
Functional, Morphological and Molecular Changes Reveal the Mechanisms Associated with Age-Related Vestibular Loss


 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Measurement of Vestibular Sensory-Evoked Potentials
2.3. Isolation and Immunofluorescent Staining of the Vestibular Sensory Epithelia
2.4. Confocal Microscopy and Quantitative Image Analysis of the Vestibular Sensory Epithelia
2.5. Vestibule Isolation, RNA Extraction, and RNA Sequencing
2.6. Differential Expression Analysis
2.7. Gene Ontology Enrichment Analysis
2.8. Statistical Analyses
3. Results
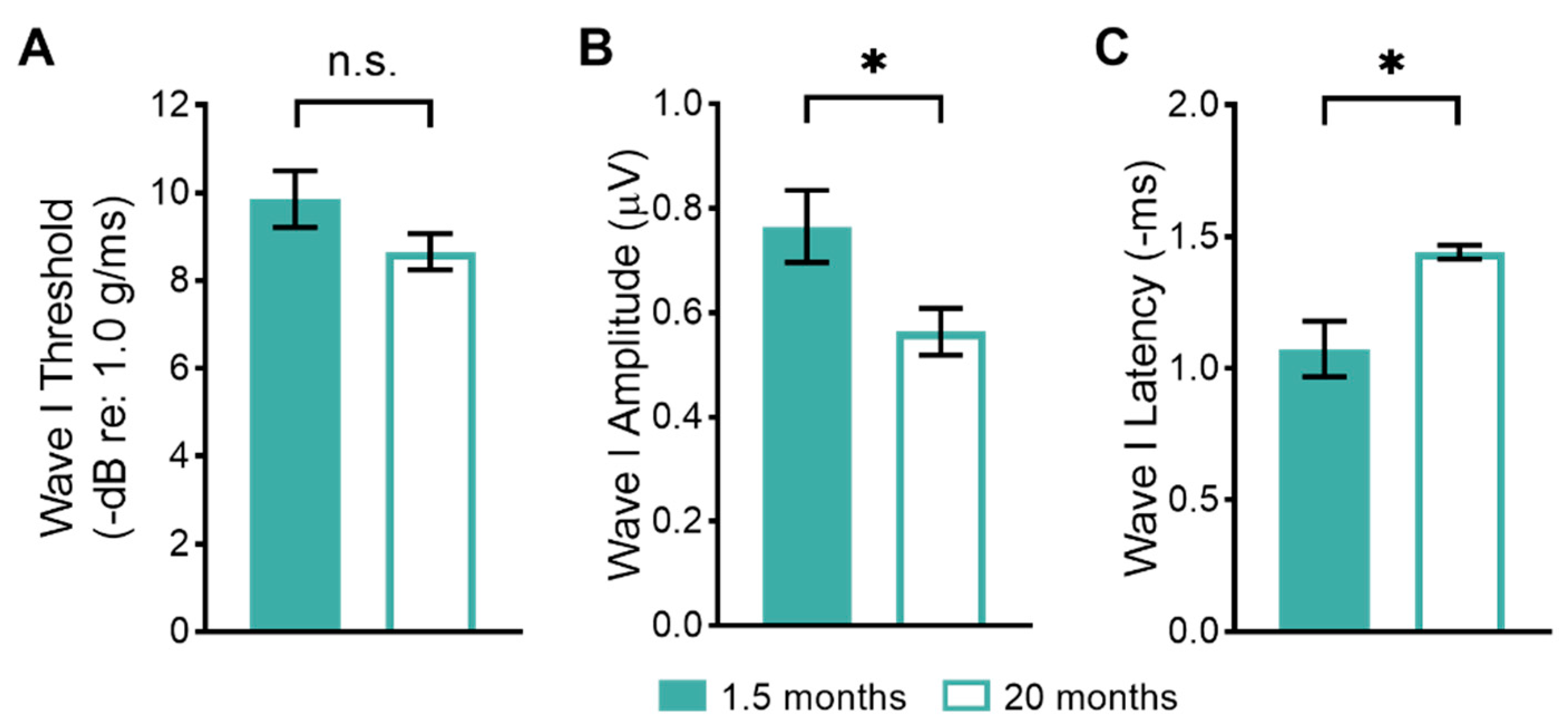
3.1. Age-Related Changes in Peripheral Vestibular Function
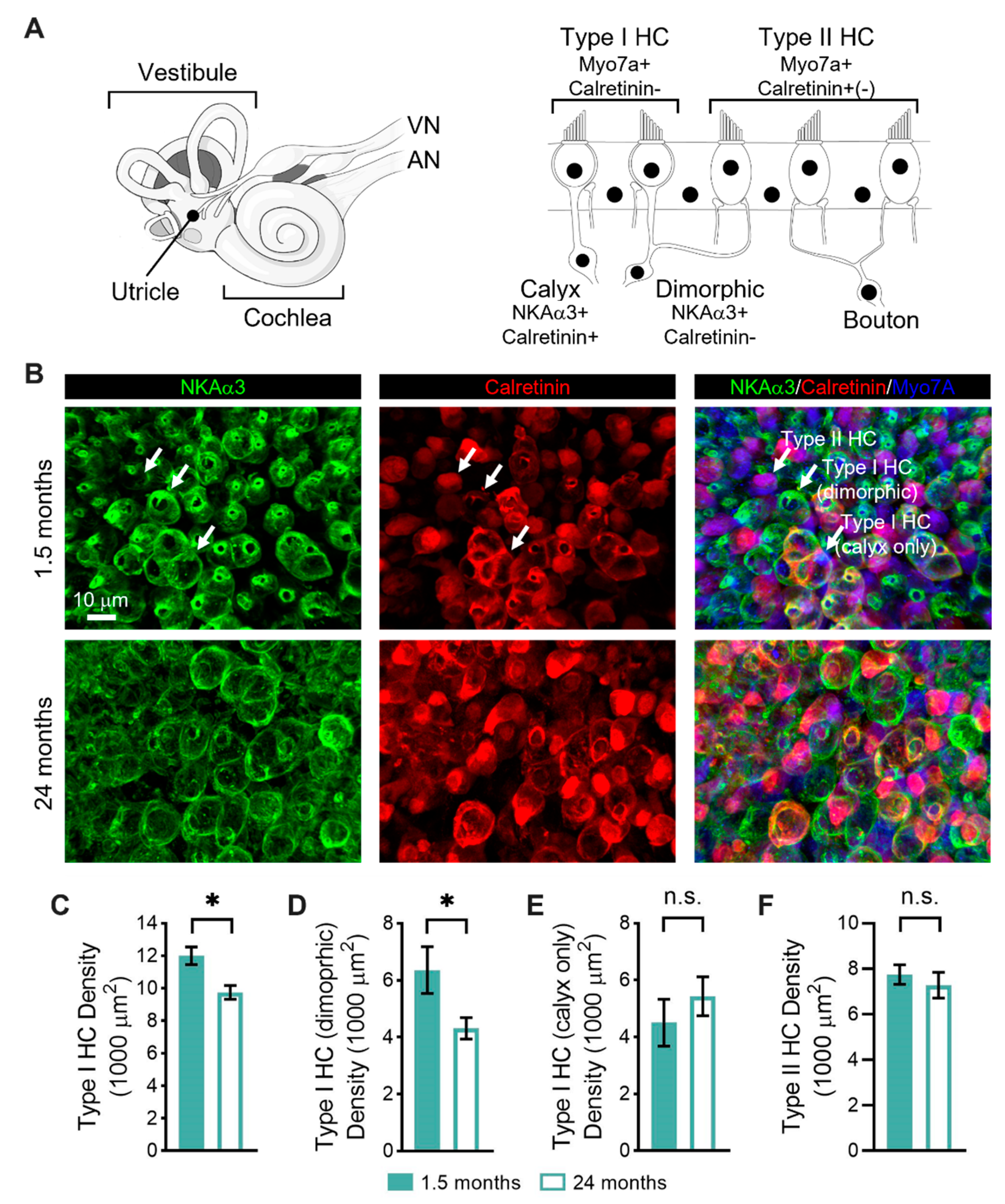
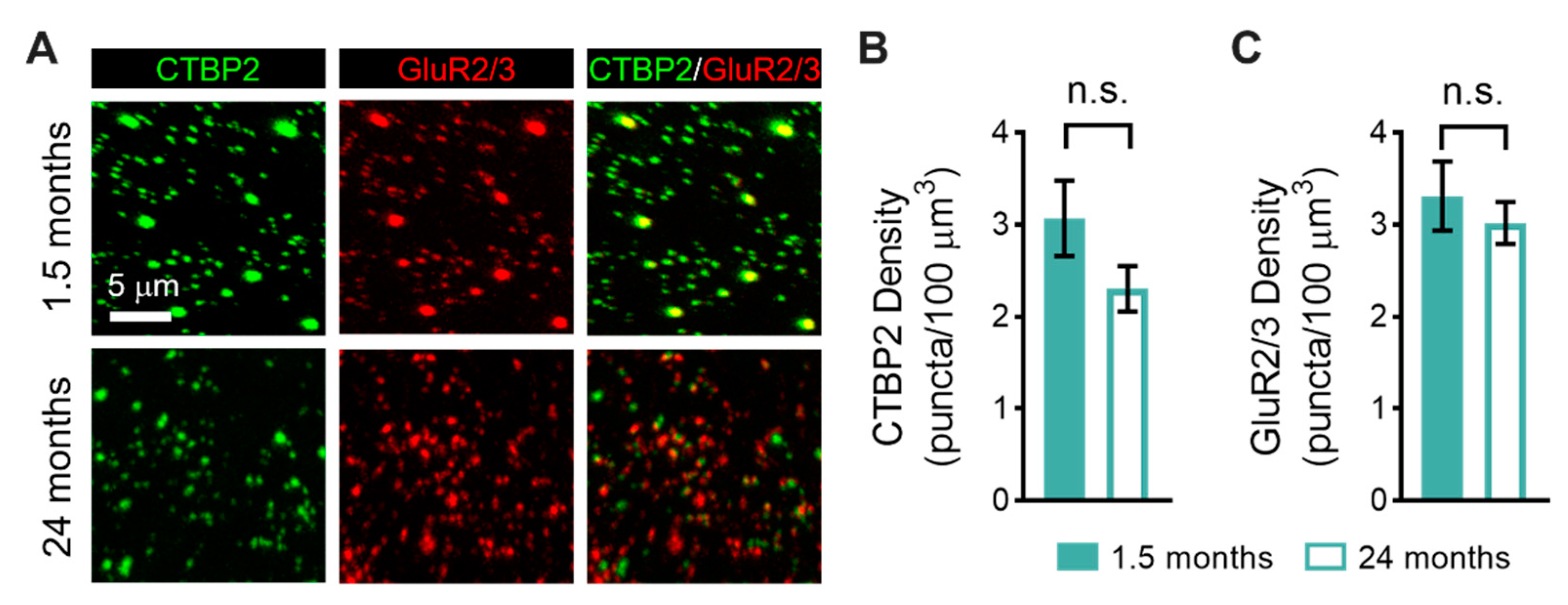
3.2. Age-Related Changes in the Sensorineural Structure of the Vestibular Sensory Epithelia
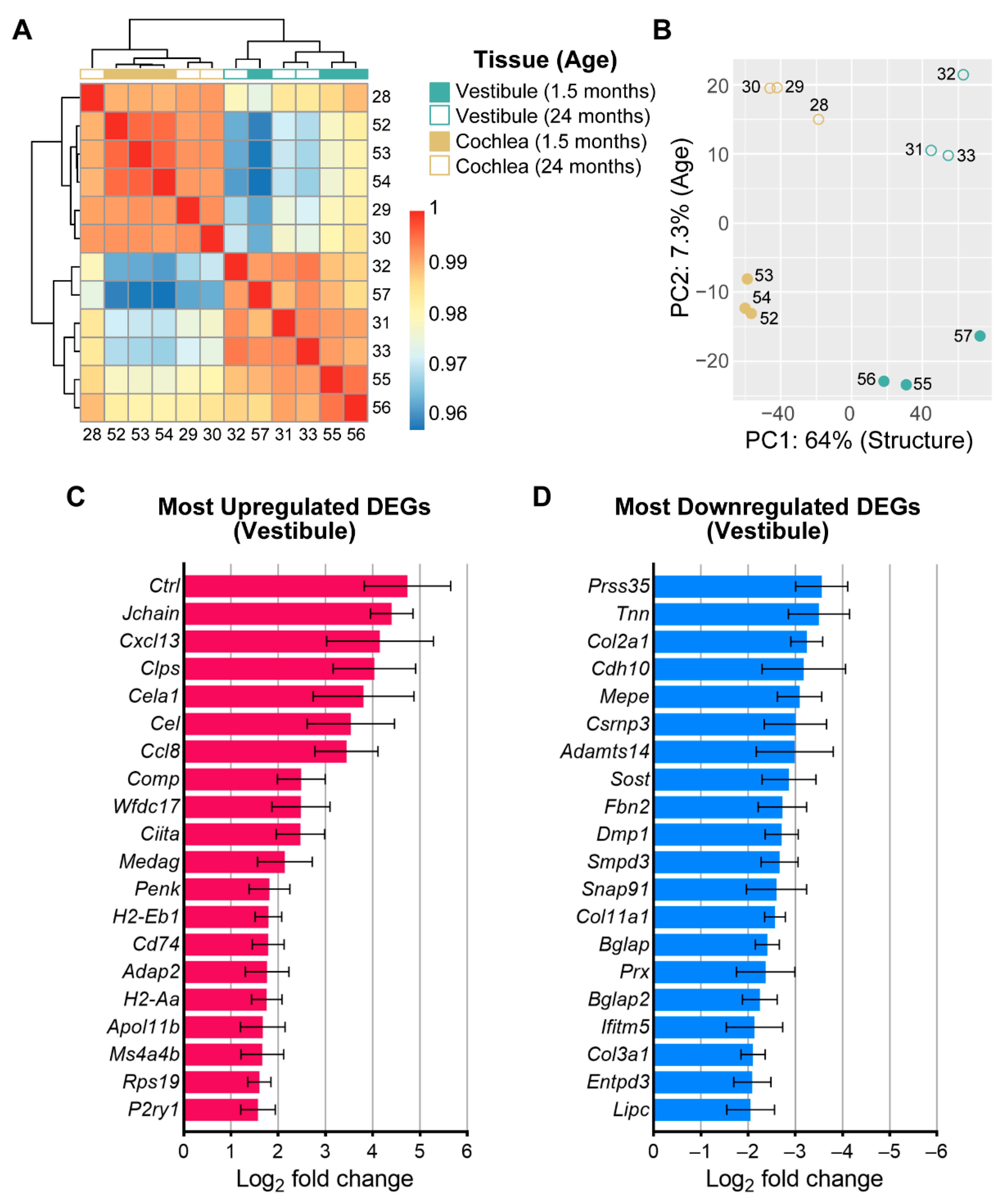
3.3. Age-Related Changes in Gene Expression in the Vestibule
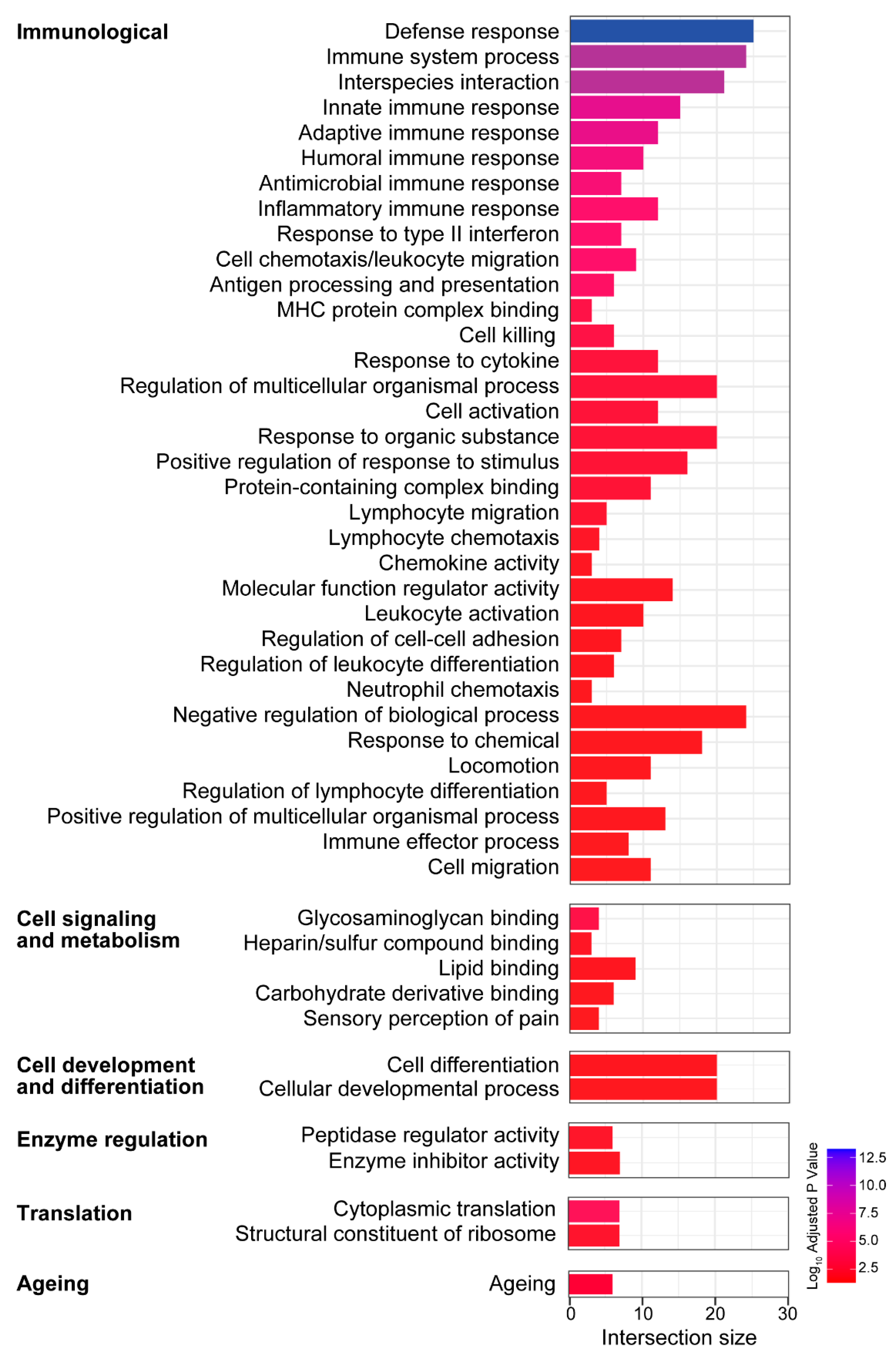
3.4. Biological Processes and Molecular Functions Associated with the Age-Related Changes in Gene Expression in the Vestibule
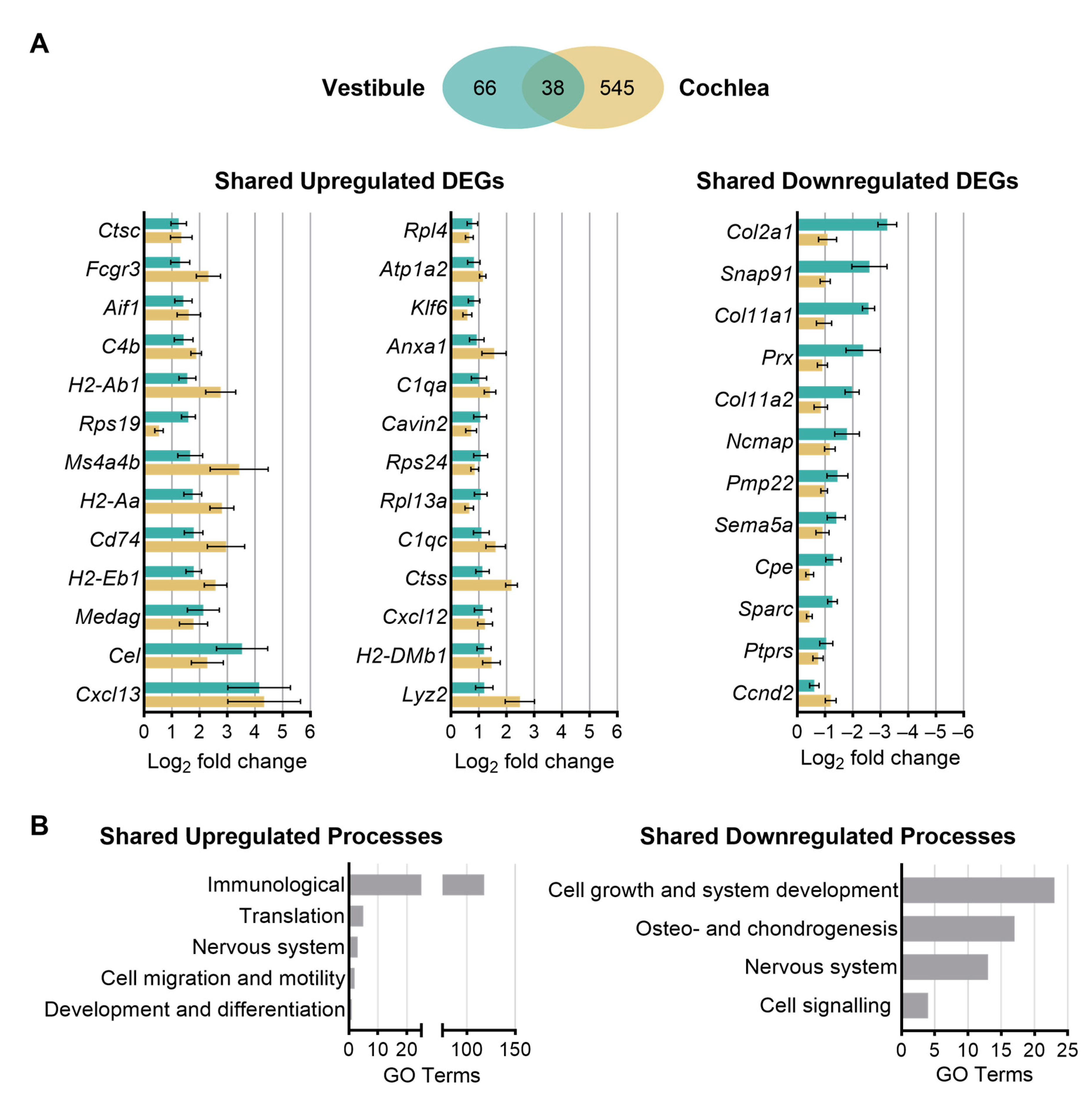
3.5. Shared and Unique Age-Related Changes in Gene Expression between the Vestibule and Cochlea
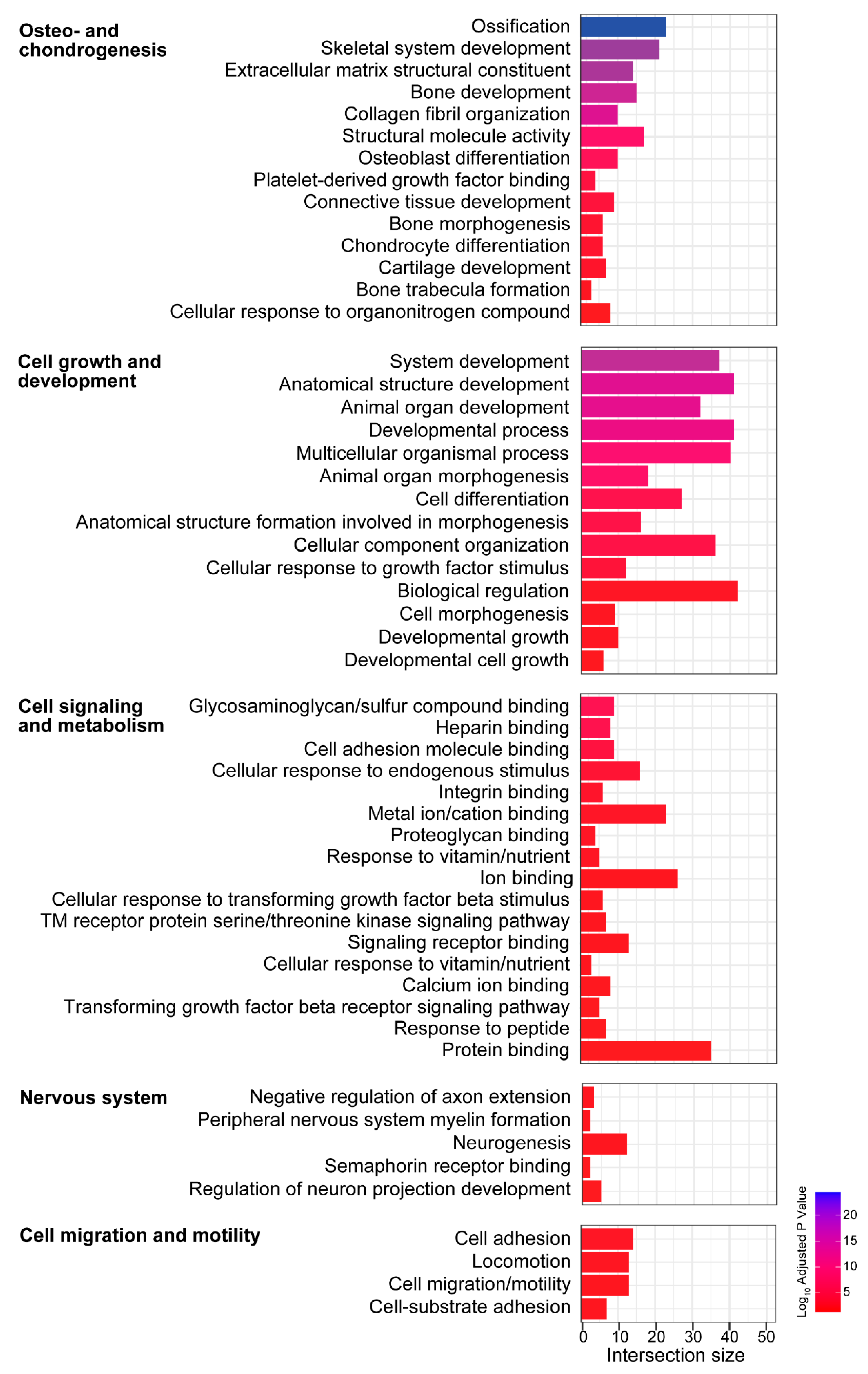
3.6. Biological Processes and Molecular Functions Associated with Age-Related Changes in Gene Expression Shared between the Vestibule and Cochlea
4. Discussion
4.1. Peripheral Vestibular Function Declines with Age
4.2. Type I Vestibular Hair Cells Decline in Abundance with Age
4.3. Age-Related Transcriptomic Changes in the Vestibule Implicate Immune and Inflammation-Related Processes
4.4. Fewer Age-Related Transcriptomic Changes in the Vestibule Compared to Cochlea
4.5. Limitations
