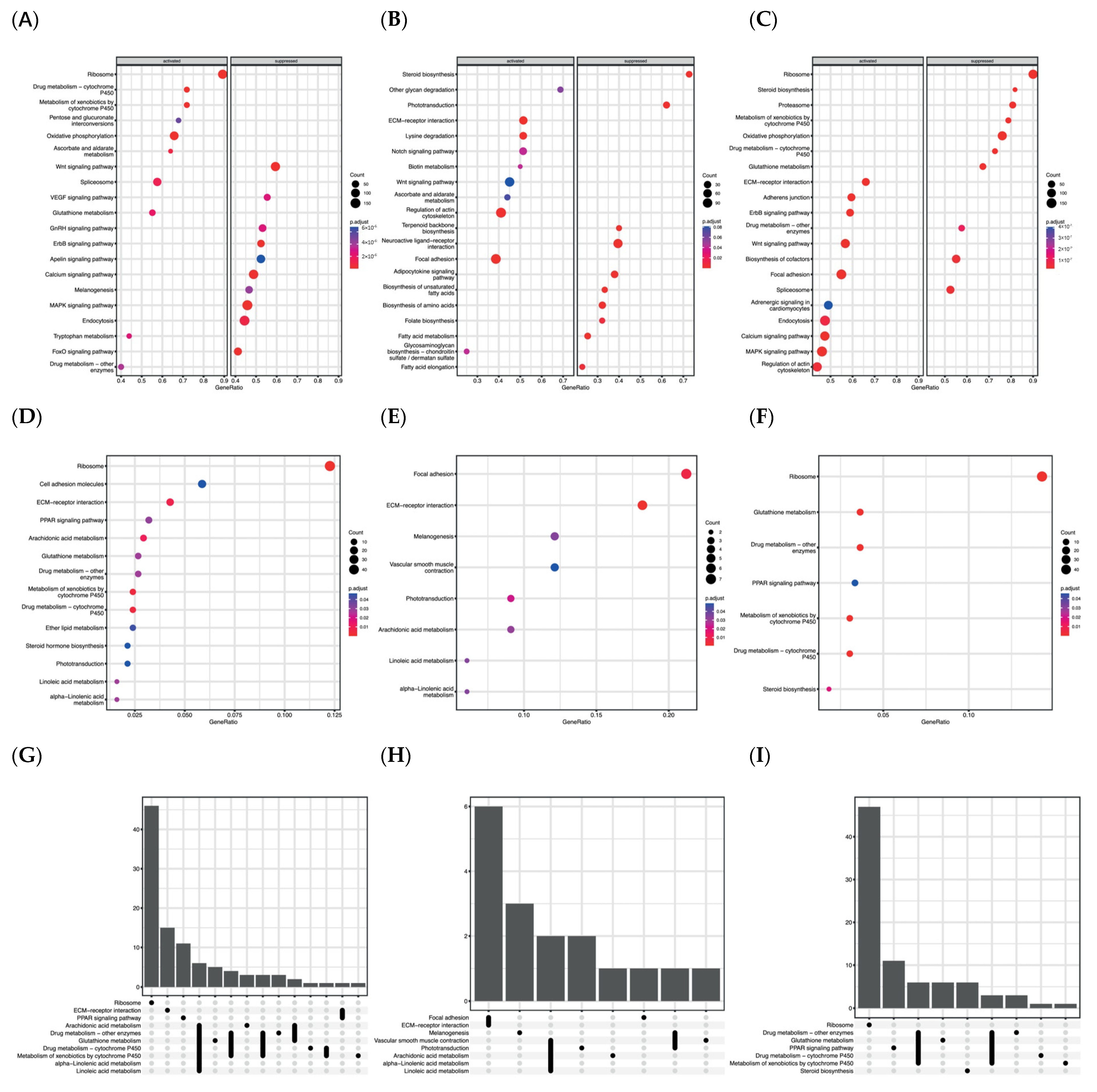
3.3. Detailed Transcriptomic Data Analysis Based on KEGG Enrichment
3.4. Detailed Transcriptomic Data Analysis Based on GO Terms Enrichment
3.5. Screening and Clustering of Interesting DEGs Retrieved among KEGG and GO Pathways
4. Discussion
4.1. Molecules That Impact Molecular Pathways Involved in Bone-Cell Differentiation
4.2. Molecules That Impact the Synthesis of ECM Components
4.3. Molecules That Impact Crucial Cellular Processes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boglione, C.; Gisbert, E.; Gavaia, P.; Witten, P.E.; Moren, M.; Fontagné, S.; Koumoundouros, G. Skeletal anomalies in reared European fish larvae and juveniles. Part 2: Main typologies, occurrences and causative factors. Rev. Aquac. 2013, 5, S99–S120. [Google Scholar] [CrossRef]
- Crockett, J.C.; Michael, J.; Coxon, F.P.; Lynne, J.; Helfrich, M.H.; Crockett, J.C.; Rogers, M.J.; Coxon, F.P.; Hocking, L.J.; Helfrich, M.H. Bone remodelling at a glance Bone Remodelling at a Glance. J. Cell. Sci. 2011, 2011, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Witten, P.E.; Hall, B.K. Teleost Skeletal Plasticity: Modulation, Adaptation, and Remodelling. Copeia 2015, 103, 727–739. [Google Scholar] [CrossRef]
- Maggio, M.; Artoni, A.; Lauretani, F.; Ruggiero, C.; Cederholm, T.; Cherubini, A.; Ceda, G.P. The effect of polyunsaturated fatty acids on bone health. Rev. Clin. Gerontol. 2011, 21, 219–232. [Google Scholar] [CrossRef]
- Harasymowicz, N.S.; Dicks, A.; Wu, C.L.; Guilak, F. Physiologic and pathologic effects of dietary free fatty acids on cells of the joint. Ann. N. Y. Acad. Sci. 2019, 1440, 36–53. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, R.C.; Moughan, P.J.; Kruger, M.C. Long-chain polyunsaturated fatty acids and the regulation of bone metabolism. Exp. Biol. Med. 2007, 232, 1275–1288. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Mandal, C.C. Omega-3 fatty acids in pathological calcification and bone health. J. Food Biochem. 2020, 44, e13333. [Google Scholar] [CrossRef]
- Watkins, B.A.; Lippman, H.E.; Le Bouteiller, L.; Li, Y.; Seifert, M.F. Bioactive fatty acids: Role in bone biology and bone cell function. Prog. Lipid Res. 2001, 40, 125–148. [Google Scholar] [CrossRef]
- Viegas, M.N.; Dias, J.; Cancela, M.L.; Laizé, V. Polyunsaturated fatty acids regulate cell proliferation, extracellular matrix mineralization and gene expression in a gilthead seabream skeletal cell line. J. Appl. Ichthyol. 2012, 28, 427–432. [Google Scholar] [CrossRef]
- Kruger, M.C.; Coetzee, M.; Haag, M.; Weiler, H. Long-chain polyunsaturated fatty acids: Selected mechanisms of action on bone. Prog. Lipid Res. 2010, 49, 438–449. [Google Scholar] [CrossRef]
- Choudhary, S.; Pilbeam, C. Prostaglandins and Bone Metabolism; Elsevier Inc.: Amsterdam, The Netherlands, 2020; ISBN 9780128148419. [Google Scholar]
- Hikiji, H.; Takato, T.; Shimizu, T.; Ishii, S. The roles of prostanoids, leukotrienes, and platelet-activating factor in bone metabolism and disease. Prog. Lipid Res. 2008, 47, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Pilbeam, C.C.; Choudhary, S.; Blackwell, K.; Raisz, L.G. Prostaglandins and Bone Metabolism; Academic Press: Cambridge, MA, USA, 2008; pp. 1235–1271. [Google Scholar]
- Kohli, P.; Levy, B.D. Resolvins and protectins: Mediating solutions to inflammation. Br. J. Pharmacol. 2009, 158, 960–971. [Google Scholar] [CrossRef] [PubMed]
- Raisz, G. The International Conference on Progress in Bone Physiologic and Pathologic Roles of Prostaglandins and Other Eicosanoids in Bone Metabolism1. J. Nutr. 1995, 125, S2024–S2027. [Google Scholar] [CrossRef] [PubMed]
- Lavado-García, J.; Roncero-Martin, R.; Moran, J.M.; Pedrera-Canal, M.; Aliaga, I.; Leal-Hernandez, O.; Rico-Martin, S.; Canal-Macias, M.L. Long-chain omega-3 polyunsaturated fatty acid dietary intake is positively associated with bone mineral density in normal and osteopenic Spanish women. PLoS ONE 2018, 13, e0190539. [Google Scholar] [CrossRef] [PubMed]
- Wiktorowska-Owczarek, A.; Berezińska, M.; Nowak, J.Z. PUFAs: Structures, metabolism and functions. Adv. Clin. Exp. Med. 2015, 24, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Azuma, M.M.; Gomes-Filho, J.E.; Ervolino, E.; Pipa, C.B.; Cardoso, C.D.; Andrada, A.C.; Kawai, T.; Cintra, L.T. Omega 3 Fatty Acids Reduce Bone Resorption While Promoting Bone Generation in Rat. J. Endod. 2017, 43, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, M.S.; Scolamacchia, M.; National, I.; Roo, J. Effects of dietary DHA and α -tocopherol on bone development, early mineralisation and oxidative stress in Sparus aurata (Linnaeus, 1758). J. Nutr. 2013, 109, 1796–1805. [Google Scholar] [CrossRef]
- El Kertaoui, N.; Lund, I.; Betancor, M.B.; Carpentier, C.; Montero, D.; Kestemont, P. Dietary DHA and ARA level and ratio affect the occurrence of skeletal anomalies in pikeperch larvae (Sander lucioperca) through a regulation of immunity and stress related gene expression. Aquaculture 2021, 544, 737060. [Google Scholar] [CrossRef]
- Berge, G.M.; Witten, P.E.; Baeverfjord, G.; Vegusdal, A.; Wadsworth, S.; Ruyter, B. Diets with different -6-3 fatty acid ratio in diets for juvenile Atlantic salmon, effects on growth, body composition, bone development and eicosanoid production. Aquaculture 2009, 296, 299–308. [Google Scholar] [CrossRef]
- Adam, A.C.; Lie, K.K.; Moren, M.; Skjærven, K.H. High dietary arachidonic acid levels induce changes in complex lipids and immune-related eicosanoids and increase levels of oxidised metabolites in zebrafish (Danio rerio). Br. J. Nutr. 2017, 117, 1075–1085. [Google Scholar] [CrossRef]
- Copeman, L.A.; Parrish, C.C.; Brown, J.A.; Harel, M. Effects of docosahexaenoic, eicosapentaenoic, and arachidonic acids on the early growth, survival, lipid composition and pigmentation of yellowtail flounder (Limanda ferruginea): A live food enrichment experiment. Aquaculture 2002, 210, 285–304. [Google Scholar] [CrossRef]
