
Phytoceramides from the Marine Sponge Monanchora clathrata: Structural Analysis and Cytoprotective Effects

 , and
, and 
Abstract

1. Introduction
2. Materials and Methods
2.1. General Procedures
2.2. Animal Material
2.3. Extraction and Isolation
2.4. Cytotoxic and Cytoprotective Activities
2.4.1. Reagents
2.4.2. Cell Lines and Culture Conditions
2.4.3. Cell Viability Assay for MDA-MB-231 and HL-60 Cells
2.4.4. Paraquat Induced In Vitro Model of Neurotoxicity
2.4.5. Analysis of ROS (Reactive Oxygen Species) Level
2.4.6. Statistical Analysis
3. Results
3.1. Analysis of the Total Ceramide of M. clathrata
3.2. Analysis of the 2-Hydroxy Fatty Acids and Sphingoid Bases Obtained from the Total Ceramide of M. clathrata
3.3. Structure Elucidation of the Phytoceramides of M. clathrata
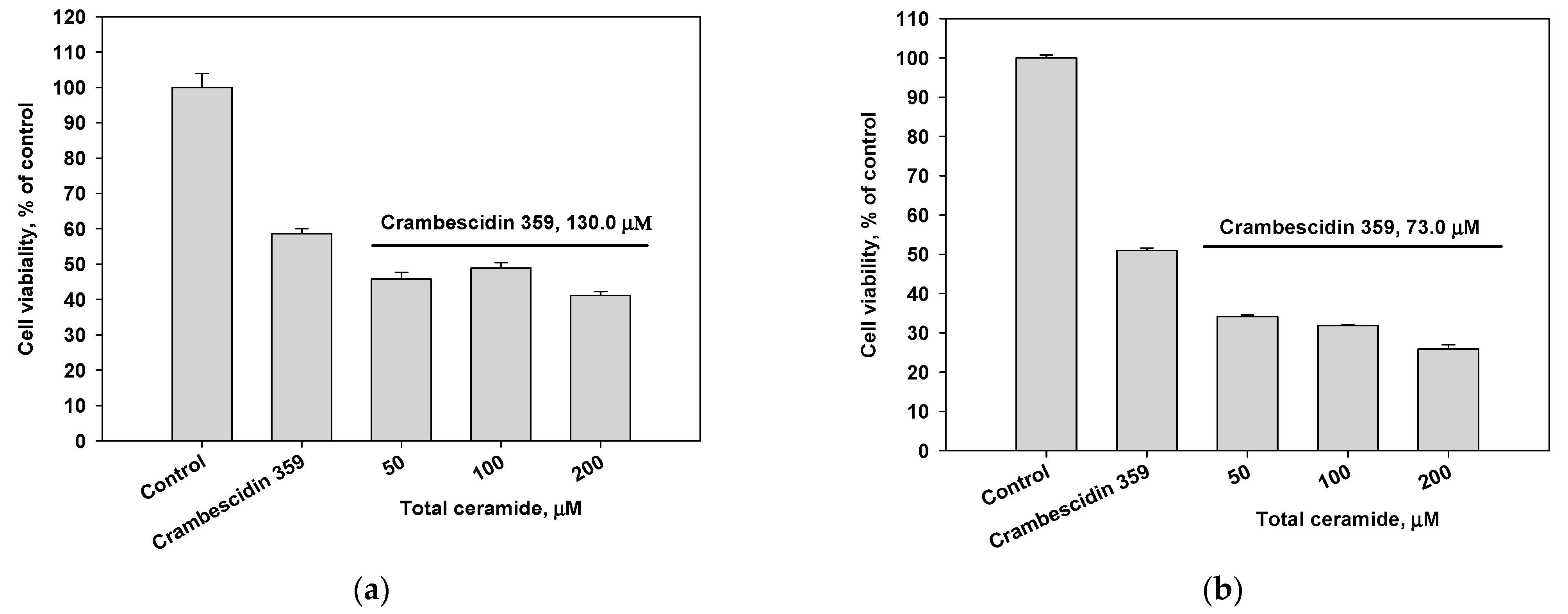
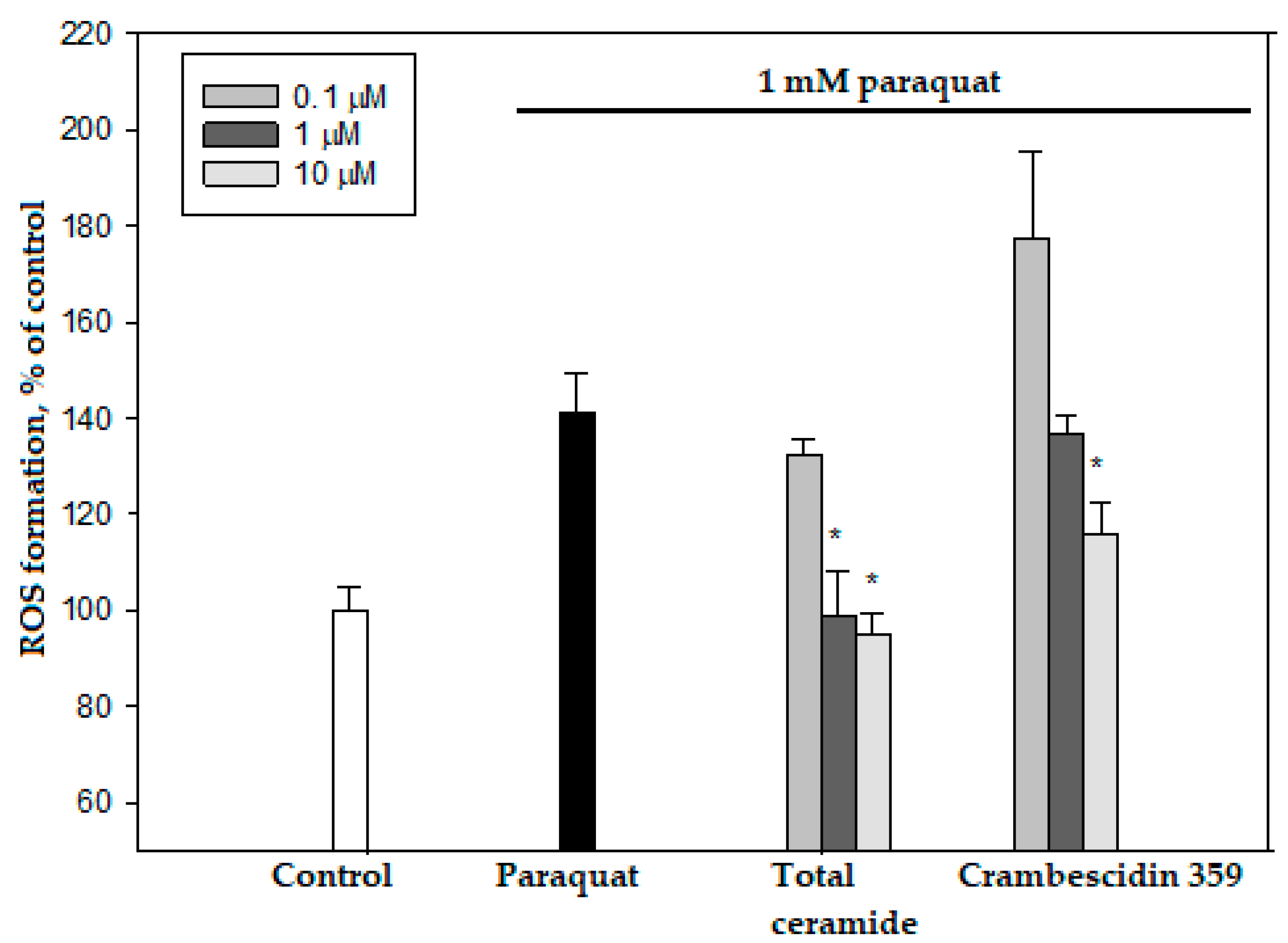
