
IL-6-Mediated Upregulated miRNAs in Extracellular Vesicles Derived from Lund Human Mesencephalic (LUHMES) Cells: Effects on Astrocytes and Microglia

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Preparation of Nluc-Fused CD81 Plasmids and Their Expression
2.3. Fractionation and Isolation of EVs in Cell Culture Medium Using Nanofiltration
2.4. ELISA Analysis with Anti-CD9, Anti-CD63, and Anti-CD81 Antibodies
2.5. Western Blot Analysis with Immunoprecipitation
2.6. Uptake of EVs and Nluc Activity Assay
2.7. Reverse Transcription and Quantitative Real-Time PCR Analysis (qRT-PCR)
2.8. miRNA Extraction
2.9. miRNA Transfection
2.10. Microarray Analysis
2.11. Statistical Analysis
3. Results
3.1. Purification of EVs Using Nanofiltration
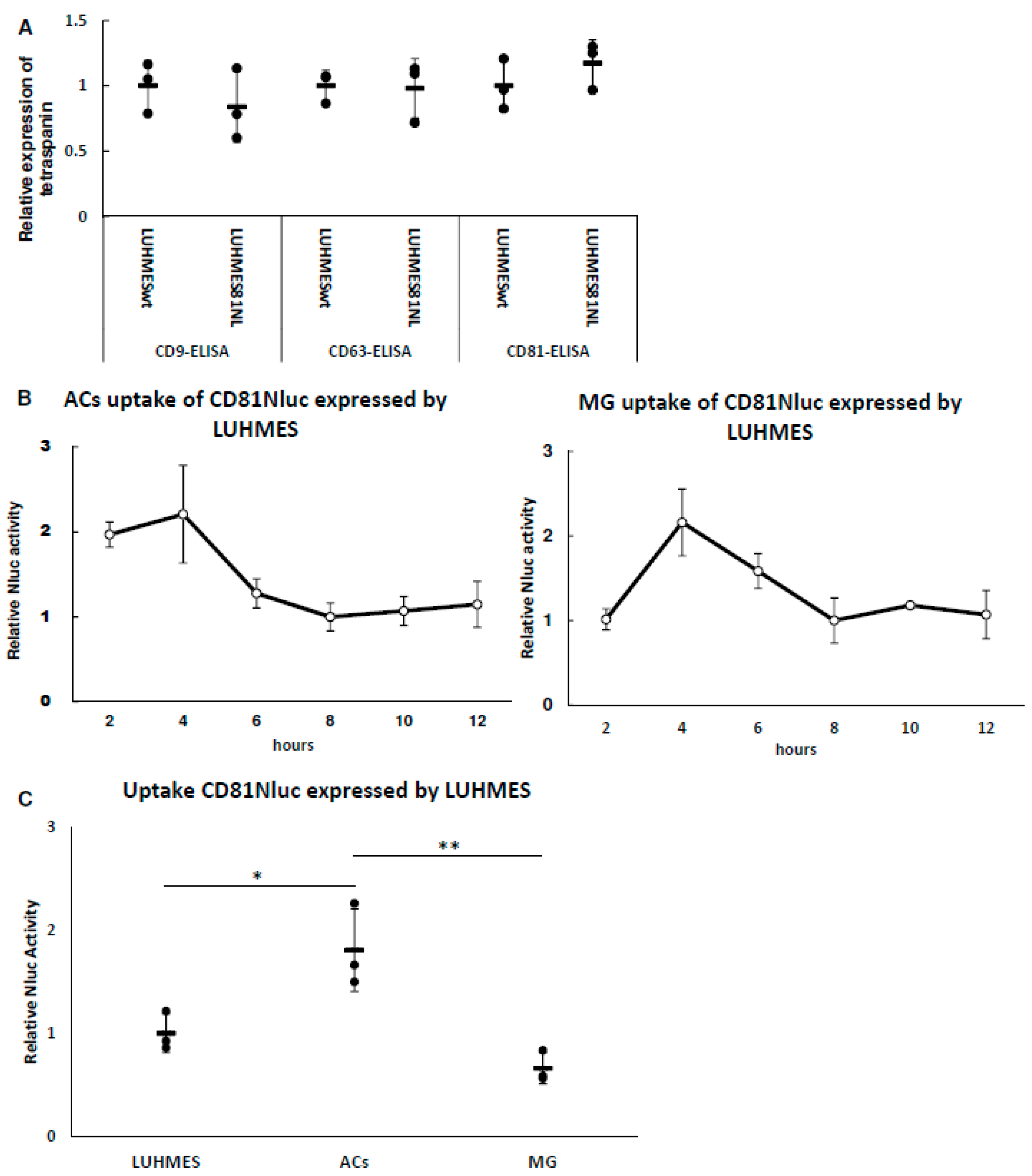
3.2. The Expression of Tetraspanin Proteins and mRNAs
3.3. Uptake of LUHMES-Derived EVs by Astrocytes (ACs) and Microglia (MG)
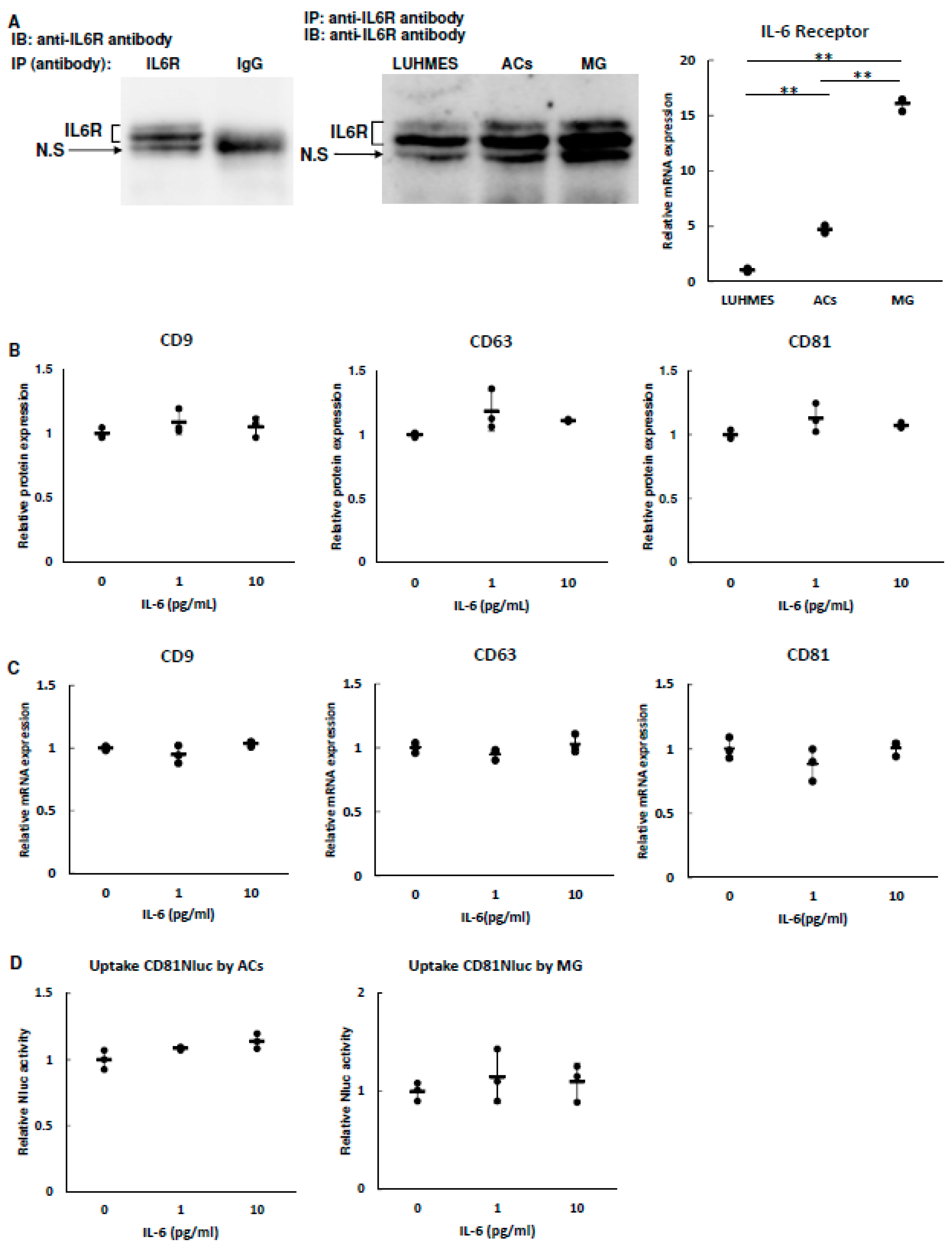
3.4. Effect of IL-6 on EV Secretion and Uptake
3.5. IL-6 Affects the Amounts of miRNAs in EVs
3.6. Target Analysis of Hsa-miR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Wu, X.; Lai, W.; Long, E.; Zhang, X.; Li, W.; Zhu, Y.; Chen, C.; Zhong, X.; Liu, Z.; et al. Prevalence of Depression and Depressive Symptoms among Outpatients: A Systematic Review and Meta-Analysis. BMJ Open 2017, 7, e017173. [Google Scholar] [CrossRef] [PubMed]
- Guilbert, J.J. The World Health Report 2002—Reducing Risks, Promoting Healthy Life. Educ. Health Abingdon Engl. 2003, 16, 230. [Google Scholar] [CrossRef]
- McCarron, R.M.; Shapiro, B.; Rawles, J.; Luo, J. Depression. Ann. Intern. Med. 2021, 174, ITC65–ITC80. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef]
- Ongür, D.; Drevets, W.C.; Price, J.L. Glial Reduction in the Subgenual Prefrontal Cortex in Mood Disorders. Proc. Natl. Acad. Sci. USA 1998, 95, 13290–13295. [Google Scholar] [CrossRef]
- Eroglu, C.; Barres, B.A. Regulation of Synaptic Connectivity by Glia. Nature 2010, 468, 223–231. [Google Scholar] [CrossRef]
- Nagy, C.; Suderman, M.; Yang, J.; Szyf, M.; Mechawar, N.; Ernst, C.; Turecki, G. Astrocytic Abnormalities and Global DNA Methylation Patterns in Depression and Suicide. Mol. Psychiatry 2015, 20, 320–328. [Google Scholar] [CrossRef]