- Campoverde, C.; Estevez, A. The effect of live food enrichment with docosahexaenoic acid (22:6n-3) rich emulsions on growth, survival and fatty acid composition of meagre (Argyrosomus regius) larvae. Aquaculture 2017, 478, 16–24. [Google Scholar] [CrossRef]
- Rezek, T.C.; Watanabe, W.O.; Harel, M.; Seaton, P.J. Effects of dietary docosahexaenoic acid (22:6n-3) and arachidonic acid (20:4n-6) on the growth, survival, stress resistance and fatty acid composition in black sea bass Centropristis striata (Linnaeus 1758) larvae. Aquac. Res. 2010, 41, 1302–1314. [Google Scholar] [CrossRef]
- Gapasin, R.S.J.; Duray, M.N. Effects of DHA-enriched live food on growth, survival and incidence of opercular deformities in milkfish (Chanos chanos). Aquaculture 2001, 193, 49–63. [Google Scholar] [CrossRef]
- Roo, F.J.; Hernández-Cruz, C.M.; Socorro, J.A.; Fernández-Palacios, H.; Montero, D.; Izquierdo, M.S. Effect of DHA content in rotifers on the occurrence of skeletal deformities in red porgy Pagrus pagrus (Linnaeus, 1758). Aquaculture 2009, 287, 84–93. [Google Scholar] [CrossRef]
- Heras, J. Fish Transcriptomics: Applied to Our Understanding of Aquaculture; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar] [CrossRef]
- Berillis, P. Skeletal Deformities in Seabreams. Understanding the Genetic Origin Can Improve Production? J. Fish 2017, 11, 57. [Google Scholar] [CrossRef]
- Riera, N.; Emilio, H.; Joaquim, J.V.; Navarro, I. Gene expression analyses in malformed skeletal structures of gilthead sea bream (Sparus aurata). J. Fish Dis. 2019, 42, 1169–1180. [Google Scholar] [CrossRef]
- Vieira, F.A.; Thorne, M.A.S.; Stueber, K.; Darias, M.; Reinhardt, R.; Clark, M.S.; Gisbert, E.; Power, D.M. Comparative analysis of a teleost skeleton transcriptome provides insight into its regulation. Gen. Comp. Endocrinol. 2013, 191, 45–58. [Google Scholar] [CrossRef]
- Ytteborg, E.; Vegusdal, A.; Witten, P.E.; Berge, G.M.; Takle, H.; Østbye, T.K.; Ruyter, B. Atlantic salmon (Salmo salar) muscle precursor cells differentiate into osteoblasts in vitro: Polyunsaturated fatty acids and hyperthermia influence gene expression and differentiation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2010, 1801, 127–137. [Google Scholar] [CrossRef]
- Pfalzgraff, T.; Borges, P.; Robaina, L.; Kaushik, S.; Izquierdo, M. Essential fatty acid requirement of juvenile meagre (Argyrosomus regius). Aquaculture 2023, 572, 739532. [Google Scholar] [CrossRef]
- Duncan, N.J.; Estévez, A.; De, U.; De Gran, P.; Gairin, I.; Roo, J.; Schuchardt, D. Aquaculture Production of Meagre (Argyrosomus regius): Hatchery Techniques, Ongrowing and Market; Woodhead Publishing Limited: Sawston, UK, 2013; ISBN 9780857097460. [Google Scholar]
- Carvalho, M.; Peres, H.; Saleh, R. Requirements for N-3 Hufa of Meagre (Argyrosomus regius, Asso, 1801) Fingerlings. In Proceedings of the 18th International Symposium on Fish Nutrition and Feeding, Las Palmas, Spain, 3–7 June 2018. [Google Scholar]
- Carvalho, M.; Montero, D.; Gesto, M.; Lencina, A.; Lund, I.; Izquierdo, M. The effect of dietary n-3 LC-PUFA on the responses to acute and prolonged stress of meagre (Argyrosomus regius, Asso 1801) juveniles. Aquaculture 2019, 506, 112–118. [Google Scholar] [CrossRef]
- Ribeiro, L.; Moura, J.; Santos, M.; Colen, R.; Rodrigues, V.; Bandarra, N.; Soares, F.; Ramalho, P.; Barata, M.; Moura, P.; et al. Effect of vegetable based diets on growth, intestinal morphology, activity of intestinal enzymes and haematological stress indicators in meagre (Argyrosomus regius). Aquaculture 2015, 447, 116–128. [Google Scholar] [CrossRef]
- Cardeira, J.; Vallés, R.; Dionísio, G.; Estévez, A.; Gisbert, E.; Pousão-Ferreira, P.; Cancela, M.L.; Gavaia, P.J. Osteology of the axial and appendicular skeletons of the meagre Argyrosomus regius (Sciaenidae) and early skeletal development at two rearing facilities. J. Appl. Ichthyol. 2012, 28, 464–470. [Google Scholar] [CrossRef]
- Papadogiannis, V.; Manousaki, T.; Nousias, O.; Tsakogiannis, A.; Kristoffersen, J.B.; Mylonas, C.C.; Batargias, C.; Chatziplis, D.; Tsigenopoulos, C.S. Chromosome genome assembly for the meagre, Argyrosomus regius, reveals species adaptations and sciaenid sex-related locus evolution. Front. Genet. 2023, 13, 1081760. [Google Scholar] [CrossRef] [PubMed]
- Campoverde, C.; Milne, D.J.; Secombes, C.J.; Estévez, A.; Gisbert, E.; Andree, K.B. Gene expression analysis of the innate immune system during early rearing and weaning of meagre (Argyrosomus regius). Fish Shellfish Immunol. 2019, 94, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Patrik, D. How does gene expression clustering work ? Nat. Biotechnol. 2005, 23, 1499–1501. [Google Scholar] [CrossRef]
- Ruiz, M.A.; Betancor, M.B.; Robaina, L.; Montero, D.; Hernández-Cruz, C.M.; Izquierdo, M.S.; Rosenlund, G.; Fontanillas, R.; Caballero, M.J. Dietary combination of vitamin E, C and K affects growth, antioxidant activity, and the incidence of systemic granulomatosis in meagre (Argyrosomus regius). Aquaculture 2019, 498, 606–620. [Google Scholar] [CrossRef]
- Eriksen, E.F. Cellular mechanisms of bone remodeling. Rev. Endocr. Metab. Disord. 2010, 11, 219–227. [Google Scholar] [CrossRef]
- Kranenbarg, S.; Van Cleynenbreugel, T.; Schipper, H.; Van Leeuwen, J. Adaptive bone formation in acellular vertebrae of sea bass (Dicentrarchus labrax L.). J. Exp. Biol. 2005, 208, 3493–3502. [Google Scholar] [CrossRef]
- Witten, P.E.; Huysseune, A. A comparative view on mechanisms and functions of skeletal remodelling in teleost fish, with special emphasis on osteoclasts and their function. Biol. Rev. 2009, 84, 315–346. [Google Scholar] [CrossRef]
- Slongo, T. Skeletal System. In Pediatric Surgery Digest; Springer: Cham, Switzerland, 2009; pp. 735–800. [Google Scholar] [CrossRef]
- Long, F.; Ornitz, D.M. Development of the endochondral skeleton. Cold Spring Harb. Perspect. Biol. 2013, 5, a008334. [Google Scholar] [CrossRef] [PubMed]
- Carter, D.R.; Beaupré, G.S. Perichondral and periosteal ossification. In Skeletal Function and Form; Cambridge University Press: Cambridge, UK, 2010; ISBN 9780511574993. [Google Scholar]