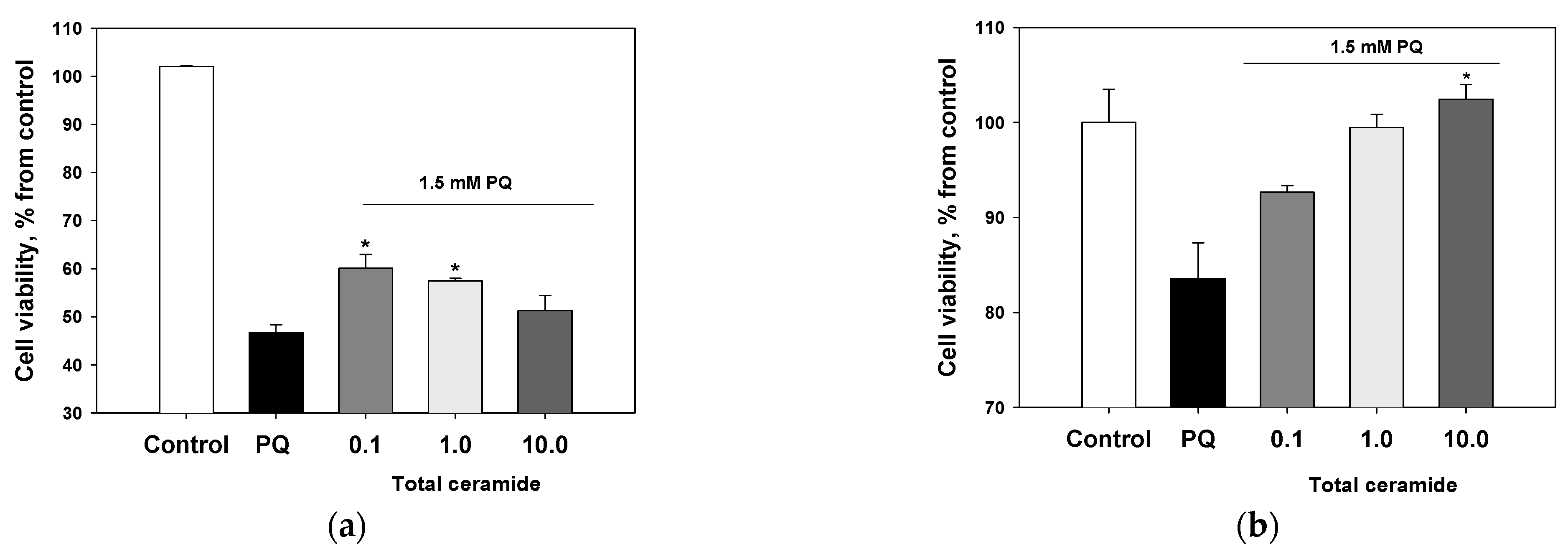
3.4. Cytoprotective and/or Cytotoxic Effects of the Phytoceramides and Crambescidin 359 from M. clathrata
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The LipidWeb. Ceramides. Available online: https://www.lipidmaps.org/resources/lipidweb/index.php?page=lipids/sphingo/ceramide/index.htm (accessed on 1 November 2022).
- Perry, D.K.; Hannun, Y.A. The role of ceramide in cell signaling. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1998, 1436, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Pettus, B.J.; Chalfant, C.E.; Hannun, Y.A. Ceramide in apoptosis: An overview and current perspectives. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2002, 1585, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Beckman, B.S.; Foroozesh, M. A review of ceramide analogs as potential anticancer agents. Future Med. Chem. 2013, 5, 1405–1421. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Tumor suppressive functions of ceramide: Evidence and mechanisms. Apoptosis 2015, 20, 689–711. [Google Scholar] [CrossRef]
- Muralidhar, P.; Radhika, P.; Krishna, N.; Venkata Rao, D.; Bheemasankara Rao, C. Sphingolipids from marine organisms: A review. Nat. Prod. Sci. 2003, 9, 117–142. [Google Scholar]
- Raslan, A.E.; Radwan, M.M.; Ahmed, S.A.; Nafady, A.M.; Zaki, M.A.; Wanas, A.S.; Abou-Karam, M.; Shier, T.W.; Hassanean, H.A.; ElSohly, M.A. Monanchoramides A–D, ceramides from the marine sponge Monanchora clathrata with cytotoxic activity. Phytochem. Lett. 2018, 23, 83–89. [Google Scholar] [CrossRef]
- The LipidWeb. Long-Chain (Sphingoid) Bases. Available online: https://www.lipidmaps.org/resources/lipidweb/index.php?page=lipids/sphingo/lcb/index.htm (accessed on 1 November 2022).
