
MicroRNA-7: A New Intervention Target for Inflammation and Related Diseases

 and
and
Abstract
1. Introduction
2. Regulatory Mechanism of miR-7 Expression
2.1. The DNA Methylation
2.2. circRNAs
2.3. lncRNA
2.4. Transcription Factors (TFs)
2.5. Target-Directed miRNA Degradation (TDMD)
2.6. Other Factors
3. miRNA-7 and Inflammatory Diseases

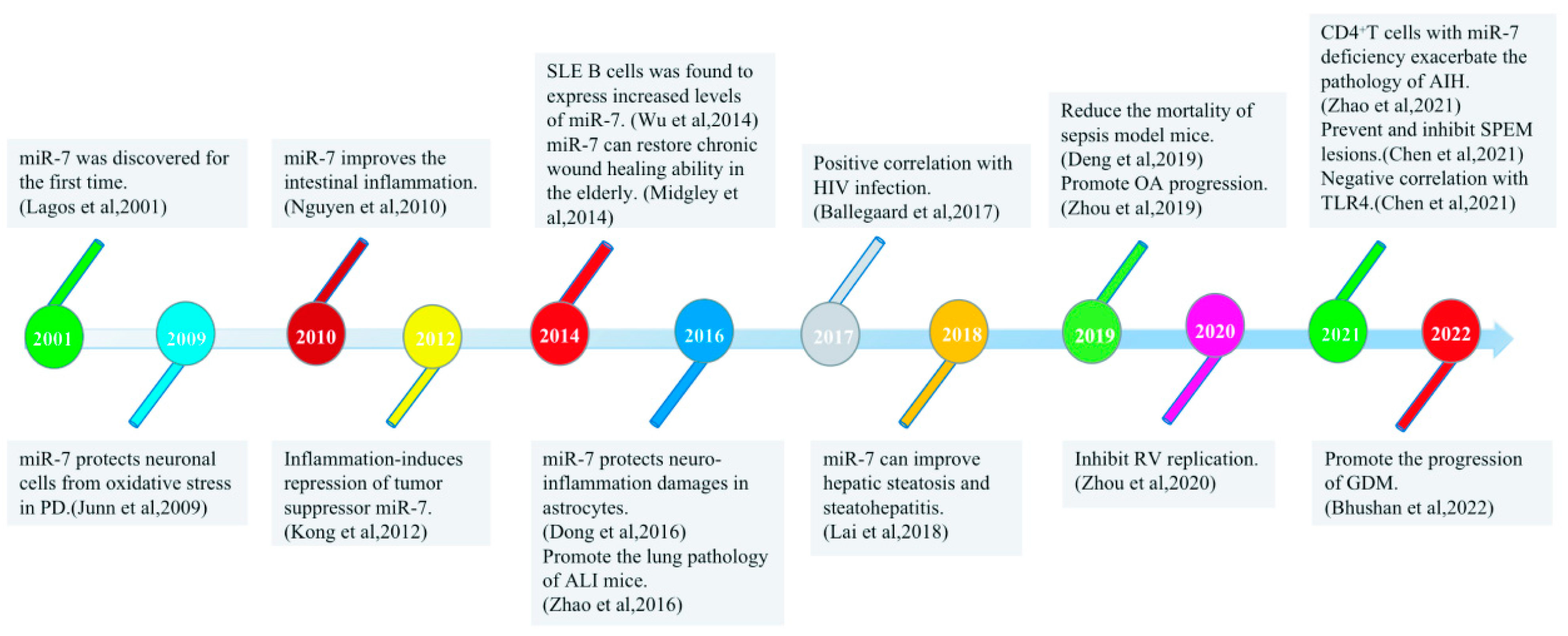{kind=link}
{kind=link}
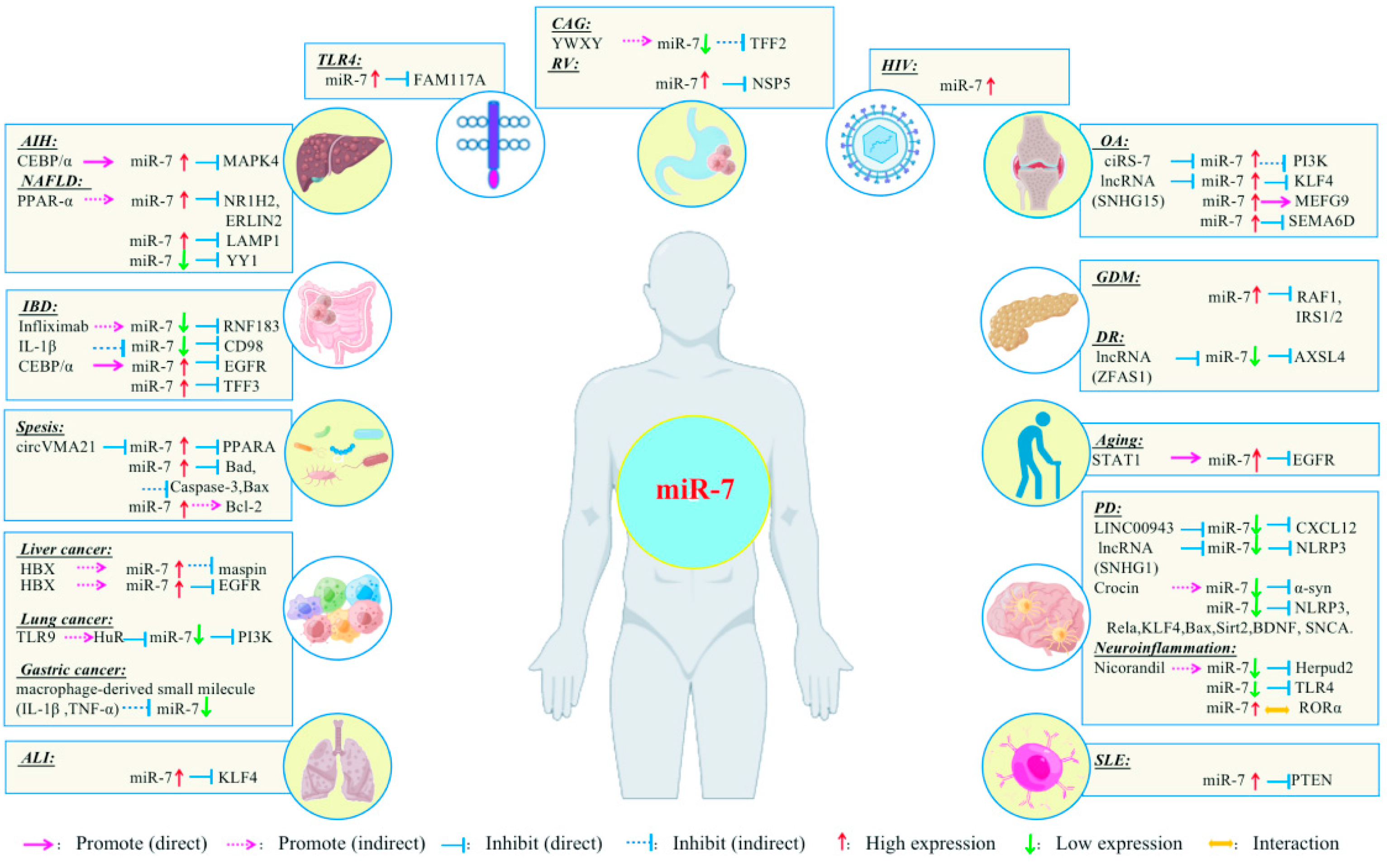{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inflammatory Related Diseases | Genes/Drugs That Regulate miR-7 | Target Genes of miR-7 | Immune Cells or Molecules | Outcome | Expression Level of miR-7 | Remark | Ref. |
|---|---|---|---|---|---|---|---|
| Lung cancer | HuR | PI3K | unknown | Inhibit the growth and metastasis of lung cancer cells by inhibiting the TLR9 signaling pathway | (↓) 95D cell | miR-7 mimic transfected (in vitro) plasmids transfected (in vivo) | [14,15] |
| Aging | STAT1 | EGFR | TGF-β1, IFN-γ | Impair age-associated loss of EGFR and hyaluronan-dependent differentiation in fibroblasts. | (↑) in aged fibroblasts | miR-7 mimic/locked nucleic acids (LNA) targeting miR-7 transfected (in vitro) | [26,36] |
| AIH | C/EBPα | MAPK4 | CD4+T, IFN-γ, IL-4 | CD4+T cells with miR-7 deficiency exacerbate the pathology of AIH | (↑) in liver tissue in mice | plasmids transfected (in vitro) | [27] |
| NAFLD | PPAR-α |
ERLIN2, NR1H2 | unknown | Promote hepatocellular lipid accumulation | (↑) in fatty liver in mice | miR-7 mimic transfected (in vitro) | [35] |
| LAMP1 | IL-1β | Promote microvascular endothelial hyperpermeability | (↑) in the sEVs of fatty liver | hepatic small extracellular vesicles (sEVs) | [37] | ||
| YY1 |
IL-1β, IL-6, TNF-α, IFN-γ | Improve hepatic steatosis and steatohepatitis | (↓) Nonalcoholic fatty liver in zebrafish in mice | miR-7 mimic transfected (in vitro) | [38] | ||
| PD | LINC00943 | CXCL12 | IL-1β, TNF-α | Promote cell viability, repress apoptosis and the inflammatory response | (↓) in MPP+-inducted SK-N-SH cells | miR-7 mimic/inhibitor transfected (in vitro) | [24] |
| SNHG1 | NLRP3 | Microglial, IL-6, TNF-α, IL-1β, IL-18 | Inhibit the activation of NLRP3 inflammatories | (↓) in LPS-induced BV2 cells | miR-7 mimic/inhibitor transfected (in vitro) | [25] | |
| NLRP3 | Microglial, IL-1β | Inhibit the activation of NLRP3 inflammasome and attenuate dopaminergic neuronal degeneration. | (↓) in the serum samples of PD patients | miR-7 mimic transfected (in vitro) stereotactically injected miR-7 mimics (in vivo) | [39,40] | ||
| RelA | TNF-α | Protect neurons, prevent cell death and promote glycolysis. | (↓) in MPP(+) -inducted SH-SY5Y cells | plasmids transfected (in vitro) | [41,42] | ||
| KLF4 | unknown | Inhibit neuronal cells apoptosis | (↓) in MPP(+) -inducted SH-SY5Y cells | miR-7 mimic transfected (in vitro) | [43] | ||
| Bax, Sirt2 | unknown | Inhibit neuronal cells apoptosis | (↓) in MPP(+) -inducted SH-SY5Y cells | miR-7 mimic transfected (in vitro) | [44] | ||
| Crocin | α-syn | unknown | miR-7 has a neuroprotective effect | (↓) in striatal tissue in mice | Unknown | [45] | |
| α-syn | unknown | Protect neuronal cells from oxidative stress | (↓) in the MPTP-induced neurotoxin model of PD in cultured cells and in mice | plasmids transfected (in vitro) | [46] | ||
| BDNF | unknown | a strong self-protective mechanism in rats at the early stage of PD. | (↓) in peripheral blood of rats with Atrazine-induced PD | Unknown | [47] | ||
| SNCA | unknown | Protect neuronal cells | (↓) in the substantia nigra of PD patients | miR-7 lentiviral vector transfected (in vitro) stereotactically injected miR-7 lentiviral vector (in vivo) | [48] | ||
|
Neuro- inflammation | TLR4 | Microglial, TLR4, IL-1β, TNF-α, IL-8 | Alleviate the inflammatory response | (↓) in patients with intracerebral hemorrhage | miR-7 mimic/inhibitor transfected (in vitro) mixture of the rAAV9-ZsGreen-miR-7 virus transfection agent (adenovirus vector)were injected into the hematoma region of the rat (in vivo) | [49] | |