4.6. Clinical Implications and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, J.R.; Shubayev, I.; Baloh, R.W.; Demer, J.L. Impairments in the initial horizontal vestibulo-ocular reflex of older humans. Exp. Brain Res. 2001, 137, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, Y.; Carey, J.P.; Della Santina, C.C.; Schubert, M.C.; Minor, L.B. Disorders of balance and vestibular function in US adults: Data from the National Health and Nutrition Examination Survey, 2001–2004. Arch. Intern. Med. 2009, 169, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Gazzola, J.M.; Ganança, F.F.; Aratani, M.C.; Perracini, M.R.; Ganança, M.M. Circumstances and consequences of falls in elderly people with vestibular disorder. Braz. J. Otorhinolaryngol. 2006, 72, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Tinetti, M.E. Clinical practice. Preventing falls in elderly persons. N. Engl. J. Med. 2003, 348, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Tinetti, M.E.; Williams, C.S. Falls, injuries due to falls, and the risk of admission to a home. N. Engl. J. Med. 1997, 337, 1279–1284. [Google Scholar] [CrossRef]
- Herdman, S.J.; Blatt, P.; Schubert, M.C.; Tusa, R.J. Falls in patients with vestibular deficits. Am. J. Otol. 2000, 21, 847–851. [Google Scholar]
- Dobbels, B.; Mertens, G.; Gilles, A.; Claes, A.; Moyaert, J.; van de Berg, R.; Van de Heyning, P.; Vanderveken, O.; Van Rompaey, V. Cognitive Function in Acquired Bilateral Vestibulopathy: A Cross-Sectional Study on Cognition, Hearing, and Vestibular Loss. Front. Neurosci. 2019, 13, 340. [Google Scholar] [CrossRef]
- Semenov, Y.R.; Bigelow, R.T.; Xue, Q.L.; du Lac, S.; Agrawal, Y. Association Between Vestibular and Cognitive Function in U.S. Adults: Data from the National Health and Nutrition Examination Survey. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 243–250. [Google Scholar] [CrossRef]
- Paplou, V.; Schubert, N.M.A.; Pyott, S.J. Age-Related Changes in the Cochlea and Vestibule: Shared Patterns and Processes. Front. Neurosci. 2021, 15, 680856. [Google Scholar] [CrossRef]
- Agrawal, Y.; Ward, B.K.; Minor, L.B. Vestibular dysfunction: Prevalence, impact and need for targeted treatment. J. Vestib. Res. 2013, 23, 113–117. [Google Scholar] [CrossRef]