- Nie, X.; Kitaoka, S.; Tanaka, K.; Segi-Nishida, E.; Imoto, Y.; Ogawa, A.; Nakano, F.; Tomohiro, A.; Nakayama, K.; Taniguchi, M.; et al. The Innate Immune Receptors TLR2/4 Mediate Repeated Social Defeat Stress-Induced Social Avoidance through Prefrontal Microglial Activation. Neuron 2018, 99, 464–479.e7. [Google Scholar] [CrossRef]
- Rose-John, S. Interleukin-6 Family Cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028415. [Google Scholar] [CrossRef]
- Qing, H.; Desrouleaux, R.; Israni-Winger, K.; Mineur, Y.S.; Fogelman, N.; Zhang, C.; Rashed, S.; Palm, N.W.; Sinha, R.; Picciotto, M.R.; et al. Origin and Function of Stress-Induced IL-6 in Murine Models. Cell 2020, 182, 372–387.e14. [Google Scholar] [CrossRef]
- Steptoe, A.; Hamer, M.; Chida, Y. The Effects of Acute Psychological Stress on Circulating Inflammatory Factors in Humans: A Review and Meta-Analysis. Brain. Behav. Immun. 2007, 21, 901–912. [Google Scholar] [CrossRef]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef]
- Brietzke, E.; Scheinberg, M.; Lafer, B. Therapeutic Potential of Interleukin-6 Antagonism in Bipolar Disorder. Med. Hypotheses 2011, 76, 21–23. [Google Scholar] [CrossRef]
- Mohr, A.M.; Mott, J.L. Overview of MicroRNA Biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Costa Verdera, H.; Gitz-Francois, J.J.; Schiffelers, R.M.; Vader, P. Cellular Uptake of Extracellular Vesicles Is Mediated by Clathrin-Independent Endocytosis and Macropinocytosis. J. Control. Release 2017, 266, 100–108. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zheng, S.; Luo, Y.; Wang, B. Exosome Theranostics: Biology and Translational Medicine. Theranostics 2018, 8, 237–255. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Schiller, E.A.; Cohen, K.; Lin, X.; El-Khawam, R.; Hanna, N. Extracellular Vesicle-MicroRNAs as Diagnostic Biomarkers in Preterm Neonates. Int. J. Mol. Sci. 2023, 24, 2622. [Google Scholar] [CrossRef]
- Rahimian, N.; Nahand, J.S.; Hamblin, M.R.; Mirzaei, H. Exosomal MicroRNA Profiling. Methods Mol. Biol. Clifton NJ 2023, 2595, 13–47. [Google Scholar] [CrossRef]