| Nicorandil | Herpud2 | IL-1β, TNF-α | Reduce the inflammatory response and astrocyte damage | (↓) in oxygen-glucose deprivation inducted astrocytes | miR-7 mimic transfected (in vitro) | [50] | |
| RORα | Microglial, IL-1β, TNF-α, IL-6, TGF-β | Negatively control the pathology of BTI | (↑) in brain tissue in BTI mice | miR-7 mimic/inhibitor transfected (in vitro) | [51] | ||
| IBD | IL-1β | CD98 | IL-1β | Improve the intestinal inflammation | (↓) in inflamed colon tissues of crohn patients | plasmids transfected (in vitro) | [52] |
| Infliximab | RNF183 |
IL-1β, TNF-α, IL-6, IL-8 | Alleviate the process of IBD | (↓) in inflamed colon tissues of IBD patients and colitic mice | miR-7 mimic/inhibitor transfected (in vitro) | [53] | |
| TFF3 | unknown | Promote the process of IBD | (↑) in the lesional tissue of IBD patients | miR-7 mimic/inhibitor transfected (in vitro) | [54,55] | ||
| C/EBPα | EGFR |
IL-6, IL-10, TNF-α, TGF-β, CD4+T, B, CD8+T | Promote the process of IBD | (↑) in colonic IECs in colitis of IBD patients | miR-7 mimic/inhibitor transfected (in vitro) IEC-specific miR-7 silencing expression vector tail vein injected (in vivo) | [56] | |
| Sepsis |
Bad, Bax, Caspase-3, BCL-2 | T | Inhibit T lymphocyte apoptosis and reduce the mortality of sepsis model mice | (↑) in plasma circulating exosomes in sepsis patients | plasmids/adenovirus transfected (in vitro) Exosomes(in vivo) | [57] | |
| circVMA21 | PPAR-α | unknown | Promote the process of sepsis-engendered AKI | (↑) in sepsis patients’ serums and LPS-stimulated HK2 cells | miR-7 mimic transfected (in vitro) | [58] | |
| SLE | PTEN | B, Tfh, IL-21 | Promotes disease manifestations in MRLlpr/lpr lupus mice, and abnormal splenic B cell subtypes | (↑) in B cells in SLE patients | miR-7 antagomir transfected (in vitro/vivo) | [59,60] | |
| OA | ciRS-7 | PI3K | IL-1β, IL-17A | Promote cartilage degradation and autophagy defects | (↑) in blood samples from OA patients | miR-7 mimic/siRNA transfected (in vitro) | [61,62] |
| MEGF9 | IL-1β | Exacerbate the OA process. | (↑) in blood samples from OA patients | miR-7 mimic/siRNA transfected (in vitro) injected miR-7 lentiviral vector (in vivo) | [63] | ||
| SNHG15 | KLF4 | IL-1β | Promote OA progression | (↑) in human OA knee cartilage tissues | miR-7 mimic/inhibitor transfected (in vitro) | [64] | |
| SEMA6D | unknown | Promote OA progression | (↑) in IL-1β-induced osteoarthritic C28/I2 chondrocytes | miR-7 mimic/inhibitor transfected (in vitro/vivo) | [65] | ||
| Gastric tumors |
macrophage derived factors | unknown | IL-1β, TNF-α | Inflammation-induced repression of miR-7 in gastric tumor cells | (↓) in human gastric cancer tissue | miR-7 mimic transfected (in vitro) | [66] |
| Liver cancer | HBx | Maspin | unknown | Promotes HCC progression | (↑) in HBx-expressing HCC cells | miR-7 inhibitor transfected (in vitro) | [67] |
| HBx | EGFR | unknown | Inhibit HCC progression | (↑) in HBx-expressing HCC cells | miR-7 inhibitor transfected (in vitro) | [68] | |
| ALI | KLF4 |
IL-1β, TNF-α, IFN-γ, CD4+T DC, CD8+T | Promote the lung pathology of ALI mice. | (↑) in the lung tissues of LPS-induced mouse ALI models | Unknown | [69] | |
| GDM | RAF1, IRS1/2 | IL-6R | Cause chronic low-grade inflammation and promote the progression of GDM. | (↑) in maternal blood of GDM patients | Unknown | [70] | |
| RV | NSP5 | unknown | Inhibit RV replication. | (↑) in RV-infected cells | miR-7 mimic/inhibitor transfected (in vitro) intragastric inoculation of miR-7 agomir/antagomir (in vivo) | [71] | |
| CAG | TFF2 | unknown | Inhibit the development and progression of SPEM. | (↓) in human SPEM lesions/gastric cancer tissue | Unknown | [72] | |
| HIV | unknown | IL-6 | miR-7 was significantly upregulated in HIV-1 infection after treatment. | (↑) in PBMCs from HIV-1-infected individuals | Unknown | [73] | |
|
Immuno- inflammatory | TLR4 | FAM177A |