- Khan, S.; Chang, R. Anatomy of the vestibular system: A review. NeuroRehabilitation 2013, 32, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Lysakowski, A.; Gaboyard-Niay, S.; Calin-Jageman, I.; Chatlani, S.; Price, S.D.; Eatock, R.A. Molecular microdomains in a sensory terminal, the vestibular calyx ending. J. Neurosci. 2011, 31, 10101–10114. [Google Scholar] [CrossRef] [PubMed]
- Barmack, N.H. Central vestibular system: Vestibular nuclei and posterior cerebellum. Brain Res. Bull. 2003, 60, 511–541. [Google Scholar] [CrossRef] [PubMed]
- Arshad, Q.; Seemungal, B.M. Age-Related Vestibular Loss: Current Understanding and Future Research Directions. Front. Neurol. 2016, 7, 231. [Google Scholar] [CrossRef] [PubMed]
- Johnsson, L.G. Degenerative changes and anomalies of the vestibular system in man. Laryngoscope 1971, 81, 1682–1694. [Google Scholar] [CrossRef] [PubMed]
- Brosel, S.; Laub, C.; Averdam, A.; Bender, A.; Elstner, M. Molecular aging of the mammalian vestibular system. Ageing Res. Rev. 2016, 26, 72–80. [Google Scholar] [CrossRef]
- Rosenhall, U. Degenerative patterns in the aging human vestibular neuro-epithelia. Acta Oto-Laryngol. 1973, 76, 208–220. [Google Scholar] [CrossRef]
- Nakayama, M.; Helfert, R.H.; Konrad, H.R.; Caspary, D.M. Scanning electron microscopic evaluation of age-related changes in the rat vestibular epithelium. Otolaryngol. Head Neck Surg. 1994, 111, 799–806. [Google Scholar]
- Iwasaki, S.; Smulders, Y.E.; Burgess, A.M.; McGarvie, L.A.; Macdougall, H.G.; Halmagyi, G.M.; Curthoys, I.S. Ocular vestibular evoked myogenic potentials to bone conducted vibration of the midline forehead at Fz in healthy subjects. Clin. Neurophysiol. 2008, 119, 2135–2147. [Google Scholar] [CrossRef]
- Iwasaki, S.; Chihara, Y.; Smulders, Y.E.; Burgess, A.M.; Halmagyi, G.M.; Curthoys, I.S.; Murofushi, T. The role of the superior vestibular nerve in generating ocular vestibular-evoked myogenic potentials to bone conducted vibration at Fz. Clin. Neurophysiol. 2009, 120, 588–593. [Google Scholar] [CrossRef]