- Hallett, S.A.; Ono, W.; Ono, N. The hypertrophic chondrocyte: To be or not to be. Histol. Histopathol. 2021, 36, 1021–1036. [Google Scholar] [CrossRef] [PubMed]
- Mackie, E.J.; Ahmed, Y.A.; Tatarczuch, L.; Chen, K.; Mirams, M. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell Biol. 2008, 40, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.A.; Martin, T.J. Coupling the activities of bone formation and resorption: A multitude of signals within the basic multicellular unit. BoneKEy Rep. 2014, 481, 1–10. [Google Scholar] [CrossRef]
- Florencio-silva, R.; Rodrigues, G.; Sasso-cerri, E.; Simões, M.J.; Cerri, P.S.; Cells, B. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. BioMed Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef]
- Chan, W.C.W.; Tan, Z.; To, M.K.T.; Chan, D. Regulation and role of transcription factors in osteogenesis. Int. J. Mol. Sci. 2021, 22, 5445. [Google Scholar] [CrossRef]
- Mohamed, A.M.F.S. An overview of bone cells and their regulating factors of differentiation. Malaysian J. Med. Sci. 2008, 15, 4–12. [Google Scholar]
- Marie, P.J. Bone cell-matrix protein interactions. Osteoporos. Int. 2009, 20, 1037–1042. [Google Scholar] [CrossRef]
- Lin, X.; Patil, S.; Gao, Y.G.; Qian, A. The Bone Extracellular Matrix in Bone Formation and Regeneration. Front. Pharmacol. 2020, 11, 00757. [Google Scholar] [CrossRef]
- Chen, S.; He, T.; Zhong, Y.; Chen, M.; Yao, Q.; Chen, D.; Shao, Z.; Xiao, G. Roles of focal adhesion proteins in skeleton and diseases. Acta Pharm. Sin. B 2022, 13, 998–1013. [Google Scholar] [CrossRef]
- Boglione, C.; Gavaia, P.; Koumoundouros, G.; Gisbert, E.; Moren, M.; Witten, P.E. Skeletal anomalies in reared European fish larvae and juveniles. Part 1: Normal and anomalous skeletogenic processes. Rev. Aquac. 2013, 5, 99–120. [Google Scholar] [CrossRef]
- Loizides, M.; Georgiou, A.N.; Somarakis, S.; Witten, P.E.; Koumoundouros, G. A new type of lordosis and vertebral body compression in Gilthead sea bream, Sparus aurata L.: Aetiology, anatomy and consequences for survival. J. Fish Dis. 2014, 37, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Witten, P.E.; Gil-Martens, L.; Hall, B.K.; Huysseune, A.; Obach, A. Compressed vertebrae in Atlantic salmon Salmo salar: Evidence for metaplastic chondrogenesis as a skeletogenic response late in ontogeny. Dis. Aquat. Organ. 2005, 64, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Ytteborg, E.; Torgersen, J.; Baeverfjord, G.; Takle, H. Morphological and molecular characterization of developing vertebral fusions using a teleost model. BMC Physiol. 2010, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Ytteborg, E.; Torgersen, J.; Baeverfjord, G.; Takle, H. The Atlantic Salmon (Salmo salar) Vertebra and Cellular Pathways to Vertebral Deformities. Health Environ. Aquac. 2012, 2012, 329–358. [Google Scholar] [CrossRef]
- Witten, P.E.; Obach, A.; Huysseune, A.; Baeverfjord, G. Vertebrae fusion in Atlantic salmon (Salmo salar): Development, aggravation and pathways of containment. Aquaculture 2006, 258, 164–172. [Google Scholar] [CrossRef]
- Helland, S.; Denstadli, V.; Witten, P.E.; Hjelde, K.; Storebakken, T.; Skrede, A.; Åsgård, T.; Baeverfjord, G. Hyper dense vertebrae and mineral content in Atlantic salmon(Salmo salar L.) fed diets with graded levels of phytic acid. Aquaculture 2006, 261, 603–614. [Google Scholar] [CrossRef]
- Gil-Martens, L. Inflammation as a potential risk factor for spinal deformities in farmed Atlantic salmon (Salmo salar L.). J. Appl. Ichthyol. 2010, 26, 350–354. [Google Scholar] [CrossRef]
- Ortiz-Delgado, J.B.; Fernández, I.; Sarasquete, C.; Gisbert, E. Normal and histopathological organization of the opercular bone and vertebrae in gilthead sea bream Sparus aurata. Aquat. Biol. 2014, 21, 67–84. [Google Scholar] [CrossRef]
- Negrín-Báez, D.; Navarro, A.; Lee-Montero, I.; Soula, M.; Afonso, J.M.; Zamorano, M.J. Inheritance of skeletal deformities in gilthead seabream (Sparus aurata) –lack of operculum, lordosis, vertebral fusion and LSK complex. J. Anim. Sci. 2015, 93, 53–61. [Google Scholar] [CrossRef][Green Version]
- Han, M.; Luo, M.; Yang, R.; Qin, J.G.; Ma, Z. Impact of temperature on survival and spinal development of golden pompano Trachinotus ovatus (Linnaeus 1758 ). Aquac. Rep. 2020, 18, 100556. [Google Scholar] [CrossRef]
- Gavaia, P.J.; Domingues, S.; Engrola, S.; Drake, P.; Sarasquete, C.; Dinis, M.T.; Cancela, M.L. Comparing skeletal development of wild and hatchery-reared Senegalese sole (Solea senegalensis, Kaup 1858): Evaluation in larval and postlarval stages. Aquac. Res. 2009, 40, 1585–1593. [Google Scholar] [CrossRef]
- De Azevedo, A.M.; Losada, A.P.; Barreiro, A.; Vázquez, S.; Quiroga, M.I. Skeletal Anomalies in Senegalese Sole (Solea senegalensis), an Anosteocytic Boned Flatfish Species. Vet. Pathol. 2019, 56, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Berillis, P. Factors That Can Lead To the Development of Skeletal. J. Fish. Sci. 2015, 9, 17–23. [Google Scholar]
- Sawada, Y.; Hattori, M.; Sudo, N.; Kato, K.; Takagi, Y.; Ura, K.; Kurata, M.; Okada, T.; Kumai, H. Hypoxic conditions induce centrum defects in red sea bream Pagrus major (Temminck and Schlegel). Aquac. Res. 2006, 37, 805–812. [Google Scholar] [CrossRef]
- Lall, S.P.; Lewis-McCrea, L.M. Role of nutrients in skeletal metabolism and pathology in fish—An overview. Aquaculture 2007, 267, 3–19. [Google Scholar] [CrossRef]
- Cobcroft, J.M.; Shu-Chien, A.C.; Kuah, M.K.; Jaya-Ram, A.; Battaglene, S.C. The effects of tank colour, live food enrichment and greenwater on the early onset of jaw malformation in striped trumpeter larvae. Aquaculture 2012, 356–357, 61–72. [Google Scholar] [CrossRef]
- Boglino, A.; Darias, M.J.; Estévez, A.; Andree, K.B.; Gisbert, E. The effect of dietary arachidonic acid during the Artemia feeding period on larval growth and skeletogenesis in Senegalese sole, Solea senegalensis. J. Appl. Ichthyol. 2012, 28, 411–418. [Google Scholar] [CrossRef]
- Liu, D.; Veit, H.P.; Denbow, D.M. Effects of long-term dietary lipids on mature bone mineral content, collagen, crosslinks, and prostaglandin E2 production in Japanese quail. Poult. Sci. 2004, 83, 1876–1883. [Google Scholar] [CrossRef]