Macrophage, TLR4, IL-1β, TNF-α, IL-6, IL-12 | Negatively regulate TLR4 signaling pathway. | (↑) in TLR4 signalling-activated bone marrow-derived macrophages stimulated by LPS | miR-7 mimic transfected (in vitro) | [74] |
3.1. Hepatitis
3.1.1. Autoimmune Hepatitis (AIH)
3.1.2. Nonalcoholic Fatty Liver Disease (NAFLD)
3.2. Parkinson’s Disease (PD)
3.3. Neuroinflammation
3.4. Inflammatory Bowel Disease (IBD)
3.5. Sepsis
3.6. Systemic Lupus Erythematosus (SLE)
3.7. Osteoarthritis (OA)
3.8. Aging
3.9. Inflammation Drives Malignancy
3.10. Other Inflammation-Related Diseases
4. Summary and Prospect
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gaber, T.; Strehl, C.; Buttgereit, F. Metabolic regulation of inflammation. Nat. Rev. Rheumatol. 2017, 13, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V. Autophagy in inflammation, infection, and immunometabolism. Immunity 2021, 54, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Eulalio, A.; Huntzinger, E.; Izaurralde, E. Getting to the root of miRNA-mediated gene silencing. Cell 2008, 132, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Yates, L.A.; Norbury, C.J.; Gilbert, R.J. The long and short of microRNA. Cell 2013, 153, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Krump, N.A.; You, J. Molecular mechanisms of viral oncogenesis in humans. Nat. Rev. Microbiol. 2018, 16, 684–698. [Google Scholar] [CrossRef]
- Mann, M.; Mehta, A.; Zhao, J.L.; Lee, K.; Marinov, G.K.; Garcia-Flores, Y.; Lu, L.F.; Rudensky, A.Y.; Baltimore, D. An NF-κB-microRNA regulatory network tunes macrophage inflammatory responses. Nat. Commun. 2017, 8, 851. [Google Scholar] [CrossRef] [PubMed]
- Safari, A.; Madadi, S.; Schwarzenbach, H.; Soleimani, M.; Safari, A.; Ahmadi, M.; Soleimani, M. MicroRNAs and their Implications in CD4+ T-cells, Oligodendrocytes and Dendritic Cells in Multiple Sclerosis Pathogenesis. Curr. Mol. Med. 2023, 23, 630–647. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef]
- Zhao, J.; Tao, Y.; Zhou, Y.; Qin, N.; Chen, C.; Tian, D.; Xu, L. MicroRNA-7: A promising new target in cancer therapy. Cancer Cell Int. 2015, 15, 103. [Google Scholar] [CrossRef]
- Horsham, J.L.; Ganda, C.; Kalinowski, F.C.; Brown, R.A.; Epis, M.R.; Leedman, P.J. MicroRNA-7: A miRNA with expanding roles in development and disease. Int. J. Biochem. Cell Biol. 2015, 69, 215–224. [Google Scholar] [CrossRef]
- Morales-Martínez, M.; Vega, M.I. Role of MicroRNA-7 (MiR-7) in Cancer Physiopathology. Int. J. Mol. Sci. 2022, 23, 9091. [Google Scholar] [CrossRef]
- Xia, J.; Cao, T.; Ma, C.; Shi, Y.; Sun, Y.; Wang, Z.P.; Ma, J. miR-7 Suppresses Tumor Progression by Directly Targeting MAP3K9 in Pancreatic Cancer. Mol. Ther. Nucleic Acids 2018, 13, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Xu, L.; Jiao, S.; Wang, Y.; Cai, Y.; Liang, Y.; Zhou, Y.; Zhou, H.; Wen, Z. TLR9 signaling promotes tumor progression of human lung cancer cell in vivo. Pathol. Oncol. Res. POR 2009, 15, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wen, Z.; Zhou, Y.; Liu, Z.; Li, Q.; Fei, G.; Luo, J.; Ren, T. MicroRNA-7-regulated TLR9 signaling-enhanced growth and metastatic potential of human lung cancer cells by altering the phosphoinositide-3-kinase, regulatory subunit 3/Akt pathway. Mol. Biol. Cell 2013, 24, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Wang, C.H.; Zhou, Y.; Liao, Z.Y.; Zhu, S.F.; Hu, Y.; Chen, C.; Luo, J.M.; Wen, Z.K.; Xu, L. TLR9 signaling repressed tumor suppressor miR-7 expression through up-regulation of HuR in human lung cancer cells. Cancer Cell Int. 2013, 13, 90. [Google Scholar] [CrossRef]
- Lei, L.; Chen, C.; Zhao, J.; Wang, H.; Guo, M.; Zhou, Y.; Luo, J.; Zhang, J.; Xu, L. Targeted Expression of miR-7 Operated by TTF-1 Promoter Inhibited the Growth of Human Lung Cancer through the NDUFA4 Pathway. Mol. Ther. Nucleic Acids 2017, 6, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Sergiev, P.V.; Aleksashin, N.A.; Chugunova, A.A.; Polikanov, Y.S.; Dontsova, O.A. Structural and evolutionary insights into ribosomal RNA methylation. Nat. Chem. Biol. 2018, 14, 226–235. [Google Scholar] [CrossRef]