- Upadhya, R.; Zingg, W.; Shetty, S.; Shetty, A.K. Astrocyte-Derived Extracellular Vesicles: Neuroreparative Properties and Role in the Pathogenesis of Neurodegenerative Disorders. J. Control. Release Off. J. Control. Release Soc. 2020, 323, 225–239. [Google Scholar] [CrossRef]
- Jotatsu, T.; Izumi, H.; Morimoto, Y.; Yatera, K. Selection of MicroRNAs in Extracellular Vesicles for Diagnosis of Malignant Pleural Mesothelioma by in Vitro Analysis. Oncol. Rep. 2020, 44, 2198–2210. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Kurita, T.; Nishio, K.; Tsukada, J.; Hachisuga, T.; Morimoto, Y.; Iwai, Y.; Izumi, H. Expression of BAF57 in Ovarian Cancer Cells and Drug Sensitivity. Cancer Sci. 2015, 106, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Izumi, H.; Kurita, T.; Koi, C.; Morimoto, Y.; Yoshino, K. UBE2L6 Is Involved in Cisplatin Resistance by Regulating the Transcription of ABCB6. Anti-Cancer Agents Med. Chem. 2020, 20, 1487–1496. [Google Scholar] [CrossRef]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Zhu, L.; Sun, H.-T.; Wang, S.; Huang, S.-L.; Zheng, Y.; Wang, C.-Q.; Hu, B.-Y.; Qin, W.; Zou, T.-T.; Fu, Y.; et al. Isolation and Characterization of Exosomes for Cancer Research. J. Hematol. Oncol. 2020, 13, 152. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Baik, J.-H. Stress and the Dopaminergic Reward System. Exp. Mol. Med. 2020, 52, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Takamura, N.; Nakagawa, S.; Masuda, T.; Boku, S.; Kato, A.; Song, N.; An, Y.; Kitaichi, Y.; Inoue, T.; Koyama, T.; et al. The Effect of Dopamine on Adult Hippocampal Neurogenesis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 50, 116–124. [Google Scholar] [CrossRef]
- Suneson, K.; Lindahl, J.; Chamli Hårsmar, S.; Söderberg, G.; Lindqvist, D. Inflammatory Depression-Mechanisms and Non-Pharmacological Interventions. Int. J. Mol. Sci. 2021, 22, 1640. [Google Scholar] [CrossRef]
- Gupta, D.; Liang, X.; Pavlova, S.; Wiklander, O.P.; Corso, G.; Zhao, Y.; Saher, O.; Bost, J.; Zickler, A.M.; Piffko, A.; et al. Quantification of Extracellular Vesicles in Vitro and in Vivo Using Sensitive Bioluminescence Imaging. J. Extracell. Vesicles 2020, 9, 1800222. [Google Scholar] [CrossRef]