- Agrawal, Y.; Zuniga, M.G.; Davalos-Bichara, M.; Schubert, M.C.; Walston, J.D.; Hughes, J.; Carey, J.P. Decline in semicircular canal and otolith function with age. Otol. Neurotol. 2012, 33, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Wan, G.; Ji, L.; Schrepfer, T.; Gong, S.; Wang, G.-P.; Corfas, G. Synaptopathy as a Mechanism for Age-Related Vestibular Dysfunction in Mice. Front. Aging Neurosci. 2019, 11, 156. [Google Scholar] [CrossRef] [PubMed]
- Watson, N.; Ding, B.; Zhu, X.; Frisina, R.D. Chronic inflammation–inflammaging–in the ageing cochlea: A novel target for future presbycusis therapy. Ageing Res. Rev. 2017, 40, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Verschuur, C.A.; Dowell, A.; Syddall, H.E.; Ntani, G.; Simmonds, S.J.; Baylis, D.; Gale, C.R.; Walsh, B.; Cooper, C.; Lord, J.M.; et al. Markers of inflammatory status are associated with hearing threshold in older people: Findings from the Hertfordshire ageing study. Age Ageing 2012, 41, 92–97. [Google Scholar] [CrossRef]
- Verschuur, C.; Agyemang-Prempeh, A.; Newman, T.A. Inflammation is associated with a worsening of presbycusis: Evidence from the MRC national study of hearing. Int. J. Audiol. 2014, 53, 469–475. [Google Scholar] [CrossRef]
- Schubert, N.M.A.; van Tuinen, M.; Pyott, S.J. Transcriptome-Guided Identification of Drugs for Repurposing to Treat Age-Related Hearing Loss. Biomolecules 2022, 12, 498. [Google Scholar] [CrossRef]
- Mock, B.E.; Vijayakumar, S.; Pierce, J.; Jones, T.A.; Jones, S.M. Differential effects of Cdh23 (753A) on auditory and vestibular functional aging in C57BL/6J mice. Neurobiol. Aging 2016, 43, 13–22. [Google Scholar] [CrossRef]
- Greguske, E.A.; Llorens, J.; Pyott, S.J. Assessment of cochlear toxicity in response to chronic 3,3′-iminodipropionitrile in mice reveals early and reversible functional loss that precedes overt histopathology. Arch. Toxicol. 2021, 95, 1003–1021. [Google Scholar] [CrossRef]