- Rosas-Alonso, R.; Galera, R.; Sánchez-Pascuala, J.J.; Casitas, R.; Burdiel, M.; Martínez-Cerón, E.; Vera, O.; Rodriguez-Antolin, C.; Pernía, O.; De Castro, J.; et al. Hypermethylation of Anti-oncogenic MicroRNA 7 is Increased in Emphysema Patients. Arch. Bronconeumol. 2020, 56, 506–513. [Google Scholar] [CrossRef]
- Hansen, T.B.; Kjems, J.; Damgaard, C.K. Circular RNA and miR-7 in cancer. Cancer Res. 2013, 73, 5609–5612. [Google Scholar] [CrossRef]
- Peng, L.; Yuan, X.Q.; Li, G.C. The emerging landscape of circular RNA ciRS-7 in cancer. Oncol. Rep. 2015, 33, 2669–2674. [Google Scholar] [CrossRef]
- Zhao, Y.; Alexandrov, P.N.; Jaber, V.; Lukiw, W.J. Deficiency in the Ubiquitin Conjugating Enzyme UBE2A in Alzheimer’s Disease (AD) is Linked to Deficits in a Natural Circular miRNA-7 Sponge (circRNA; ciRS-7). Genes 2016, 7, 116. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Sun, H.; Zhang, J.M.; Wang, M.; Du, X.J.; Zhang, J.L. Long non-coding RNA ANRIL down-regulates microRNA-7 to protect human trabecular meshwork cells in an experimental model for glaucoma. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3173–3182. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Z.; Yang, J.; Wang, J.; Wu, Y.; Zhu, R.; Liu, Q.; Xie, P. lncRNA ZFAS1 Positively Facilitates Endothelial Ferroptosis via miR-7-5p/ACSL4 Axis in Diabetic Retinopathy. Oxid. Med. Cell. Longev. 2022, 2022, 9004738. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.; Wang, B.; Lu, Q.; Chen, B.; Yang, H. LINC00943 knockdown exerts neuroprotective effects in Parkinson’s disease through regulates CXCL12 expression by sponging miR-7-5p. Genes Genom. 2021, 43, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Wang, T.; Qu, Q.; Kang, T.; Yang, Q. Long Noncoding RNA SNHG1 Promotes Neuroinflammation in Parkinson’s Disease via Regulating miR-7/NLRP3 Pathway. Neuroscience 2018, 388, 118–127. [Google Scholar] [CrossRef]
- Midgley, A.C.; Morris, G.; Phillips, A.O.; Steadman, R. 17β-estradiol ameliorates age-associated loss of fibroblast function by attenuating IFN-γ/STAT1-dependent miR-7 upregulation. Aging Cell 2016, 15, 531–541. [Google Scholar] [CrossRef]
- Zhao, J.; Chu, F.; Xu, H.; Guo, M.; Shan, S.; Zheng, W.; Tao, Y.; Zhou, Y.; Hu, Y.; Chen, C.; et al. C/EBPα/miR-7 Controls CD4+ T-Cell Activation and Function and Orchestrates Experimental Autoimmune Hepatitis in Mice. Hepatology 2021, 74, 379–396. [Google Scholar] [CrossRef]
- Kleaveland, B.; Shi, C.Y.; Stefano, J.; Bartel, D.P. A Network of Noncoding Regulatory RNAs Acts in the Mammalian Brain. Cell 2018, 174, 350–362.e17. [Google Scholar] [CrossRef]
- Shi, C.Y.; Kingston, E.R.; Kleaveland, B.; Lin, D.H.; Stubna, M.W.; Bartel, D.P. The ZSWIM8 ubiquitin ligase mediates target-directed microRNA degradation. Science 2020, 370, eabc9359. [Google Scholar] [CrossRef]
- Han, J.; LaVigne, C.A.; Jones, B.T.; Zhang, H.; Gillett, F.; Mendell, J.T. A ubiquitin ligase mediates target-directed microRNA decay independently of tailing and trimming. Science 2020, 370, eabc9546. [Google Scholar] [CrossRef]
- Reddy, S.D.; Ohshiro, K.; Rayala, S.K.; Kumar, R. MicroRNA-7, a homeobox D10 target, inhibits p21-activated kinase 1 and regulates its functions. Cancer Res. 2008, 68, 8195–8200. [Google Scholar] [CrossRef] [PubMed]
- LaPierre, M.P.; Lawler, K.; Godbersen, S.; Farooqi, I.S.; Stoffel, M. MicroRNA-7 regulates melanocortin circuits involved in mammalian energy homeostasis. Nat. Commun. 2022, 13, 5733. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, N.R.; de Lima Alves, F.; de Andrés-Aguayo, L.; Graf, T.; Cáceres, J.F.; Rappsilber, J.; Michlewski, G. Tissue-specific control of brain-enriched miR-7 biogenesis. Genes Dev. 2013, 27, 24–38. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, D.; Zhang, S.; Wei, X.; Song, J.; Zhang, Y.; Jin, M.; Shen, Z.; Wang, X.; Feng, Z.; et al. Host-virus interaction: The antiviral defense function of small interfering RNAs can be enhanced by host microRNA-7 in vitro. Sci. Rep. 2015, 5, 9722. [Google Scholar] [CrossRef]