- Jung, J.-C.; Lee, Y.; Moon, S.; Ryu, J.H.; Oh, S. Phytoceramide shows neuroprotection and ameliorates scopolamine-induced memory impairment. Molecules 2011, 16, 9090–9100. [Google Scholar] [CrossRef]
- Jang, J.Y.; Lee, H.K.; Yoo, H.-S.; Seong, Y.H. Phytoceramide ameliorates β-amyloid protein-induced memory impairment and neuronal death in mice. Arch. Pharm. Res. 2017, 40, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Custodia, A.; Aramburu-Núñez, M.; Correa-Paz, C.; Posado-Fernández, A.; Gómez-Larrauri, A.; Castillo, J.; Gómez-Muñoz, A.; Sobrino, T.; Ouro, A. Ceramide Metabolism and Parkinson’s Disease—Therapeutic Targets. Biomolecules 2021, 11, 945. [Google Scholar] [CrossRef] [PubMed]
- Santalova, E.A. Structural Analysis of Sterols and Cytotoxic Compounds from Some Marine Sponges. Ph.D. Thesis, Pacific Institute of Bioorganic Chemistry, Vladivostok, Russia, 2005. [Google Scholar]
- Aveldaño, M.I.; Horrocks, L.A. Quantitative release of fatty acids from lipids by a simple hydrolysis procedure. J. Lipid Res. 1983, 24, 1101–1105. [Google Scholar] [CrossRef]
- Santalova, E.A.; Denisenko, V.A.; Dmitrenok, P.S.; Drozdov, A.L.; Stonik, V.A. Cerebrosides from a Far-Eastern glass sponge Aulosaccus sp. Lipids 2015, 50, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Santalova, E.A.; Svetashev, V.I. Preparation of 4,4-dimethyloxazoline and pyrrolidine derivatives from fatty acid methyl esters using sodium borohydride: Mild and simple one-pot derivatization procedures for a gas chromatographic–mass spectrometric analysis of fatty acids. Nat. Prod. Commun. 2022, 17, 1934578X221131408. [Google Scholar] [CrossRef]
- AOCS Lipid Library. Preparation of Ester Derivatives of Fatty Acids for Chromatographic Analysis. Available online: https://lipidlibrary.aocs.org/lipid-analysis/selected-topics-in-the-analysis-of-lipids/preparation-of-ester-derivatives-of-fatty-acids-for-chromatographic-analysis (accessed on 12 November 2022).
- The LipidWeb. Mass Spectrometry of Methyl Esters. Hydroxy Fatty Acids (Not Further Derivatized). Available online: https://www.lipidmaps.org/resources/lipidweb/index.php?page=ms/methesters/me-hydroxy-1/index.htm (accessed on 20 November 2022).