- Mathew, B.; Torres, L.A.; Gamboa Acha, L.; Tran, S.; Liu, A.; Patel, R.; Chennakesavalu, M.; Aneesh, A.; Huang, C.-C.; Feinstein, D.L.; et al. Uptake and Distribution of Administered Bone Marrow Mesenchymal Stem Cell Extracellular Vesicles in Retina. Cells 2021, 10, 730. [Google Scholar] [CrossRef]
- Horibe, S.; Tanahashi, T.; Kawauchi, S.; Murakami, Y.; Rikitake, Y. Mechanism of Recipient Cell-Dependent Differences in Exosome Uptake. BMC Cancer 2018, 18, 47. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A.D.; Dastgheyb, R.M.; Yoo, S.-W.; Trout, A.; Talbot, C.C.; Hao, H.; Witwer, K.W.; Haughey, N.J. TNFα and IL-1β Modify the MiRNA Cargo of Astrocyte Shed Extracellular Vesicles to Regulate Neurotrophic Signaling in Neurons. Cell Death Dis. 2018, 9, 363. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Xu, Q.; Wu, Z.; Chen, Y.; Shu, Y.; Cao, X.; Chen, M.; Zhang, B.; Zhou, Y.; Yao, R.; et al. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Ameliorate IL-6-Induced Acute Liver Injury through MiR-455-3p. Stem Cell Res. Ther. 2020, 11, 37. [Google Scholar] [CrossRef]
- Issler, O.; Haramati, S.; Paul, E.D.; Maeno, H.; Navon, I.; Zwang, R.; Gil, S.; Mayberg, H.S.; Dunlop, B.W.; Menke, A.; et al. MicroRNA 135 Is Essential for Chronic Stress Resiliency, Antidepressant Efficacy, and Intact Serotonergic Activity. Neuron 2014, 83, 344–360. [Google Scholar] [CrossRef] [PubMed]
- Marshe, V.S.; Islam, F.; Maciukiewicz, M.; Fiori, L.M.; Yerko, V.; Yang, J.; Turecki, G.; Foster, J.A.; Kennedy, S.H.; Blumberger, D.M.; et al. Validation Study of MicroRNAs Previously Associated with Antidepressant Response in Older Adults Treated for Late-Life Depression with Venlafaxine. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 100, 109867. [Google Scholar] [CrossRef]
- Fujitani, M.; Yamagishi, S.; Che, Y.H.; Hata, K.; Kubo, T.; Ino, H.; Tohyama, M.; Yamashita, T. P311 Accelerates Nerve Regeneration of the Axotomized Facial Nerve. J. Neurochem. 2004, 91, 737–744. [Google Scholar] [CrossRef]
- Pan, D.; Zhe, X.; Jakkaraju, S.; Taylor, G.A.; Schuger, L. P311 Induces a TGF-Beta1-Independent, Nonfibrogenic Myofibroblast Phenotype. J. Clin. Investig. 2002, 110, 1349–1358. [Google Scholar] [CrossRef]