- Singaravelu, R.; Quan, C.; Powdrill, M.H.; Shaw, T.A.; Srinivasan, P.; Lyn, R.K.; Alonzi, R.C.; Jones, D.M.; Filip, R.; Russell, R.S.; et al. MicroRNA-7 mediates cross-talk between metabolic signaling pathways in the liver. Sci. Rep. 2018, 8, 361. [Google Scholar] [CrossRef]
- Midgley, A.C.; Bowen, T.; Phillips, A.O.; Steadman, R. MicroRNA-7 inhibition rescues age-associated loss of epidermal growth factor receptor and hyaluronan-dependent differentiation in fibroblasts. Aging Cell 2014, 13, 235–244. [Google Scholar] [CrossRef]
- Zuo, R.; Ye, L.F.; Huang, Y.; Song, Z.Q.; Wang, L.; Zhi, H.; Zhang, M.Y.; Li, J.Y.; Zhu, L.; Xiao, W.J.; et al. Hepatic small extracellular vesicles promote microvascular endothelial hyperpermeability during NAFLD via novel-miRNA-7. J. Nanobiotechnol. 2021, 19, 396. [Google Scholar] [CrossRef]
- Lai, C.Y.; Lin, C.Y.; Hsu, C.C.; Yeh, K.Y.; Her, G.M. Liver-directed microRNA-7a depletion induces nonalcoholic fatty liver disease by stabilizing YY1-mediated lipogenic pathways in zebrafish. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2018, 1863, 844–856. [Google Scholar] [CrossRef]
- Zhou, Y.; Lu, M.; Du, R.H.; Qiao, C.; Jiang, C.Y.; Zhang, K.Z.; Ding, J.H.; Hu, G. MicroRNA-7 targets Nod-like receptor protein 3 inflammasome to modulate neuroinflammation in the pathogenesis of Parkinson’s disease. Mol. Neurodegener. 2016, 11, 28. [Google Scholar] [CrossRef]
- Fan, Z.; Lu, M.; Qiao, C.; Zhou, Y.; Ding, J.H.; Hu, G. MicroRNA-7 Enhances Subventricular Zone Neurogenesis by Inhibiting NLRP3/Caspase-1 Axis in Adult Neural Stem Cells. Mol. Neurobiol. 2016, 53, 7057–7069. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.C.; Chae, Y.J.; Kabaria, S.; Chaudhuri, A.D.; Jain, M.R.; Li, H.; Mouradian, M.M.; Junn, E. MicroRNA-7 protects against 1-methyl-4-phenylpyridinium-induced cell death by targeting RelA. J. Neurosci. 2014, 34, 12725–12737. [Google Scholar] [CrossRef]
- Chaudhuri, A.D.; Kabaria, S.; Choi, D.C.; Mouradian, M.M.; Junn, E. MicroRNA-7 Promotes Glycolysis to Protect against 1-Methyl-4-phenylpyridinium-induced Cell Death. J. Biol. Chem. 2015, 290, 12425–12434. [Google Scholar] [CrossRef]
- Kong, B.; Wu, P.C.; Chen, L.; Yang, T.; Yuan, Y.Q.; Kuang, Y.Q.; Cheng, L.; Zhou, H.T.; Gu, J.W. microRNA-7 Protects Against 1-Methyl-4-Phenylpyridinium Iodide-Induced Cell Apoptosis in SH-SY5Y Cells by Directly Targeting Krüpple-Like Factor 4. DNA Cell Biol. 2016, 35, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lv, X.; Zhai, K.; Xu, R.; Zhang, Y.; Zhao, S.; Qin, X.; Yin, L.; Lou, J. MicroRNA-7 inhibits neuronal apoptosis in a cellular Parkinson’s disease model by targeting Bax and Sirt2. Am. J. Transl. Res. 2016, 8, 993–1004. [Google Scholar] [PubMed]
- Salama, R.M.; Abdel-Latif, G.A.; Abbas, S.S.; El Magdoub, H.M.; Schaalan, M.F. Neuroprotective effect of crocin against rotenone-induced Parkinson’s disease in rats: Interplay between PI3K/Akt/mTOR signaling pathway and enhanced expression of miRNA-7 and miRNA-221. Neuropharmacology 2020, 164, 107900. [Google Scholar] [CrossRef] [PubMed]
- Junn, E.; Lee, K.W.; Jeong, B.S.; Chan, T.W.; Im, J.Y.; Mouradian, M.M. Repression of alpha-synuclein expression and toxicity by microRNA-7. Proc. Natl. Acad. Sci. USA 2009, 106, 13052–13057. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Jiang, Y.; Xu, Y.; Li, Y.; Li, B. Identification of miRNA-7 as a regulator of brain-derived neurotrophic factor/α-synuclein axis in atrazine-induced Parkinson’s disease by peripheral blood and brain microRNA profiling. Chemosphere 2019, 233, 542–548. [Google Scholar] [CrossRef] [PubMed]
- McMillan, K.J.; Murray, T.K.; Bengoa-Vergniory, N.; Cordero-Llana, O.; Cooper, J.; Buckley, A.; Wade-Martins, R.; Uney, J.B.; O’Neill, M.J.; Wong, L.F.; et al. Loss of MicroRNA-7 Regulation Leads to α-Synuclein Accumulation and Dopaminergic Neuronal Loss In Vivo. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 2404–2414. [Google Scholar] [CrossRef]