- Braude, J.P.; Vijayakumar, S.; Baumgarner, K.; Laurine, R.; Jones, T.A.; Jones, S.M.; Pyott, S.J. Deletion of Shank1 has minimal effects on the molecular composition and function of glutamatergic afferent postsynapses in the mouse inner ear. Hear. Res. 2015, 321, 52–64. [Google Scholar] [CrossRef]
- Schuth, O.; McLean, W.J.; Eatock, R.A.; Pyott, S.J. Distribution of Na,K-ATPase alpha subunits in rat vestibular sensory epithelia. J. Assoc. Res. Otolaryngol. 2014, 15, 739–754. [Google Scholar] [CrossRef]
- Reijntjes, D.O.J.; Lee, J.H.; Park, S.; Schubert, N.M.A.; van Tuinen, M.; Vijayakumar, S.; Jones, T.A.; Jones, S.M.; Gratton, M.A.; Xia, X.-M.; et al. Sodium-activated potassium channels shape peripheral auditory function and activity of the primary auditory neurons in mice. Sci. Rep. 2019, 9, 2573. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Use R! Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Kolde, R. Pheatmap: Pretty Heatmaps, Version 1.0.12. 2015. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 20 April 2022).
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef] [PubMed]
- Nazareth, A.M.; Jones, T.A. Central and peripheral components of short latency vestibular responses in the chicken. J. Vestib. Res. 1998, 8, 233–252. [Google Scholar] [CrossRef]
- Chihara, Y.; Wang, V.; Brown, D.J. Evidence for the utricular origin of the vestibular short-latency-evoked potential (VsEP) to bone-conducted vibration in guinea pig. Exp. Brain Res. 2013, 229, 157–170. [Google Scholar] [CrossRef]
- Geng, R.; Geller, S.F.; Hayashi, T.; Ray, C.A.; Reh, T.A.; Bermingham-McDonogh, O.; Jones, S.M.; Wright, C.G.; Melki, S.; Imanishi, Y.; et al. Usher syndrome IIIA gene clarin-1 is essential for hair cell function and associated neural activation. Hum. Mol. Genet. 2009, 18, 2748–2760. [Google Scholar] [CrossRef]