- Mariani, L.; McDonough, W.S.; Hoelzinger, D.B.; Beaudry, C.; Kaczmarek, E.; Coons, S.W.; Giese, A.; Moghaddam, M.; Seiler, R.W.; Berens, M.E. Identification and Validation of P311 as a Glioblastoma Invasion Gene Using Laser Capture Microdissection. Cancer Res. 2001, 61, 4190–4196. [Google Scholar]
- Seddik, R.; Jungblut, S.P.; Silander, O.K.; Rajalu, M.; Fritzius, T.; Besseyrias, V.; Jacquier, V.; Fakler, B.; Gassmann, M.; Bettler, B. Opposite Effects of KCTD Subunit Domains on GABA(B) Receptor-Mediated Desensitization. J. Biol. Chem. 2012, 287, 39869–39877. [Google Scholar] [CrossRef]
- Miller, G.E.; Chen, E.; Sze, J.; Marin, T.; Arevalo, J.M.G.; Doll, R.; Ma, R.; Cole, S.W. A Functional Genomic Fingerprint of Chronic Stress in Humans: Blunted Glucocorticoid and Increased NF-KappaB Signaling. Biol. Psychiatry 2008, 64, 266–272. [Google Scholar] [CrossRef]
- Wu, T.-N.; Chen, C.-K.; Lee, C.-S.; Wu, B.-J.; Sun, H.-J.; Chang, C.-H.; Chen, C.-Y.; Wu, L.S.-H.; Cheng, A.T.-A. Lithium and GADL1 Regulate Glycogen Synthase Kinase-3 Activity to Modulate KCTD12 Expression. Sci. Rep. 2019, 9, 10255. [Google Scholar] [CrossRef]
- Deng, S.-L.; Hu, Z.-L.; Mao, L.; Gao, B.; Yang, Q.; Wang, F.; Chen, J.-G. The Effects of Kctd12, an Auxiliary Subunit of GABAB Receptor in Dentate Gyrus on Behavioral Response to Chronic Social Defeat Stress in Mice. Pharmacol. Res. 2021, 163, 105355. [Google Scholar] [CrossRef] [PubMed]
- Schwenk, J.; Metz, M.; Zolles, G.; Turecek, R.; Fritzius, T.; Bildl, W.; Tarusawa, E.; Kulik, A.; Unger, A.; Ivankova, K.; et al. Native GABA(B) Receptors Are Heteromultimers with a Family of Auxiliary Subunits. Nature 2010, 465, 231–235. [Google Scholar] [CrossRef]
- Metz, M.; Gassmann, M.; Fakler, B.; Schaeren-Wiemers, N.; Bettler, B. Distribution of the Auxiliary GABAB Receptor Subunits KCTD8, 12, 12b, and 16 in the Mouse Brain. J. Comp. Neurol. 2011, 519, 1435–1454. [Google Scholar] [CrossRef]
- Yu, N.-K.; Kim, H.F.; Shim, J.; Kim, S.; Kim, D.W.; Kwak, C.; Sim, S.-E.; Choi, J.-H.; Ahn, S.; Yoo, J.; et al. A Transducible Nuclear/Nucleolar Protein, MLLP, Regulates Neuronal Morphogenesis and Synaptic Transmission. Sci. Rep. 2016, 6, 22892. [Google Scholar] [CrossRef] [PubMed]