- Zhang, X.D.; Fan, Q.Y.; Qiu, Z.; Chen, S. MiR-7 alleviates secondary inflammatory response of microglia caused by cerebral hemorrhage through inhibiting TLR4 expression. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5597–5604. [Google Scholar] [CrossRef]
- Dong, Y.F.; Chen, Z.Z.; Zhao, Z.; Yang, D.D.; Yan, H.; Ji, J.; Sun, X.L. Potential role of microRNA-7 in the anti-neuroinflammation effects of nicorandil in astrocytes induced by oxygen-glucose deprivation. J. Neuroinflammation 2016, 13, 60. [Google Scholar] [CrossRef]
- Yue, D.; Zhao, J.; Chen, H.; Guo, M.; Chen, C.; Zhou, Y.; Xu, L. MicroRNA-7, synergizes with RORα, negatively controls the pathology of brain tissue inflammation. J. Neuroinflammation 2020, 17, 28. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Dalmasso, G.; Yan, Y.; Laroui, H.; Dahan, S.; Mayer, L.; Sitaraman, S.V.; Merlin, D. MicroRNA-7 modulates CD98 expression during intestinal epithelial cell differentiation. J. Biol. Chem. 2010, 285, 1479–1489. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhang, S.; Chao, K.; Feng, R.; Wang, H.; Li, M.; Chen, B.; He, Y.; Zeng, Z.; Chen, M. E3 Ubiquitin ligase RNF183 Is a Novel Regulator in Inflammatory Bowel Disease. J. Crohn’s Colitis 2016, 10, 713–725. [Google Scholar] [CrossRef]
- Guo, J.; Sun, M.; Teng, X.; Xu, L. MicroRNA-7-5p regulates the expression of TFF3 in inflammatory bowel disease. Mol. Med. Rep. 2017, 16, 1200–1206. [Google Scholar] [CrossRef][Green Version]
- Guo, J.; Xu, L.; Teng, X.; Sun, M. MicroRNA-7-5p regulates the proliferation and migration of intestinal epithelial cells by targeting trefoil factor 3 via inhibiting the phosphoinositide 3-kinase/Akt signalling pathway. Int. J. Mol. Med. 2017, 40, 1435–1443. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, M.; Yan, Y.; Wang, Y.; Zhao, X.; Yang, J.; Chen, J.; Chen, C.; Tang, L.; Zeng, W.; et al. The miR-7/EGFR axis controls the epithelial cell immunomodulation and regeneration and orchestrates the pathology in inflammatory bowel disease. J. Adv. Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.N.; Li, Y.Q.; Liu, Y.; Li, Q.; Hu, Y.; Xu, J.Q.; Sun, T.Y.; Xie, L.X. Exosomes derived from plasma of septic patients inhibit apoptosis of T lymphocytes by down-regulating bad via hsa-miR-7-5p. Biochem. Biophys. Res. Commun. 2019, 513, 958–966. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, F.; Tian, Q.; Sheng, K. CircVMA21 ameliorates lipopolysaccharide (LPS)-induced HK-2 cell injury depending on the regulation of miR-7-5p/PPARA. Autoimmunity 2022, 55, 136–146. [Google Scholar] [CrossRef]
- Wu, X.N.; Ye, Y.X.; Niu, J.W.; Li, Y.; Li, X.; You, X.; Chen, H.; Zhao, L.D.; Zeng, X.F.; Zhang, F.C.; et al. Defective PTEN regulation contributes to B cell hyperresponsiveness in systemic lupus erythematosus. Sci. Transl. Transl. Med. 2014, 6, 246ra99. [Google Scholar] [CrossRef]
- Wang, M.; Chen, H.; Qiu, J.; Yang, H.X.; Zhang, C.Y.; Fei, Y.Y.; Zhao, L.D.; Zhou, J.X.; Wang, L.; Wu, Q.J.; et al. Antagonizing miR-7 suppresses B cell hyperresponsiveness and inhibits lupus development. J. Autoimmun. 2020, 109, 102440. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, L.; Fan, G.; Yang, H.; Wu, L.; Huang, Y.; Xu, N.; Li, J. Role of the ciRS-7/miR-7 axis in the regulation of proliferation, apoptosis and inflammation of chondrocytes induced by IL-1β. Int. Immunopharmacol. 2019, 71, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, J.; Zhou, Y.; Yang, Z.; Yang, H.; Li, D.; Zhang, J.; Zhang, Y.; Xu, N.; Huang, Y.; et al. Down-regulated ciRS-7/up-regulated miR-7 axis aggravated cartilage degradation and autophagy defection by PI3K/AKT/mTOR activation mediated by IL-17A in osteoarthritis. Aging 2020, 12, 20163–20183. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhou, X.; Xu, K.; Hu, P.; Bao, J.; Li, J.; Zhu, J.; Wu, L. miR-7/EGFR/MEGF9 axis regulates cartilage degradation in osteoarthritis via PI3K/AKT/mTOR signaling pathway. Bioengineered 2021, 12, 8622–8634. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guo, H.; Li, L.; Bao, D.; Gao, F.; Li, Q.; Huang, Q.; Duan, X.; Xiang, Z. Long Non-Coding RNA (lncRNA) Small Nucleolar RNA Host Gene 15 (SNHG15) Alleviates Osteoarthritis Progression by Regulation of Extracellular Matrix Homeostasis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e923868. [Google Scholar] [CrossRef]