- Krey, J.F.; Krystofiak, E.S.; Dumont, R.A.; Vijayakumar, S.; Choi, D.; Rivero, F.; Kachar, B.; Jones, S.M.; Barr-Gillespie, P.G. Plastin 1 widens stereocilia by transforming actin filament packing from hexagonal to liquid. J. Cell Biol. 2016, 215, 467–482. [Google Scholar] [CrossRef]
- Ono, K.; Keller, J.; Ramírez, O.L.; Garrido, A.G.; Zobeiri, O.A.; Chang, H.H.V.; Vijayakumar, S.; Ayiotis, A.; Duester, G.; Della Santina, C.C.; et al. Retinoic acid degradation shapes zonal development of vestibular organs and sensitivity to transient linear accelerations. Nat. Commun. 2020, 11, 63. [Google Scholar] [CrossRef]
- Hasson, T.; Gillespie, P.G.; Garcia, J.A.; MacDonald, R.B.; Zhao, Y.-D.; Yee, A.G.; Mooseker, M.S.; Corey, D.P. Unconventional myosins in inner-ear sensory epithelia. J. Cell Biol. 1997, 137, 1287–1307. [Google Scholar] [CrossRef]
- Desai, S.S.; Ali, H.; Lysakowski, A. Comparative morphology of rodent vestibular periphery. II. Cristae ampullares. J. Neurophysiol. 2005, 93, 267–280. [Google Scholar] [CrossRef]
- Desmadryl, G.; Dechesne, C.J. Calretinin immunoreactivity in chinchilla and guinea pig vestibular end organs characterizes the calyx unit subpopulation. Exp. Brain Res. 1992, 89, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Leonard, R.B.; Kevetter, G.A. Molecular probes of the vestibular nerve. I. Peripheral termination patterns of calretinin, calbindin and peripherin containing fibers. Brain Res. 2002, 928, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Blake, J.A.; Baldarelli, R.; Kadin, J.A.; Richardson, J.E.; Smith, C.L.; Bult, C.J. Mouse Genome Database (MGD): Knowledgebase for mouse-human comparative biology. Nucleic Acids Res. 2021, 49, D981–D987. [Google Scholar] [CrossRef]
- Gao, S.-H.; Liu, S.-Z.; Wang, G.-Z.; Zhou, G.-B. CXCL13 in Cancer and Other Diseases: Biological Functions, Clinical Significance, and Therapeutic Opportunities. Life 2021, 11, 1282. [Google Scholar] [CrossRef] [PubMed]