- Cahu, C.; Infante, J.Z.; Takeuchi, T. Nutritional components affecting skeletal development in fish larvae. Aquaculture 2003, 227, 245–258. [Google Scholar] [CrossRef]
- Lau, B.Y.Y.; Cohen, D.J.A.; Ward, W.E.; Ma, D.W.L. Investigating the role of polyunsaturated fatty acids in bone development using animal models. Molecules 2013, 18, 14203–14227. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Zhang, K.; Wei, Y.; Hua, W.; Gao, Y.; Li, X.; Ye, L. Therapeutic potentials and modulatory mechanisms of fatty acids in bone. Cell Prolif. 2020, 53, e12735. [Google Scholar] [CrossRef] [PubMed]
- Roo, J.; Izquierdo, M.S. Effect of increasing n-3 HUFA content in enriched Artemia on growth, survival and skeleton anomalies occurrence of greater amberjack Seriola dumerili larvae. Aquaculture 2019, 500, 651–659. [Google Scholar] [CrossRef]
- Lean, J.M.; Jagger, C.J.; Kirstein, B.; Fuller, K.; Chambers, T.J. Hydrogen peroxide is essential for estrogen-deficiency bone loss and osteoclast formation. Endocrinology 2005, 146, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Wauquier, F.; Leotoing, L. Oxidative stress in bone remodelling and disease. Trends Mol. Med. 2009, 15, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Gil Martens, L.; Lock, E.J.; Fjelldal, P.G.; Wargelius, A.; Araujo, P.; Torstensen, B.E.; Witten, P.E.; Hansen, T.; Waagbø, R.; Ørnsrud, R. Dietary fatty acids and inflammation in the vertebral column of Atlantic salmon, Salmo salar L., smolts: A possible link to spinal deformities. J. Fish Dis. 2010, 33, 957–972. [Google Scholar] [CrossRef] [PubMed]
- Gavaia, P.J.; Cancela, M.L.; Laize, V. Fish: A suitable system to model human bone disorders and discover drugs with osteogenic or osteotoxic activities. Drug Discov. Today Dis. Models 2014, 13, 29–37. [Google Scholar] [CrossRef]
- Harris, M.P.; Henke, K.; Hawkins, M.B.; Witten, P.E. Fish is Fish: The use of experimental model species to reveal causes of skeletal diversity in evolution and disease. J. Appl. Ichthyol. 2014, 30, 616–629. [Google Scholar] [CrossRef]
- Casado-Díaz, A.; Santiago-Mora, R.; Dorado, G.; Quesada-Gómez, J.M. The omega-6 arachidonic fatty acid, but not the omega-3 fatty acids, inhibits osteoblastogenesis and induces adipogenesis of human mesenchymal stem cells: Potential implication in osteoporosis. Osteoporos. Int. 2013, 24, 1647–1661. [Google Scholar] [CrossRef]
- Boeyens, J.C.A.; Deepak, V.; Chua, W.H.; Kruger, M.C.; Joubert, A.M.; Coetzee, M. Effects of ω3- and ω6-polyunsaturated fatty acids on RANKL-induced osteoclast differentiation of RAW264.7 cells: A comparative in vitro study. Nutrients 2014, 6, 2584–2601. [Google Scholar] [CrossRef] [PubMed]
- Cornish, J.; Macgibbon, A.; Lin, J.; Watson, M.; Callon, K.E.; Tong, P.C.; Dunford, J.E.; Van Der Does, Y.; Williams, G.A.; Grey, A.B.; et al. Modulation of Osteoclastogenesis by Fatty Acids. Endocrinology 2008, 149, 5688–5695. [Google Scholar] [CrossRef] [PubMed]
- Musacchio, E.; Priante, G.; Budakovic, A.; Baggio, B. Effects of unsaturated free fatty acids on adhesion and on gene expression of extracellular matrix macromolecules in human Osteoblast-like cell cultures. Connect. Tissue Res. 2007, 48, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Ambrogini, E.; Han, L.; Manolagas, S.C.; Jilka, R.L. Increased lipid oxidation causes oxidative stress, increased peroxisome proliferator-activated receptor-γ expression, and diminished pro-osteogenic Wnt signaling in the skeleton. J. Biol. Chem. 2009, 284, 27438–27448. [Google Scholar] [CrossRef] [PubMed]
- Claassen, N.; Coetzer, H.; Steinmann, C.M.L.; Kruger, M.C. The effect of different n-6 n-3 essential fatty acid ratios on calcium balance and bone in rats. Prostaglandins Leukot. Essent. Fat. Acids 1995, 53, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Kruger, M.C.; Coetzer, H.; De Winter, R.; Gericke, G.; Van Papendorp, D.H. Calcium, gamma-linolenic acid and eicosapentaenoic acid supplementation in senile osteoporosis. Aging Clin. Exp. Res. 1998, 10, 385–394. [Google Scholar] [CrossRef]
- Wann, A.K.T.; Mistry, J.; Blain, E.J.; Michael-Titus, A.T.; Knight, M.M. Eicosapentaenoic acid and docosahexaenoic acid reduce interleukin-1β-mediated cartilage degradation. Arthritis Res. Ther. 2010, 12, R207. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef]
- De Crombrugghe, B.; Lefebvre, V.; Behringer, R.R.; Bi, W.; Murakami, S.; Huang, W. Transcriptional mechanisms of chondrocyte differentiation. Matrix Biol. 2000, 19, 389–394. [Google Scholar] [CrossRef]
- Sims, N.A.; Martin, T.J. Coupling signals between the osteoclast and osteoblast: How are messages transmitted between these temporary visitors to the bone surface? Front. Endocrinol. 2015, 6, 00041. [Google Scholar] [CrossRef]
- Negishi-Koga, T.; Takayanagi, H. Bone cell communication factors and Semaphorins. Bonekey Rep. 2012, 1, 183. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef] [PubMed]
- Akune, T.; Ohba, S.; Kamekura, S.; Yamaguchi, M.; Chung, U.; Kubota, N.; Terauchi, Y.; Harada, Y.; Azuma, Y.; Nakamura, K.; et al. PPAR γ insufficiency enhances osteogenesis through osteoblast formation from bone marrow progenitors. J. Clin. Investig. 2004, 113, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Giovannone, D.; Paul, S.; Schindler, S.; Arata, C.; Farmer, D.T.; Patel, P.; Smeeton, J.; Gage Crump, J. Programmed conversion of hypertrophic chondrocytes into osteoblasts and marrow adipocytes within zebrafish bones. Elife 2019, 8, e42736. [Google Scholar] [CrossRef]
- Takigawa, Y.; Hata, K.; Muramatsu, S.; Amano, K.; Ono, K.; Wakabayashi, M.; Matsuda, A.; Takada, K.; Nishimura, R.; Yoneda, T. The transcription factor Znf219 regulates chondrocyte differentiation by assembling a transcription factory with Sox9. J. Cell Sci. 2010, 123, 3780–3788. [Google Scholar] [CrossRef]
- Paudel, S.; Gjorcheska, S.; Bump, P.; Barske, L. Patterning of cartilaginous condensations in the developing facial skeleton. Dev. Biol. 2022, 486, 44–55. [Google Scholar] [CrossRef]