- Yang, H.; Yang, Z.; Yu, Z.; Xiong, C.; Zhang, Y.; Zhang, J.; Huang, Y.; Zhou, X.; Li, J.; Xu, N. SEMA6D, Negatively Regulated by miR-7, Contributes to C28/I2 chondrocyte’s Catabolic and Anabolic Activities via p38 Signaling Pathway. Oxid. Med. Cell. Longev. 2022, 2022, 9674221. [Google Scholar] [CrossRef]
- Kong, D.; Piao, Y.S.; Yamashita, S.; Oshima, H.; Oguma, K.; Fushida, S.; Fujimura, T.; Minamoto, T.; Seno, H.; Yamada, Y.; et al. Inflammation-induced repression of tumor suppressor miR-7 in gastric tumor cells. Oncogene 2012, 31, 3949–3960. [Google Scholar] [CrossRef]
- Chen, W.S.; Liu, L.C.; Yen, C.J.; Chen, Y.J.; Chen, J.Y.; Ho, C.Y.; Liu, S.H.; Chen, C.C.; Huang, W.C. Nuclear IKKα mediates microRNA-7/-103/107/21 inductions to downregulate maspin expression in response to HBx overexpression. Oncotarget 2016, 7, 56309–56323. [Google Scholar] [CrossRef]
- Chen, Y.J.; Chien, P.H.; Chen, W.S.; Chien, Y.F.; Hsu, Y.Y.; Wang, L.Y.; Chen, J.Y.; Lin, C.W.; Huang, T.C.; Yu, Y.L.; et al. Hepatitis B Virus-Encoded X Protein Downregulates EGFR Expression via Inducing MicroRNA-7 in Hepatocellular Carcinoma Cells. Evid. Based Complement. Altern. Med. Ecam 2013, 2013, 682380. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, C.; Guo, M.; Tao, Y.; Cui, P.; Zhou, Y.; Qin, N.; Zheng, J.; Zhang, J.; Xu, L. MicroRNA-7 Deficiency Ameliorates the Pathologies of Acute Lung Injury through Elevating KLF4. Front. Immunol. 2016, 7, 389. [Google Scholar] [CrossRef]
- Bhushan, R.; Rani, A.; Gupta, D.; Ali, A.; Dubey, P.K. MicroRNA-7 Regulates Insulin Signaling Pathway by Targeting IRS1, IRS2, and RAF1 Genes in Gestational Diabetes Mellitus. MicroRNA 2022, 11, 57–72. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, L.; Du, J.; Hu, X.; Xie, Y.; Wu, J.; Lin, X.; Yin, N.; Sun, M.; Li, H. MicroRNA-7 Inhibits Rotavirus Replication by Targeting Viral NSP5 In Vivo and In Vitro. Viruses 2020, 12, 209. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Q.; Tian, F.L.; Zhang, J.W.; Yang, X.J.; Li, Y.P. Preventive and inhibitive effects of Yiwei Xiaoyu granules on the development and progression of spasmolytic polypeptide-expressing metaplasia lesions. World J. Gastrointest. Oncol. 2021, 13, 1741–1754. [Google Scholar] [CrossRef] [PubMed]
- Ballegaard, V.; Ralfkiaer, U.; Pedersen, K.K.; Hove, M.; Koplev, S.; Brændstrup, P.; Ryder, L.P.; Madsen, H.O.; Gerstoft, J.; Grønbæk, K.; et al. MicroRNA-210, MicroRNA-331, and MicroRNA-7 Are Differentially Regulated in Treated HIV-1-Infected Individuals and Are Associated With Markers of Systemic Inflammation. J. Acquir. Immune Defic. Syndr. 2017, 74, e104–e113. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Guo, M.; Yue, D.; Zhao, J.; Zhou, Y.; Chen, C.; Liang, G.; Xu, L. MicroRNA-7 negatively regulates Toll-like receptor 4 signaling pathway through FAM177A. Immunology 2021, 162, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Renand, A.; Cervera-Marzal, I.; Gil, L.; Dong, C.; Garcia, A.; Kervagoret, E.; Aublé, H.; Habes, S.; Chevalier, C.; Vavasseur, F.; et al. Integrative molecular profiling of autoreactive CD4 T cells in autoimmune hepatitis. J. Hepatol. 2020, 73, 1379–1390. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Arab, J.P.; Reyes, D.; Lapitz, A.; Moshage, H.; Bañales, J.M.; Arrese, M. Extracellular Vesicles in NAFLD/ALD: From Pathobiology to Therapy. Cells 2020, 9, 817. [Google Scholar] [CrossRef] [PubMed]