- Beswick, E.J.; Reyes, V.E. CD74 in antigen presentation, inflammation, and cancers of the gastrointestinal tract. World J. Gastroenterol. 2009, 15, 2855–2861. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Jones, T.A.; Johnson, K.R.; Yu, H.; Erway, L.C.; Zheng, Q.Y. A comparison of vestibular and auditory phenotypes in inbred mouse strains. Brain Res. 2006, 1091, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Kohrman, D.C.; Wan, G.; Cassinotti, L.; Corfas, G. Hidden Hearing Loss: A Disorder with Multiple Etiologies and Mechanisms. Cold Spring Harb. Perspect. Med. 2020, 10, a035493. [Google Scholar] [CrossRef]
- Zalewski, C.K. Aging of the Human Vestibular System. Semin. Hear. 2015, 36, 175–196. [Google Scholar] [CrossRef]
- Maroto, A.F.; Borrajo, M.; Prades, S.; Callejo, A.; Amilibia, E.; Pérez-Grau, M.; Roca-Ribas, F.; Castellanos, E.; Barrallo-Gimeno, A.; Llorens, J. The vestibular calyceal junction is dismantled following subchronic streptomycin in rats and sensory epithelium stress in humans. Arch. Toxicol. 2023, 97, 1943–1961. [Google Scholar] [CrossRef]
- Sadeghi, S.G.; Pyott, S.J.; Yu, Z.; Glowatzki, E. Glutamatergic signaling at the vestibular hair cell calyx synapse. J. Neurosci. 2014, 34, 14536–14550. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Demontiero, O.; Vidal, C.; Duque, G. Aging and bone loss: New insights for the clinician. Ther. Adv. Musculoskelet. Dis. 2012, 4, 61–76. [Google Scholar] [CrossRef]
- Wright, N.C.; Looker, A.C.; Saag, K.G.; Curtis, J.R.; Delzell, E.S.; Randall, S.; Dawson-Hughes, B. The recent prevalence of osteoporosis and low bone mass in the United States based on bone mineral density at the femoral neck or lumbar pine. J. Bone Miner. Res. 2014, 29, 2520–2526. [Google Scholar] [CrossRef]
- Schubert, N.M.A.; Roelofs, C.G.; Free, R.H.; Wiersinga-Post, J.E.C.; Pyott, S.J. Age-related High-frequency Hearing Loss Is Not Associated with Horizontal Semicircular Canal Function. Ear Hear. 2022, 43, 1845–1852. [Google Scholar] [CrossRef]