- Horn, D.H.S.; Pretorius, Y.Y. Wool wax. Part VI. The synthesis and stereochemistry of the straight-chain α-hydroxy-acids. J. Chem. Soc. 1954, 1954, 1460–1464. [Google Scholar] [CrossRef]
- Koike, K.; Sugimoto, M.; Nakahara, Y.; Ogawa, T. Total synthesis of cerebrosides: (2S,3R,4E)-1-O-β-D-galactopyranosyl-N-(2′R and 2′S)-2′-hydroxytetracosanoylsphingenine. Carbohydr. Res. 1987, 162, 237–246. [Google Scholar] [CrossRef]
- Sugiyama, S.; Honda, M.; Komori, T. Biologically active glycosides from Asteroidea. XV. Asymmetric synthesis of phytosphingosine and phytosphingosine anhydro base: Assignment of the absolute stereochemistry. Liebigs Ann. Chem. 1988, 1988, 619–625. [Google Scholar] [CrossRef]
- Santalova, E.A.; Denisenko, V.A.; Dmitrenok, P.S.; Ha, D.T.; Hung, N.A.; Drozdov, A.L.; Malyarenko, O.S.; Thuy, T.T.T.; Quan, P.M.; Long, P.Q. Occurrence of melibiose-containing glycosphingolipids in a sample of a sponge-coral association (Desmapsamma anchorata/Carijoa riisei). Chem. Biodivers. 2019, 16, e1800401. [Google Scholar] [CrossRef]
- Hsu, F.-F. Complete structural characterization of ceramides as [M–H]– ions by multiple-stage linear ion trap mass spectrometry. Biochimie 2016, 130, 63–75. [Google Scholar] [CrossRef]
- Kudryavtsev, D.; Makarieva, T.; Utkina, N.; Santalova, E.; Kryukova, E.; Methfessel, C.; Tsetlin, V.; Stonik, V.; Kasheverov, I. Marine Natural Products Acting on the Acetylcholine-Binding Protein and Nicotinic Receptors: From Computer Modeling to Binding Studies and Electrophysiology. Mar. Drugs 2014, 12, 1859–1875. [Google Scholar] [CrossRef]
- Makotrova, T.A. A role of alpha-7 nicotinic acetylcholine receptors in pharmacotherapy of neurodegenerative diseases. Zhurnal Nevrol. Psikhiatrii imeni SS Korsakova 2012, 112, 57–59. [Google Scholar]
- Database SciFinder. Available online: https://scifinder-n.cas.org (accessed on 17 January 2023).
- Hirsch, S.; Kashman, Y. New glycosphigolipids from marine organisms. Tetrahedron 1989, 45, 3897–3906. [Google Scholar] [CrossRef]
- Berlinck, R.G.S. Chromatographic approach to polar compounds: Isolation of hydrophilic constituents of the marine sponge Crambe crambe. Quim. Nova 1994, 17, 167–171. [Google Scholar]
- Xu, S.-H.; Yang, K. Three new ceramides from the sponge Spongia suriganensis. Chin. J. Org. Chem. 2006, 26, 56–59. [Google Scholar]
- Lakshmi, V.; Raghubir, R.; Gupta, P. New ceramides from the sponge Cinachyra cavernosa. J. Asian Nat. Prod. Res. 2008, 10, 747–751. [Google Scholar] [CrossRef]
- Xu, S.; Liao, X.; Li, H. Compounds with nitrogen from the sponge Hyatella sp. Tianran Chanwu Yanjiu Yu Kaifa 2009, 21, 63–65. [Google Scholar]