- Sperber, S.M.; Dawid, I.B. Barx1 Is Necessary for Ectomesenchyme Proliferation and Osteochondroprogenitor Condensation in the Zebrafish Pharyngeal Arches. Dev. Biol. 2008, 321, 101–110. [Google Scholar] [CrossRef]
- Ionescu, A.; Kozhemyakina, E.; Nicolae, C.; Kaestner, K.H.; Olsen, B.R.; Lassar, A.B. FoxA Family Members Are Crucial Regulators of the Hypertrophic Chondrocyte Differentiation Program. Dev. Cell 2012, 22, 927–939. [Google Scholar] [CrossRef]
- Bell, N.; Bhagat, S.; Muruganandan, S.; Kim, R.; Ho, K.; Pierce, R.; Kozhemyakina, E.; Lassar, A.B.; Gamer, L.; Rosen, V.; et al. Overexpression of transcription factor FoxA2 in the developing skeleton causes an enlargement of the cartilage hypertrophic zone, but it does not trigger ectopic differentiation in immature chondrocytes. Bone 2022, 160, 116418. [Google Scholar] [CrossRef]
- Arnold, M.A.; Kim, Y.; Czubryt, M.P.; Phan, D.; McAnally, J.; Qi, X.; Shelton, J.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. MEF2C Transcription Factor Controls Chondrocyte Hypertrophy and Bone Development. Dev. Cell 2007, 12, 377–389. [Google Scholar] [CrossRef]
- Blixt, N.; Norton, A.; Zhang, A.; Aparicio, C.; Prasad, H.; Gopalakrishnan, R.; Jensen, E.D.; Mansky, K.C. Loss of myocyte enhancer factor 2 expression in osteoclasts leads to opposing skeletal phenotypes. Bone 2020, 138, 115466. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Murata, K.; Mun, S.H.; Bae, S.; Lee, Y.J.; Pannellini, T.; Kang, K.; Oliver, D.; Park-Min, K.H.; Ivashkiv, L.B. MEF2C regulates osteoclastogenesis and pathologic bone resorption via c-FOS. Bone Res. 2021, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Izawa, T.; Rohatgi, N.; Fukunaga, T.; Wang, Q.T.; Silva, M.J.; Gardner, M.J.; McDaniel, M.L.; Abumrad, N.A.; Semenkovich, C.F.; Teitelbaum, S.L.; et al. ASXL2 Regulates Glucose, Lipid, and Skeletal Homeostasis. Cell Rep. 2015, 11, 1625–1637. [Google Scholar] [CrossRef] [PubMed]
- Goos, J.A.C.; Vogel, W.K.; Mlcochova, H.; Millard, C.J.; Esfandiari, E.; Selman, W.H.; Calpena, E.; Koelling, N.; Carpenter, E.L.; Swagemakers, S.M.A.; et al. A de novo substitution in BCL11B leads to loss of interaction with transcriptional complexes and craniosynostosis. Hum. Mol. Genet. 2019, 28, 2501–2513. [Google Scholar] [CrossRef]
- Holmes, G.; Van Bakel, H.; Zhou, X.; Losic, B.; Jabs, E.W. Gene Expression Patterns BCL11B expression in intramembranous osteogenesis during murine craniofacial suture development. Gene Expr. Patterns 2015, 17, 16–25. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, W.; Yang, S.; Shao, J.; Li, Y.P. Signaling and transcriptional regulation in osteoblast commitment and differentiation. Front Biosci. 2007, 12, 3068–3092. [Google Scholar] [CrossRef]
- Smith, S.Y. Bone physiology and biology. In Bone Toxicology; Springer: Berlin/Heidelberg, Germany, 2018; Chapter 2; ISBN 9783319561929. [Google Scholar]
- Chen, G.; Deng, C.; Li, Y.P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef]
- Chau, J.F.L.; Leong, W.F.; Li, B. Signaling pathways governing osteoblast proliferation, differentiation and function. Histol. Histopathol. 2009, 24, 1593–1606. [Google Scholar] [CrossRef]
- Wu, M.; Chen, G.; Li, Y.P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Sanford, L.P.; Ormsby, I.; Gittenberger-de Groot, A.C.; Sariola, H.; Friedman, R.; Boivin, G.P.; Cardell, E.L.; Doetschman, T. TGFβ2 knockout mice have multiple developmental defects that are non-overlapping with other TGFβ knockout phenotypes. Development 1997, 124, 2659–2670. [Google Scholar] [CrossRef]
- Spencer, G.J.; Utting, J.C.; Etheridge, S.L.; Arnett, T.R.; Genever, P.G. Wnt signalling in osteoblasts regulates expression of the receptor activator of NFκB ligand and inhibits osteoclastogenesis in vitro. J. Cell Sci. 2006, 119, 1283–1296. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Chen, M.; Zuscik, M.; Wu, Q.; Wang, Y.J.; Rosier, R.N.; O’Keefe, R.J.; Chen, D. Inhibition of β-catenin signaling in articular chondrocytes results in articular cartilage destruction. Arthritis Rheum. 2008, 58, 2053–2064. [Google Scholar] [CrossRef] [PubMed]
- Riddle, R.C.; Diegel, C.R.; Leslie, J.M.; van Koevering, K.K.; Faugere, M.C.; Clemens, T.L.; Williams, B.O. Lrp5 and Lrp6 Exert Overlapping Functions in Osteoblasts during Postnatal Bone Acquisition. PLoS ONE 2013, 8, e63323. [Google Scholar] [CrossRef] [PubMed]
- Joeng, K.S.; Schumacher, C.A.; Zylstra-Diegel, C.R.; Long, F.; Williams, B.O. Lrp5 and Lrp6 redundantly control skeletal development in the mouse embryo. Dev. Biol. 2011, 359, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Repudi, S.R.; Patra, M.; Sen, M. WISP3-IGF1 interaction regulates chondrocyte hypertrophy. J. Cell Sci. 2013, 126, 1650–1658. [Google Scholar] [CrossRef] [PubMed]
- Engin, F.; Yao, Z.; Yang, T.; Zhou, G.; Bertin, T.; Jiang, M.M.; Chen, Y.; Wang, L.; Zheng, H.; Sutton, R.E.; et al. Dimorphic effects of Notch signaling in bone homeostasis. Nat. Med. 2008, 14, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Canalis, E. Notch and the regulation of osteoclast differentiation and function. Bone 2020, 138, 115474. [Google Scholar] [CrossRef]
- Mak, K.K.; Chen, M.H.; Day, T.F.; Chuang, P.T.; Yang, Y. Wnt/β-catenin signaling interacts differentially with Ihh signaling in controlling endochondral bone and synovial joint formation. Development 2006, 133, 3695–3707. [Google Scholar] [CrossRef]
- Mao, L.; Wang, L.; Xu, J.; Zou, J. The role of integrin family in bone metabolism and tumor bone metastasis. Cell Death Discov. 2023, 9, 119. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, L.; Wang, N.; Li, J.; He, F.; Li, X.; Wu, S. Unexpected role of matrix Gla protein in osteoclasts: Inhibiting osteoclast differentiation and bone resorption. Mol. Cell. Biol. 2019, 39, e00012-19. [Google Scholar] [CrossRef]