- Castaño, J.; Solanas, G.; Casagolda, D.; Raurell, I.; Villagrasa, P.; Bustelo, X.R.; García de Herreros, A.; Duñach, M. Specific Phosphorylation of P120-Catenin Regulatory Domain Differently Modulates Its Binding to RhoA. Mol. Cell. Biol. 2007, 27, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bassuk, A.G.; Merl-Pham, J.; Hsu, C.-W.; Colgan, D.F.; Li, X.; Au, K.S.; Zhang, L.; Smemo, S.; Justus, S.; et al. Catenin Delta-1 (CTNND1) Phosphorylation Controls the Mesenchymal to Epithelial Transition in Astrocytic Tumors. Hum. Mol. Genet. 2016, 25, 4201–4210. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA IDs | miRNA Expression in EVs | miRNA Expression in Cells | ||||
|---|---|---|---|---|---|---|
| 1 pg/mL IL-6 | 10 pg/mL IL-6 | Ratio * | p-Value ** | ACs | MG | |
| hsa-miR-135a-3p | 120.4 (±25.1) | 284.5 (±39.5) | 2.4 | 0.0495 | 17.4 (±6.4) | 48.0 (±5.2) |
| hsa-miR-11399 | 242.3 (±81.4) | 538.8 (±151.4) | 2.2 | 0.0495 | 15.9 (±5.5) | 32.6 (±3.1) |
| hsa-miR-6790-3p | 176.4 (±89.3) | 383.5 (±29.0) | 2.2 | 0.0495 | 26.5 (±9.3) | 67.3 (±4.6) |
| Cell | Transfected miRNA | Number of mRNAs |
|---|---|---|
| ACs | hsa-miR-135-3p | 2 |
| hsa-miR-6790-3p | 15 | |
| hsa-miR-11399 | 547 | |
| MG | hsa-miR-135-3p | 13 |
| hsa-miR-6790-3p | 34 | |
| hsa-miR-11399 | 205 | |
| Common | hsa-miR-135-3p | 1 |
| hsa-miR-6790-3p | 10 | |
| hsa-miR-11399 | 47 |
| Transfected miRNAs | mRNAs Expression Less than 1/2 | ||
|---|---|---|---|
| hsa-miR-135-3p | UBE2D3 | ||
| hsa-miR-6790-3p | MINOS1-NBL1 | MRPL11 | |
| PTS | RPEL1 | ||
| RAB6A | LLPH | ||
| RPE | IMP4 | ||
| SET | EI24 | ||
| hsa-miR-11399 | ACTR3 | TOR1AIP1 | BCCIP |
| CDKN1A | AMD1 | SERINC1 | |
| CENPF | GATC | SOAT1 | |
| NCKAP1 | ANXA1 | TOP2A | |
| SUPT16H | IPO5 | HIST2H2AA4 | |
| MRFAP1L1 | LIMS1 | CHCHD7 | |
| CREB1 | FAM45A | HIST1H2BG | |
| CTNND1 | ZNF117 | HIST1H2BH | |
| ARL6IP6 | LARS | CASP3 | |
| ZNF836 | ZFR | MARVELD1 | |
| ZNF721 | POLR2B | WDR75DGKZ | |
| EFNB2 | ANLN | USO1PRPF4B | |
| RHOBTB3 | ZFAND6 | HIST3H2A | |
| PUM2 | NDFIP2 | TTC37 | |
| SF3B1 | NOL8 | G3BP2 | |
| BAMBI | EEF1A1P31 | ||