- Shiga, A.; Nakagawa, T.; Nakayama, M.; Endo, T.; Iguchi, F.; Kim, T.-S.; Naito, Y.; Ito, J. Aging effects on vestibulo-ocular responses in C57BL/6 mice: Comparison with alteration in auditory function. Audiol. Neurotol. 2005, 10, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Matern, M.; Vijayakumar, S.; Margulies, Z.; Milon, B.; Song, Y.; Elkon, R.; Zhang, X.; Jones, S.M.; Hertzano, R. Gfi1(Cre) mice have early onset progressive hearing loss and induce recombination in numerous inner ear non-hair cells. Sci. Rep. 2017, 7, 42079. [Google Scholar] [CrossRef] [PubMed]
- Orvis, J.; Gottfried, B.; Kancherla, J.; Adkins, R.S.; Song, Y.; Dror, A.A.; Olley, D.; Rose, K.; Chrysostomou, E.; Kelly, M.C.; et al. gEAR: Gene Expression Analysis Resource portal for community-driven, multi-omic data exploration. Nat. Methods 2021, 18, 843–844. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paplou, V.G.; Schubert, N.M.A.; van Tuinen, M.; Vijayakumar, S.; Pyott, S.J. Functional, Morphological and Molecular Changes Reveal the Mechanisms Associated with Age-Related Vestibular Loss. Biomolecules 2023, 13, 1429. https://doi.org/10.3390/biom13091429
Paplou VG, Schubert NMA, van Tuinen M, Vijayakumar S, Pyott SJ. Functional, Morphological and Molecular Changes Reveal the Mechanisms Associated with Age-Related Vestibular Loss. Biomolecules. 2023; 13(9):1429. https://doi.org/10.3390/biom13091429
Chicago/Turabian StylePaplou, Vasiliki Georgia, Nick M. A. Schubert, Marcel van Tuinen, Sarath Vijayakumar, and Sonja J. Pyott. 2023. "Functional, Morphological and Molecular Changes Reveal the Mechanisms Associated with Age-Related Vestibular Loss" Biomolecules 13, no. 9: 1429. https://doi.org/10.3390/biom13091429
APA StylePaplou, V. G., Schubert, N. M. A., van Tuinen, M., Vijayakumar, S., & Pyott, S. J. (2023). Functional, Morphological and Molecular Changes Reveal the Mechanisms Associated with Age-Related Vestibular Loss. Biomolecules, 13(9), 1429. https://doi.org/10.3390/biom13091429