- Yagami, K.; Suh, J.Y.; Enomoto-Iwamoto, M.; Koyama, E.; Abrams, W.R.; Shapiro, I.M.; Pacifici, M.; Iwamoto, M. Matrix GLA protein is a developmental regulator of chondrocyte mineralization and, when constitutively expressed, blocks endochondral and intramembranous ossification in the limb. J. Cell Biol. 1999, 147, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Kwan, K.M.; Zhang, Z.; Deng, J.M.; Darnay, B.G.; Behringer, R.R.; Nakamura, T.; de Crombrugghe, B.; Akiyama, H. Cthrci is a positive regulator of osteoblastic bone formation. PLoS ONE 2008, 3, e3174. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, K.; Kohara, Y.; Naoe, Y.; Watanabe, A.; Ito, M.; Ikeda, K.; Takeshita, S. WAIF1 Is a Cell-Surface CTHRC1 Binding Protein Coupling Bone Resorption and Formation. J. Bone Miner. Res. 2018, 33, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Imuta, Y.; Nishioka, N.; Kiyonari, H.; Sasaki, H. Short limbs, cleft palate, and delayed formation of flat proliferative chondrocytes in mice with targeted disruption of a putative protein kinase gene, Pkdcc (AW548124). Dev. Dyn. 2009, 238, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Gersch, R.P.; Hadjiargyrou, M. Mustn1 is expressed during chondrogenesis and is necessary for chondrocyte proliferation and differentiation in vitro. Bone 2009, 45, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Linares, G.R.; Brommage, R.; Powell, D.R.; Xing, W.; Chen, S.T.; Alshbool, F.Z.; Lau, K.H.W.; Wergedal, J.E.; Mohan, S. Claudin 18 is a novel negative regulator of bone resorption and osteoclast differentiation. J. Bone Miner. Res. 2012, 27, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, J.; Zhang, F.P.; Surmann-Schmitt, C.; Honkala, S.; Stock, M.; Poutanen, M.; Säämänen, A.M. Defects in chondrocyte maturation and secondary ossification in mouse knee joint epiphyses due to Snorc deficiency. Osteoarthr. Cartil. 2017, 25, 1132–1142. [Google Scholar] [CrossRef]
- Shimazu, A.; Nah, H.D.; Kirsch, T.; Koyama, E.; Leatherman, J.L.; Golden, E.B.; Kosher, R.A.; Pacifici, M. Syndecan-3 and the control of chondrocyte proliferation during endochondral ossification. Exp. Cell Res. 1996, 229, 126–136. [Google Scholar] [CrossRef]
- Mansouri, R.; Jouan, Y.; Hay, E.; Blin-Wakkach, C.; Frain, M.; Ostertag, A.; Le Henaff, C.; Marty, C.; Geoffroy, V.; Marie, P.J.; et al. Osteoblastic heparan sulfate glycosaminoglycans control bone remodeling by regulating Wnt signaling and the crosstalk between bone surface and marrow cells. Cell Death Dis. 2017, 8, e2902. [Google Scholar] [CrossRef]
- Kirsch, T.; Koyama, E.; Liu, M.; Golub, E.E.; Pacifici, M. Syndecan-3 is a selective regulator of chondrocyte proliferation. J. Biol. Chem. 2002, 277, 42171–42177. [Google Scholar] [CrossRef]
- Teplyuk, N.M.; Haupt, L.M.; Ling, L.; Dombrowski, C.; Kin, F.; Nathan, S.S.; Lian, J.B.; Stein, J.L.; Stein, G.S.; Simon, M.; et al. The osteogenic transcription factor Runx2 regulates components of the fibroblast growth factor/proteoglycan signaling axis in osteoblasts. J. Cell. Biochem. 2010, 107, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hao, J.; Duan, X.; Wu, N.; Zhou, Z.; Yang, F.; Li, J.; Zhao, Z.; Huang, S. The role of semaphorin 3A in bone remodeling. Front. Cell. Neurosci. 2017, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Xing, Q.; Feng, J.; Zhang, X. Semaphorin3B Promotes Proliferation and Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells in a High-Glucose Microenvironment. Stem Cells Int. 2021, 2021, 6637176. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.; Burt-Pichat, B.; Mallein-Gerin, F.; Merle, B.; Delmas, P.D.; Skerry, T.M.; Vico, L.; Malaval, L.; Chenu, C. Expression of Semaphorin-3A and its receptors in endochondral ossification: Potential role in skeletal development and innervation. Dev. Dyn. 2005, 234, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Mizuhashi, K.; Kanamoto, T.; Moriishi, T.; Muranishi, Y.; Miyazaki, T.; Terada, K.; Omori, Y.; Ito, M.; Komori, T.; Furukawa, T. Filamin-interacting proteins, Cfm1 and Cfm2, are essential for the formation of cartilaginous skeletal elements. Hum. Mol. Genet. 2014, 23, 2953–2967. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Park, H.R.; Lee, E.J.; Jang, J.A.; Han, M.S.; Kim, G.W.; Jeong, J.H.; Choi, J.Y.; Beier, F.; Jung, Y.K. Dicam promotes proliferation and maturation of chondrocyte through Indian hedgehog signaling in primary cilia. Osteoarthr. Cartil. 2018, 26, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J. Osteoblast dysfunctions in bone diseases: From cellular and molecular mechanisms to therapeutic strategies. Cell. Mol. Life Sci. 2015, 72, 1347–1361. [Google Scholar] [CrossRef]
- Sroga, G.E.; Vashishth, D. Effects of bone matrix proteins on fracture and fragility in osteoporosis. Curr. Osteoporos. Rep. 2012, 10, 141–150. [Google Scholar] [CrossRef]
- Cheah, K.S.E.; Lau, E.T.; Au, P.K.C.; Tam, P.P.L. Expression of the mouse α1(II) collagen gene is not restricted to cartilage during development. Development 1991, 111, 945–953. [Google Scholar] [CrossRef]
- Lavrijsen, I.C.M.; Leegwater, P.A.J.; Martin, A.J.; Harris, S.J.; Tryfonidou, M.A.; Heuven, H.C.M.; Hazewinkel, H.A.W. Genome wide analysis indicates genes for basement membrane and cartilage matrix proteins as candidates for hip dysplasia in labrador retrievers. PLoS ONE 2014, 9, e87735. [Google Scholar] [CrossRef]
- Koivunen, J.; Kemppainen, A.V.; Finnilä, M.A.; Keski-Filppula, R.; Härönen, H.; Tu, H.; Pellikka, H.; Heikkinen, A.; Kylmäoja, E.; Sormunen, R.; et al. Collagen XIII-derived ectodomain regulates bone angiogenesis and intracortical remodeling. Matrix Biol. 2019, 83, 6–25. [Google Scholar] [CrossRef] [PubMed]
- Yun-Feng, W.; Matsuo, N.; Sumiyoshi, H.; Yoshioka, H. Sp7/Osterix up-regulates the mouse pro-α3(V) collagen gene (Col5a3) during the osteoblast differentiation. Biochem. Biophys. Res. Commun. 2010, 394, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Foldager, C.B.; Toh, W.S.; Christensen, B.B.; Lind, M.; Gomoll, A.H.; Spector, M. Collagen Type IV and Laminin Expressions during Cartilage Repair and in Late Clinically Failed Repair Tissues from Human Subjects. Cartilage 2016, 7, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Heilig, J.; Dietmar, H.F.; Brachvogel, B.; Paulsson, M.; Zaucke, F.; Niehoff, A. Collagen IX deficiency leads to premature vascularization and ossification of murine femoral heads through an imbalance of pro- and antiangiogenic factors. Osteoarthr. Cartil. 2020, 28, 988–999. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, B.C.; Belluoccio, D.; Rowley, L.; Little, C.B.; Hansen, U.; Bateman, J.F. Cartilage intermediate layer protein 2 (CILP-2) is expressed in articular and meniscal cartilage and down-regulated in experimental osteoarthritis. J. Biol. Chem. 2011, 286, 37758–37767. [Google Scholar] [CrossRef]