- Abdelhameed, R.; Elgawish, M.S.; Mira, A.; Ibrahim, A.K.; Ahmed, S.A.; Shimizu, K.; Yamada, K. Anti-choline esterase activity of ceramides from the Red Sea marine sponge Mycale euplectellioides. RSC Adv. 2016, 6, 20422–20430. [Google Scholar] [CrossRef]
- Inagaki, M.; Ikeda, Y.; Kawatake, S.; Nakamura, K.; Tanaka, M.; Misawa, E.; Yamada, M.; Higuchi, R. Isolation and structure of four new ceramides from the starfish Luidia maculata. Chem. Pharm. Bull. 2006, 54, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Abdelhameed, R.F.; Ibrahim, A.K.; Yamada, K.; Ahmed, S.A. Cytotoxic and anti-inflammatory compounds from Red Sea grass Thalassodendron ciliatum. Med. Chem. Res. 2018, 27, 1238–1244. [Google Scholar] [CrossRef]
- Lawson, M.P.; Bergquist, P.R.; Cambie, R.C. Fatty acid composition and classification of the Porifera. Biochem. Syst. Ecol. 1984, 12, 375–393. [Google Scholar] [CrossRef]
- Shi, Y.; Moazami, Y.; Pierce, J.G. Structure, synthesis and biological properties of the pentacyclic guanidinium alkaloids. Bioorg. Med. Chem. 2017, 25, 2817–2824. [Google Scholar] [CrossRef]
- Raslan, A.E.; Radwan, M.M.; Ahmed, S.A.; Nafady, A.M.; Metwaly, A.M.; Eissa, I.H.; Jacob, M.R.; Hassanean, H.A.; ElSohly, M.A.; Wanas, A.S. Antifungal guanidine alkaloids from the marine sponges Monanchora clathrata and Monanchora unguiculata. Phytochem. Lett. 2022, 51, 132–139. [Google Scholar] [CrossRef]
- Bove, J.; Prou, D.; Perier, C.; Przedborski, S. Toxin-induced models of Parkinson’s disease. NeuroRX 2005, 2, 484–494. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH (mult., J Hz) | δC |
|---|---|---|
| Sphingoid base moieties | ||
| 1 | 4.44 (dd, 5.0, 10.8), 4.53 (dd, 4.5, 10.8) | 61.9 |
| 2 | 5.12 (m) | 52.9 |
| 3 | 4.37 (dd, 5.0, 6.3) | 76.7 |
| 4 | 4.30 (m) | 72.9 |
| 5 | 1.95 (m), 2.26 (m) | 34.0 |
| 6 | 1.72 (m), 1.95 (m) | 26.4 |
| NH | 8.58 (d, 9.0) | – |
| N-Acyl moieties | ||
| 1/ | – | 175.1 |
| 2/ | 4.635 (dd, 3.8, 7.8) | 72.3 |
| 3/ | 2.06 (m), 2.25 (m) | 35.5 |
| 4/ | 1.77 (m) | 25.6 |
| General features of hydrocarbon chains ** | ||
| CH2-pool | 1.20–1.50, m | 28.0–30.1 |
| (n–2) CH2 of normal-chain form | 1.24 (m) | 31.9 |
| (n–1) CH2 of normal-chain form | 1.245 (m) | 22.7 |
| CH3 of normal-chain form | 0.88 (t, 7.0) | 14.0 |
| (n–2) CH2 of iso-form | 1.16 (m) | 39.1 |
| (n–1) CH of iso-form | 1.50 (sep, 6.6) | 27.5 |
| CH3 of iso-form | 0.88 (d, 6.6) | 22.5 |
| Molecular Formulae | [M − H]− | Sphingoid Base/2-Hydroxy Fatty Acid * |
|---|---|---|
| C39H79NO5 | 640.5886 | C17/C22, C18/C21 |
| C40H81NO5 | 654.6045 | C18/C22 |
| C41H83NO5 | 668.6196 | C17/C24, C18/C23, C19/C22 |