- Titze-de-Almeida, R.; Titze-de-Almeida, S.S. miR-7 Replacement Therapy in Parkinson’s Disease. Curr. Gene Ther. 2018, 18, 143–153. [Google Scholar] [CrossRef]
- Choi, D.C.; Yoo, M.; Kabaria, S.; Junn, E. MicroRNA-7 facilitates the degradation of alpha-synuclein and its aggregates by promoting autophagy. Neurosci. Lett. 2018, 678, 118–123. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, M.; Yan, R.; Liu, J.; Maddila, S.; Junn, E.; Mouradian, M.M. MicroRNA-7 Protects Against Neurodegeneration Induced by α-Synuclein Preformed Fibrils in the Mouse Brain. Neurotherapeutics 2021, 18, 2529–2540. [Google Scholar] [CrossRef]
- Olsen, T.; Goll, R.; Cui, G.; Christiansen, I.; Florholmen, J. TNF-alpha gene expression in colorectal mucosa as a predictor of remission after induction therapy with infliximab in ulcerative colitis. Cytokine 2009, 46, 222–227. [Google Scholar] [CrossRef]
- Ceccarelli, F.; Perricone, C.; Borgiani, P.; Ciccacci, C.; Rufini, S.; Cipriano, E.; Alessandri, C.; Spinelli, F.R.; Sili Scavalli, A.; Novelli, G.; et al. Genetic Factors in Systemic Lupus Erythematosus: Contribution to Disease Phenotype. J. Immunol. Res. 2015, 2015, 745647. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Taniguchi, K.; Karin, M. NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, K.; Liao, Z.; Li, Y.; Yang, H.; Chen, C.; Zhou, Y.A.; Tao, Y.; Guo, M.; Ren, T.; et al. Promoter mutation of tumor suppressor microRNA-7 is associated with poor prognosis of lung cancer. Mol. Clin. Oncol. 2015, 3, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Seddiki, N.; Brezar, V.; Ruffin, N.; Lévy, Y.; Swaminathan, S. Role of miR-155 in the regulation of lymphocyte immune function and disease. Immunology 2014, 142, 32–38. [Google Scholar] [CrossRef]
- Banerjee, S.; Xie, N.; Cui, H.; Tan, Z.; Yang, S.; Icyuz, M.; Abraham, E.; Liu, G. MicroRNA let-7c regulates macrophage polarization. J. Immunol. 2013, 190, 6542–6549. [Google Scholar] [CrossRef]
- Seok, H.; Lee, H.; Jang, E.S.; Chi, S.W. Evaluation and control of miRNA-like off-target repression for RNA interference. Cell. Mol. Life Sci. CMLS 2018, 75, 797–814. [Google Scholar] [CrossRef]
- Pothoulakis, C.; Torre-Rojas, M.; Duran-Padilla, M.A.; Gevorkian, J.; Zoras, O.; Chrysos, E.; Chalkiadakis, G.; Baritaki, S. CRHR2/Ucn2 signaling is a novel regulator of miR-7/YY1/Fas circuitry contributing to reversal of colorectal cancer cell resistance to Fas-mediated apoptosis. Int. J. Cancer 2018, 142, 334–346. [Google Scholar] [CrossRef]
- Chen, C.; Yue, D.; Lei, L.; Wang, H.; Lu, J.; Zhou, Y.; Liu, S.; Ding, T.; Guo, M.; Xu, L. Promoter-Operating Targeted Expression of Gene Therapy in Cancer: Current Stage and Prospect. Mol. Ther. Nucleic Acids 2018, 11, 508–514. [Google Scholar] [CrossRef]
- Babae, N.; Bourajjaj, M.; Liu, Y.; Van Beijnum, J.R.; Cerisoli, F.; Scaria, P.V.; Verheul, M.; Van Berkel, M.P.; Pieters, E.H.; Van Haastert, R.J.; et al. Systemic miRNA-7 delivery inhibits tumor angiogenesis and growth in murine xenograft glioblastoma. Oncotarget 2014, 5, 6687–6700. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Guo, M.; Zhao, X.; Zhao, J.; Chen, L.; He, Z.; Xu, L.; Zha, Y. MicroRNA-7: A New Intervention Target for Inflammation and Related Diseases. Biomolecules 2023, 13, 1185. https://doi.org/10.3390/biom13081185
Chen C, Guo M, Zhao X, Zhao J, Chen L, He Z, Xu L, Zha Y. MicroRNA-7: A New Intervention Target for Inflammation and Related Diseases. Biomolecules. 2023; 13(8):1185. https://doi.org/10.3390/biom13081185
Chicago/Turabian StyleChen, Chao, Mengmeng Guo, Xu Zhao, Juanjuan Zhao, Longqing Chen, Zhixu He, Lin Xu, and Yan Zha. 2023. "MicroRNA-7: A New Intervention Target for Inflammation and Related Diseases" Biomolecules 13, no. 8: 1185. https://doi.org/10.3390/biom13081185
APA StyleChen, C., Guo, M., Zhao, X., Zhao, J., Chen, L., He, Z., Xu, L., & Zha, Y. (2023). MicroRNA-7: A New Intervention Target for Inflammation and Related Diseases. Biomolecules, 13(8), 1185. https://doi.org/10.3390/biom13081185