- Horner, A.; Bord, S.; Kelsall, A.W.; Coleman, N.; Compston, J.E. Tie2 ligands angiopoietin-1 and angiopoietin-2 are coexpressed with vascular endothelial cell growth factor in growing human bone. Bone 2001, 28, 65–71. [Google Scholar] [CrossRef]
- Xie, A.; Xue, J.; Shen, G.; Nie, L. Thrombospondin-1 inhibits ossification of tissue engineered cartilage constructed by ADSCs. Am. J. Transl. Res. 2017, 9, 3487–3498. [Google Scholar]
- Simantov, R.; Silverstein, R.L. CD36: A Critical Anti-Angiogenic Receptor. J. Biol. Chem. 2003, 8, s874–s882. [Google Scholar] [CrossRef]
- Galeano-Garces, C.; Camilleri, E.T.; Riester, S.M.; Dudakovic, A.; Larson, D.R.; Qu, W.; Smith, J.; Dietz, A.B.; Im, H.J.; Krych, A.J.; et al. Molecular Validation of Chondrogenic Differentiation and Hypoxia Responsiveness of Platelet-Lysate Expanded Adipose Tissue–Derived Human Mesenchymal Stromal Cells. Cartilage 2017, 8, 283–299. [Google Scholar] [CrossRef]
- Coulson-Thomas, Y.M.; Coulson-Thomas, V.J.; Norton, A.L.; Gesteira, T.F.; Cavalheiro, R.P.; Meneghetti, M.C.Z.; Martins, J.R.; Dixon, R.A.; Nader, H.B. The identification of proteoglycans and glycosaminoglycans in archaeological human bones and teeth. PLoS ONE 2015, 10, 0131105. [Google Scholar] [CrossRef]
- Han, J.; Li, D.; Qu, C.; Wang, D.; Wang, L.; Guo, X.; Lammi, M.J. Altered expression of chondroitin sulfate structure modifying sulfotransferases in the articular cartilage from adult osteoarthritis and Kashin-Beck disease. Osteoarthr. Cartil. 2017, 25, 1372–1375. [Google Scholar] [CrossRef] [PubMed]
- Nakamichi, Y.; Shukunami, C.; Yamada, T.; Aihara, K.; Kawano, H.; Sato, T.; Nishizaki, Y.; Yamamoto, Y.; Shindo, M.; Yoshimura, K.; et al. Chondromodulin I Is a Bone Remodeling Factor. Mol. Cell. Biol. 2003, 23, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Norris, E.L.; Brachvogel, B.; Angelucci, C.; Zivkovic, S.; Gordon, L.; Bernardo, B.C.; Stermann, J.; Sekiguchi, K.; Gorman, J.J.; et al. Changes in the chondrocyte and extracellular matrix proteome during post-natal mouse cartilage development. Mol. Cell. Proteom. 2012, 11, M111.014159. [Google Scholar] [CrossRef]
- Nicolae, C.; Ko, Y.P.; Miosge, N.; Niehoff, A.; Studer, D.; Enggist, L.; Hunziker, E.B.; Paulsson, M.; Wagener, R.; Aszodi, A. Abnormal collagen fibrils in cartilage of matrilin-1/matrilin-3-deficient mice. J. Biol. Chem. 2007, 282, 22163–22175. [Google Scholar] [CrossRef] [PubMed]
- Siller, A.F.; Whyte, M.P. Alkaline Phosphatase: Discovery and Naming of Our Favorite Enzyme. J. Bone Miner. Res. 2018, 33, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Inada, M.; Wang, Y.; Byrne, M.H.; Rahman, M.U.; Miyaura, C.; López-Otín, C.; Krane, S.M. Critical roles for collagenase-3 (Mmp13) in development of growth plate cartilage and in endochondral ossification. Proc. Natl. Acad. Sci. USA 2004, 101, 17192–17197. [Google Scholar] [CrossRef] [PubMed]
- Herroon, M.K.; Rajagurubandara, E.; Hardaway, A.L.; Powell, K.; Turchick, A.; Feldmann, D.; Podgorski, I. Bone marrow adipocytes promote tumor growth in bone via FABP4-dependent mechanisms. Oncotarget 2013, 4, 2108–2123. [Google Scholar] [CrossRef]
- Kevorkova, O.; Martineau, C.; Martin-Falstrault, L.; Sanchez-Dardon, J.; Brissette, L.; Moreau, R. Low-Bone-Mass Phenotype of Deficient Mice for the Cluster of Differentiation 36 (CD36). PLoS ONE 2013, 8, 0077701. [Google Scholar] [CrossRef]
- Koduru, S.V.; Sun, B.H.; Walker, J.M.; Zhu, M.; Simpson, C.; Dhodapkar, M.; Insogna, K.L. The contribution of cross-talk between the cell-surface proteins CD36 and CD47-TSP-1 in osteoclast formation and function. J. Biol. Chem. 2018, 293, 15055–15069. [Google Scholar] [CrossRef]
- Weiss, R.E.; Reddi, A.H.; Nimni, M.E. Somatostatin can locally inhibit proliferation and differentiation of cartilage and bone precursor cells. Calcif. Tissue Int. 1981, 33, 425–430. [Google Scholar] [CrossRef]
- Genetos, D.C.; Wong, A.; Weber, T.J.; Karin, N.J.; Yellowley, C.E. Impaired osteoblast differentiation in annexin A2- And -A5-deficient cells. PLoS ONE 2014, 9, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Mori, Y.; Mori, D.; Chung, U.I.; Tanaka, S.; Heierhorst, J.; Buchou, T.; Baudier, J.; Kawaguchi, H.; Saito, T. S100A1 and S100B are dispensable for endochondral ossification during skeletal development. Biomed. Res. 2014, 35, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Nalesso, G.; Thorup, A.S.; Eldridge, S.E.; De Palma, A.; Kaur, A.; Peddireddi, K.; Blighe, K.; Rana, S.; Stott, B.; Vincent, T.L.; et al. Calcium calmodulin kinase II activity is required for cartilage homeostasis in osteoarthritis. Sci. Rep. 2021, 11, 5682. [Google Scholar] [CrossRef] [PubMed]
- Taschner, M.J.; Rafigh, M.; Lampert, F.; Schnaiter, S.; Hartmann, C. Ca2+/Calmodulin-dependent kinase II signaling causes skeletal overgrowth and premature chondrocyte maturation. Dev. Biol. 2008, 317, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.; Che, X.; Ang, M.J.; Cheon, S.; Lee, J.; Kim, K.S.; Lee, C.H.; Lee, S.Y.; Yang, H.Y.; Moon, C.; et al. Peroxiredoxin 5 regulates osteogenic differentiation through interaction with hnRNPK during bone regeneration. Elife 2023, 12, e80122. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Kim, D.Y.; Lee, D.S.; Kim, J.W.; Koh, J.T.; Kim, E.J.; Jang, W.G. Peroxiredoxin II negatively regulates BMP2-induced osteoblast differentiation and bone formation via PP2A Cα-mediated Smad1/5/9 dephosphorylation. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Board, P.G.; Coggan, M.; Chelvanayagam, G.; Easteal, S.; Jermiin, L.S.; Schulte, G.K.; Danley, D.E.; Hoth, L.R.; Griffor, M.C.; Kamath, A.V.; et al. Identification, characterization, and crystal structure of the omega class glutathione transferases. J. Biol. Chem. 2000, 275, 24798–24806. [Google Scholar] [CrossRef]