| C42H85NO5 | 682.6354 | C17/C25, C18/C24, C19/C23 |
| C43H87NO5 | 696.6510 | C18/C25, C19/C24 |
| C44H89NO5 | 710.6665 | C18/C26, C19/C25 |
| C45H91NO5 | 724.6828 | C19/C26 |
| Fatty Acid | Retention Time (min) | % | GC-MS, m/z |
|---|---|---|---|
| (2R)-2-OH-21:0 | 18.47 | 4.2 | 356 [M]+, 297 [M − 59]+, 90 |
| (2R)-2-OH-22:0 | 19.24 | 39.0 | 370 [M]+, 311 [M − 59]+, 90 |
| (2R)-2-OH-i-23:0 | 19.72 | 1.9 | 384 [M]+, 325 [M − 59]+, 90 |
| (2R)-2-OH-23:0 | 20.02 | 5.4 | 384 [M]+, 325 [M − 59]+, 90 |
| (2R)-2-OH-24:0 | 20.95 | 28.2 | 398 [M]+, 339 [M − 59]+, 90 |
| (2R)-2-OH-25:0 | 22.02 | 17.8 | 412 [M]+, 353 [M − 59]+, 90 |
| (2R)-2-OH-26:0 | 23.28 | 3.5 | 426 [M]+, 367 [M − 59]+, 90 |
| Sphingoid Base | RRT ** | % in Fraction *** | GC-MS, m/z (Relative Intensity, %) |
|---|---|---|---|
| i-t17:0 | 0.94 | tr. | 338 [M − AcOCH2 − AcOH]+ (11), 296 [M − AcOCH2 − AcOH − CH2CO]+ (10), 291 [M − 3AcOH]+ (9), 278 [M − AcOCH2 − 2AcOH]+ (15), 144 [AcOCH2CH(NHAc)]+ (68), 102 [AcOCH2CH(NHAc) − CH2CO]+ (45), 84 [AcOCH2CH(NHAc) − AcOH]+ (100) |
| n-t17:0 | 0.96 | 2.6 | 410/412 [M − AcOH − H/M − AcNH2]+ (0.5/0.4), 338 [M − AcOCH2 − AcOH]+ (10), 296 [M − AcOCH2 − AcOH − CH2CO]+ (12), 291 [M − 3AcOH]+ (12), 278 [M − AcOCH2 − 2AcOH]+ (16), 144 [AcOCH2CH(NHAc)]+ (78), 102 [AcOCH2CH(NHAc) − CH2CO]+ (48), 84 [AcOCH2CH(NHAc) − AcOH]+ (100) |
| i-t18:0 | 0.98 | 80.2 | 424/425/426 [M − AcOH − H/M − AcOH/M − AcNH2]+ (0.6/0.3/0.6), 352 [M − AcOCH2 − AcOH]+ (11), 310 [M − AcOCH2 − AcOH − CH2CO]+ (9), 305 [M − 3AcOH]+ (10), 292 [M − AcOCH2 − 2AcOH]+ (18), 144 [AcOCH2CH(NHAc)]+ (82), 102 [AcOCH2CH(NHAc) − CH2CO]+ (46), 84 [AcOCH2CH(NHAc) − AcOH]+ (100) |
| n-t18:0 | 1.00 | 1.5 | 424/425/426 [M − AcOH − H/M − AcOH/M − AcNH2]+ (0.6/0.2/0.4), 352 [M − AcOCH2 − AcOH]+ (10), 310 [M − AcOCH2 − AcOH − CH2CO]+ (12), 305 [M − 3AcOH]+ (12), 292 [M − AcOCH2 − 2AcOH]+ (16), 144 [AcOCH2CH(NHAc)]+ (79), 102 [AcOCH2CH(NHAc) − CH2CO]+ (46), 84 [AcOCH2CH(NHAc) − AcOH]+ (100) |
| i-t19:0 | 1.03 | 3.3 | 438/440 [M − AcOH − H/M − AcNH2]+ (0.6/0.2), 366 [M − AcOCH2 − AcOH]+ (11), 324 [M − AcOCH2 − AcOH − CH2CO]+ (12), 319 [M − 3AcOH]+ (10), 306 [M − AcOCH2 − 2AcOH]+ (17), 144 [AcOCH2CH(NHAc)]+ (81), 102 [AcOCH2CH(NHAc) − CH2CO]+ (43), 84 [AcOCH2CH(NHAc) − AcOH]+ (100) |
| ai-t19:0 | 1.04 | 12.2 | 438/439/440 [M − AcOH − H/M − AcOH/M − AcNH2]+ (0.5/0.2/0.6), 366 [M − AcOCH2 − AcOH]+ (10), 324 [M − AcOCH2 − AcOH − CH2CO]+ (11), 319 [M − 3AcOH]+ (10), 306 [M − AcOCH2 − 2AcOH]+ (18), 144 [AcOCH2CH(NHAc)]+ (82), 102 [AcOCH2CH(NHAc) − CH2CO]+ (43), 84 [AcOCH2CH(NHAc) − AcOH]+ (100) |
| RP-HPLC Fraction | Ceramide | % in Fraction * | % in Total Ceramide * | Composition: Sphingoid Base/2-Hydroxy Acid |