| Gene | hsa-miR-135-3p | hsa-miR-6790-3p | hsa-miR-11399-3p |
|---|---|---|---|
| NREP of ACs | NE | ↓ | ↓ |
| RPEL1 of ACs | NE | ↓ | ↓ |
| CTNND1 of MG | NE | ↓ | ↓ |
| KCTD12 of MG | NE | ↓ | ↓↓ |
| miRNA Name | hsa-miR-135a-3p | hsa-miR-11399 | hsa-miR-6790-3p |
|---|---|---|---|
| miRNA sequence | UAUAGGGAUUGGAGCCGUGGCG | UUCAGGUCUGGGGCUGAAACCU | CGACCUCGGCGACCCCUCACU |
| NREP (Target Scan) | none | none | UGUAUGGGUAUUGAUGAGGUCAU |
| NREP (Emboss water) | none | GAUGCUGCCACAGGACCUGA | GUGUAUGGGUAUUGAUGAGGUC |
| KCTD12 (Target Scan) | none | none | CUCCAAGCGCCGUAGGAGGUCAC |
| KCTD12 (Emboss water) | none | GGUUGCAGCUCCUGAGUGCAGCGCGGCUUCCUG | GCCCGGGGCCGCCGCCCUCG |
| LLPH (Target Scan) | none | none | GAGGCAGGUGGAUCACGAGGUCA |
| LLPH (Emboss water) | none | none | UGAGGCAGAUGGAUCACCUGAGGUC |
| CTNND1 (Target Scan) | none | none | none |
| CTNND1 (Emboss water) | none | GGCUCCGCCCCUUACCUUCA | AGAGAGGAGUGAAGCUCGCCGGAAAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishi, K.; Izumi, H.; Tomonaga, T.; Nagano, C.; Morimoto, Y.; Horie, S. IL-6-Mediated Upregulated miRNAs in Extracellular Vesicles Derived from Lund Human Mesencephalic (LUHMES) Cells: Effects on Astrocytes and Microglia. Biomolecules 2023, 13, 718. https://doi.org/10.3390/biom13050718
Nishi K, Izumi H, Tomonaga T, Nagano C, Morimoto Y, Horie S. IL-6-Mediated Upregulated miRNAs in Extracellular Vesicles Derived from Lund Human Mesencephalic (LUHMES) Cells: Effects on Astrocytes and Microglia. Biomolecules. 2023; 13(5):718. https://doi.org/10.3390/biom13050718
Chicago/Turabian StyleNishi, Kento, Hiroto Izumi, Taisuke Tomonaga, Chikage Nagano, Yasuo Morimoto, and Seichi Horie. 2023. "IL-6-Mediated Upregulated miRNAs in Extracellular Vesicles Derived from Lund Human Mesencephalic (LUHMES) Cells: Effects on Astrocytes and Microglia" Biomolecules 13, no. 5: 718. https://doi.org/10.3390/biom13050718
APA StyleNishi, K., Izumi, H., Tomonaga, T., Nagano, C., Morimoto, Y., & Horie, S. (2023). IL-6-Mediated Upregulated miRNAs in Extracellular Vesicles Derived from Lund Human Mesencephalic (LUHMES) Cells: Effects on Astrocytes and Microglia. Biomolecules, 13(5), 718. https://doi.org/10.3390/biom13050718