- Kazek, M.; Kaczmarek, A.; Wrońska, A.K.; Boguś, M.I. Conidiobolus coronatus induces oxidative stress and autophagy response in Galleria mellonella larvae. PLoS ONE 2020, 15, e0228407. [Google Scholar] [CrossRef]
- Chung, K.W.; Lee, E.K.; Lee, M.K.; Oh, G.T.; Yu, B.P.; Chung, H.Y. Impairment of PPARα and the fatty acid oxidation pathway aggravates renal fibrosis during aging. J Am Soc Nephrol. 2018, 29, 1223–1237. [Google Scholar] [CrossRef]
- Pégorier, J.P.; Le May, C.; Girard, J. Control of gene expression by fatty acids. J. Nutr. 2004, 134, 2444–2449. [Google Scholar] [CrossRef]
- Luján-Amoraga, L.; Marques, C.L.; Boglione, C.; Gavaia, P.J.; Castanho, S.; Bandarra, N.; Pousão-Ferreira, P.; Ribeiro, L. Dietary Omega-3 and Vitamin E on Skeletal Development, Bone Mineralisation and Their Relation with Oxidative Stress on Meagre, Argyrosomus regius Larvae; Aquaculture Research Station EPPO (IPMA): Olhão, Portugal, 2024; (to be submitted). [Google Scholar]
- Koren, N.; Simsa-maziel, S.; Shahar, R.; Schwartz, B.; Monsonego-ornan, E. ScienceDirect Exposure to omega-3 fatty acids at early age accelerate bone growth and improve bone quality. J. Nutr. Biochem. 2014, 25, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.; Betancor, M.B.; Roo, J.; Hernandez-Cruz, C.M.; Moyano, F.J.; Izquierdo, M. Optimum soybean lecithin contents in microdiets for gilthead seabream (Sparus aurata) larvae. Aquac. Nutr. 2013, 19, 585–597. [Google Scholar] [CrossRef]
- Boglino, A.; José, M.; Ortiz-delgado, J.B.; Özcan, F.; Estévez, A.; Andree, K.B.; Hontoria, F.; Sarasquete, C.; Gisbert, E. Commercial products for Artemia enrichment affect growth performance, digestive system maturation, ossi fi cation and incidence of skeletal deformities in Senegalese sole (Solea senegalensis) larvae. Aquaculture 2012, 324, 290–302. [Google Scholar] [CrossRef]
- Villeneuve, L.; Gisbert, E.; Zambonino-Infante, J.L.; Quazuguel, P.; Cahu, C.L. Effect of nature of dietary lipids on European sea bass morphogenesis: Implication of retinoid receptors. Br. J. Nutr. 2005, 94, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Mesa-rodriguez, A.; Betancor, B.; Hern, C.M. Effect of increasing docosahexaenoic acid content in weaning diets on survival, growth and skeletal anomalies of longfin yellowtail (Seriola rivoliana, Valenciennes 1833 ). Aquac. Res. 2018, 19, 1200–1209. [Google Scholar] [CrossRef]
- Betancor, M.B.; Atalah, E.; Caballero, M.; Benítez-Santana, T.; Roo, J.; Montero, D.; Izquierdo, M. α-Tocopherol in weaning diets for European sea bass (Dicentrarchus labrax) improves survival and reduces tissue damage caused by excess dietary DHA contents. Aquac. Nutr. 2011, 17, e112–e122. [Google Scholar] [CrossRef]
- Simões, T.; Fonseca, S.B.; Augusto, A.; Granada, L.; Ozório, R.O.A.; Gonçalves, J.F.M.; Pascoal, L.A.F.; Silva, J.H.V.; Lemos, M.F.L. Changes in fatty acid profile and chemical composition of meagre (Argyrosomus regius) fed with different lipid and selenium levels. Eur. J. Lipid Sci. Technol. 2017, 119, 1600016. [Google Scholar] [CrossRef]
- Raisz, L.G.; Pilbeam, C.C.; Fall, P.M. Prostaglandins: Mechanisms of action and regulation of production in bone. Osteoporos. Int. 1993, 3, 136–140. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Pair | Primer Sequence (5′-3′) | Length (bp) | GenBank Reference or Publication |
|---|---|---|---|---|
| sp7 | Sp7_fwd | TTCCTTTTGCGGCTTCAGAG | 142 bp | GFVG01056384.1 |
| Sp7_rev | GCCTGCACACACACATACAA | |||
| mgp | mgp_fwd | GCTGGCATTGAATCCCACAT | 171 bp | GFVG01044233.1 |
| mgp_rev | TGTTTCGGTCACCATCCACT | |||
| bmp2 | bmp2_fwd | TGTGGAATTTATCGGAGCCCA | 113 bp | GFVG01014899.1 |
| bmp2 _rev | CGAGCAGCAGTACCATGAGA | |||
| ef1a | ef1a _ fwd | GGTGCTGGACAAACTGAAGG | 161 bp | Ruiz et al. (2019) [42] |
| ef1a _ rev | GAACTCACCAACACCAGCAG | |||
| tub | tub _ fwd | GGAGTACCCCGATCGTATCA | 196 bp | Ruiz et al. (2019) [42] |
| tub _ rev | AGATGTCATACAGGGCCTCG |
| Reference Diet | Medium Diet | High Diet | |||||||
|---|---|---|---|---|---|---|---|---|---|
| R1 | R2 | R3 | M1 | M2 | M3 | H1 | H2 | H3 | |
| Total HQ reads | 180,488,089 | 164,478,326 | 163,786,858 | 152,078,855 | 179,103,633 | 253,267,203 | 164,384,967 | 150,515,239 | 163,786,580 |
| Mapped reads | 118,135,719 | 111,751,235 | 114,019,947 | 105,130,555 | 118,434,455 | 169,468,930 | 111,221,544 | 105,927,381 | 114,019,947 |
| Mapping rate (%) | 64.45% | 67.94% | 69.61% | 69.13% | 66.13% | 66.91% | 67.66% | 70.38% | 69.61% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luján-Amoraga, L.; Delgado-Martín, B.; Lourenço-Marques, C.; Gavaia, P.J.; Bravo, J.; Bandarra, N.M.; Dominguez, D.; Izquierdo, M.S.; Pousão-Ferreira, P.; Ribeiro, L. Exploring Omega-3′s Impact on the Expression of Bone-Related Genes in Meagre (Argyrosomus regius). Biomolecules 2024, 14, 56. https://doi.org/10.3390/biom14010056
Luján-Amoraga L, Delgado-Martín B, Lourenço-Marques C, Gavaia PJ, Bravo J, Bandarra NM, Dominguez D, Izquierdo MS, Pousão-Ferreira P, Ribeiro L. Exploring Omega-3′s Impact on the Expression of Bone-Related Genes in Meagre (Argyrosomus regius). Biomolecules. 2024; 14(1):56. https://doi.org/10.3390/biom14010056
Chicago/Turabian StyleLuján-Amoraga, Leticia, Belén Delgado-Martín, Cátia Lourenço-Marques, Paulo J. Gavaia, Jimena Bravo, Narcisa M. Bandarra, David Dominguez, Marisol S. Izquierdo, Pedro Pousão-Ferreira, and Laura Ribeiro. 2024. "Exploring Omega-3′s Impact on the Expression of Bone-Related Genes in Meagre (Argyrosomus regius)" Biomolecules 14, no. 1: 56. https://doi.org/10.3390/biom14010056
APA StyleLuján-Amoraga, L., Delgado-Martín, B., Lourenço-Marques, C., Gavaia, P. J., Bravo, J., Bandarra, N. M., Dominguez, D., Izquierdo, M. S., Pousão-Ferreira, P., & Ribeiro, L. (2024). Exploring Omega-3′s Impact on the Expression of Bone-Related Genes in Meagre (Argyrosomus regius). Biomolecules, 14(1), 56. https://doi.org/10.3390/biom14010056