|---|---|---|---|---|
| I | 1b | 3.3 | 0.2 | i-t17:0/(2R)-2-OH-22:0 |
| 2b | 30.7 | 1.4 | n-t17:0/(2R)-2-OH-22:0 | |
| 3a | 61.3 | 2.9 | i-t18:0/(2R)-2-OH-21:0 | |
| 4a | 4.7 | tr. | n-t18:0/(2R)-2-OH-21:0 | |
| II | 3b | 98.1 | 26.7 | i-t18:0/(2R)-2-OH-22:0 |
| 4b | 1.9 | 0.5 | n-t18:0/(2R)-2-OH-22:0 | |
| III | 2e | 8.6 | 1.1 | n-t17:0/(2R)-2-OH-24:0 |
| 3c | 20.6 | 2.8 | i-t18:0/(2R)-2-OH-23:0 | |
| 3d | 11.8 | 1.5 | i-t18:0/(2R)-2-OH-i-23:0 | |
| 4d | 0.9 | tr. | n-t18:0/(2R)-2-OH-23:0 | |
| 5b | 20.7 | 1.5 | i-t19:0/(2R)-2-OH-22:0 | |
| 6b | 36.4 | 5.6 | ai-t19:0/(2R)-2-OH-22:0 | |
| IV | 2f | 17.9 | 0.7 | n-t17:0/(2R)-2-OH-25:0 |
| 3e | 73.3 | 20.0 | i-t18:0/(2R)-2-OH-24:0 | |
| 4e | 1.5 | tr. | n-t18:0/(2R)-2-OH-24:0 | |
| 5c | 1.3 | tr. | i-t19:0/(2R)-2-OH-23:0 | |
| 5d | 0.5 | tr. | i-t19:0/(2R)-2-OH-i-23:0 | |
| 6c | 3.9 | 1.0 | ai-t19:0/(2R)-2-OH-23:0 | |
| 6d | 1.4 | tr. | ai-t19:0/(2R)-2-OH-i-23:0 | |
| V | 3f | 67.6 | 14.5 | i-t18:0/(2R)-2-OH-25:0 |
| 4f | 1.4 | 0.3 | n-t18:0/(2R)-2-OH-25:0 | |
| 5e | 8.1 | 1.6 | i-t19:0/(2R)-2-OH-24:0 | |
| 6e | 22.9 | 5.8 | ai-t19:0/(2R)-2-OH-24:0 | |
| VI | 3g | 44.7 | 3.4 | i-t18:0/(2R)-2-OH-26:0 |
| 5f | 11.8 | 0.9 | i-t19:0/(2R)-2-OH-25:0 | |
| 6f | 43.4 | 3.3 | ai-t19:0/(2R)-2-OH-25:0 | |
| VII | 5g | 18.2 | 0.2 | i-t19:0/(2R)-2-OH-26:0 |
| 6g | 81.8 | 0.9 | ai-t19:0/(2R)-2-OH-26:0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santalova, E.A.; Kuzmich, A.S.; Chingizova, E.A.; Menchinskaya, E.S.; Pislyagin, E.A.; Dmitrenok, P.S. Phytoceramides from the Marine Sponge Monanchora clathrata: Structural Analysis and Cytoprotective Effects. Biomolecules 2023, 13, 677. https://doi.org/10.3390/biom13040677
Santalova EA, Kuzmich AS, Chingizova EA, Menchinskaya ES, Pislyagin EA, Dmitrenok PS. Phytoceramides from the Marine Sponge Monanchora clathrata: Structural Analysis and Cytoprotective Effects. Biomolecules. 2023; 13(4):677. https://doi.org/10.3390/biom13040677
Chicago/Turabian StyleSantalova, Elena A., Alexandra S. Kuzmich, Ekaterina A. Chingizova, Ekaterina S. Menchinskaya, Evgeny A. Pislyagin, and Pavel S. Dmitrenok. 2023. "Phytoceramides from the Marine Sponge Monanchora clathrata: Structural Analysis and Cytoprotective Effects" Biomolecules 13, no. 4: 677. https://doi.org/10.3390/biom13040677
APA StyleSantalova, E. A., Kuzmich, A. S., Chingizova, E. A., Menchinskaya, E. S., Pislyagin, E. A., & Dmitrenok, P. S. (2023). Phytoceramides from the Marine Sponge Monanchora clathrata: Structural Analysis and Cytoprotective Effects. Biomolecules, 13(4), 677. https://doi.org/10.3390/biom13040677