
Programming DNA Reaction Networks Using Allosteric DNA Hairpins


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Chemical Reagents
2.2. DNA Assembly
2.3. Native PAGE
2.4. Fluorescence Spectroscopy
3. Results
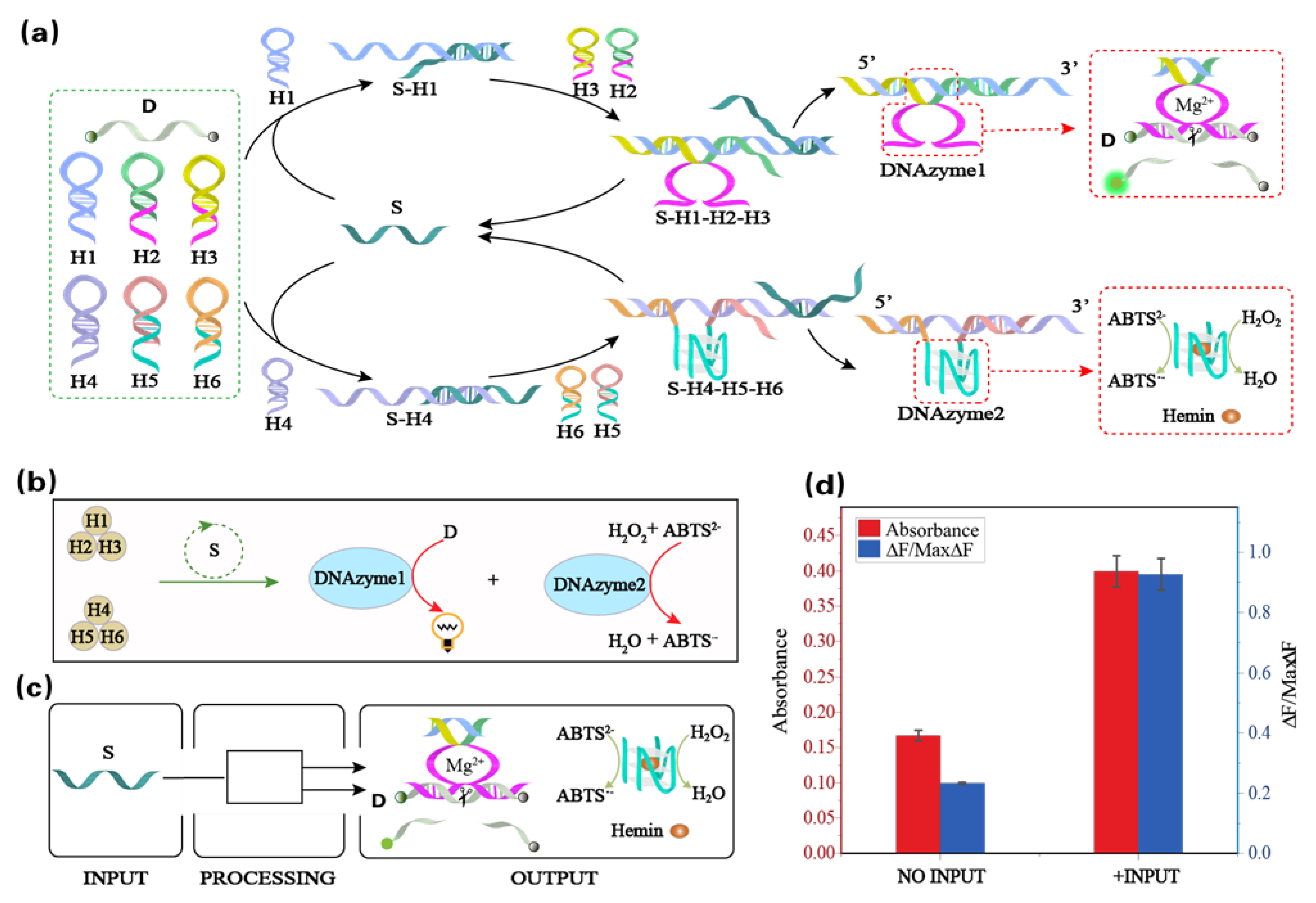
3.1. The Principle and Verification of the Mg2+-Ion-Dependent DNAzyme Dynamic Assembly Module
3.2. The Principle and Verification of the Hemin/G-quadruplex DNAzyme Dynamic Assembly Module
3.3. The Principle and Verification of the Single-Input Dual-Output Module
3.4. The Principle and Verification of the Reversible Regulatory DNA Reaction Network
3.4.1. The Principle of the Reversible Regulatory DNA Reaction Network
3.4.2. Verification of the Reversible Regulatory DNA Reaction Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barabasi, A.L.; Oltvai, Z.N. Network biology: Understanding the cell′s functional organization. Nat. Rev. Genet. 2004, 5, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Purvis, J.E.; Lahav, G. Encoding and Decoding Cellular Information through Signaling Dynamics. Cell 2013, 152, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.Q.; Wang, Y.; Yan, J.H.; Wen, N.C.; Xiong, H.J.; Cai, S.D.; He, Q.Y.; Peng, D.M.; Liu, Z.B.; Liu, Y.F. Dynamic DNA Assemblies in Biomedical Applications. Adv. Sci. 2020, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Tikhomirov, G.; Petersen, P.; Qian, L.L. Fractal assembly of micrometre-scale DNA origami arrays with arbitrary patterns. Nature 2017, 552, 67–71. [Google Scholar] [CrossRef]
- Evans, C.G.; Winfree, E. Physical principles for DNA tile self-assembly. Chem. Soc. Rev. 2017, 46, 3808–3829. [Google Scholar] [CrossRef]
- Wang, R.X.; Ji, C.J.; Zhang, Y.X.; Li, Y. Focus, Fusion, and Rectify: Context-Aware Learning for COVID-19 Lung Infection Segmentation. IEEE Trans. Neural Netw. Learn. Syst. 2022, 33, 12–24. [Google Scholar] [CrossRef]
- Gong, X.; Li, R.M.; Wang, J.; Wei, J.; Ma, K.; Liu, X.Q.; Wang, F. A Smart Theranostic Nanocapsule for Spatiotemporally Programmable Photo-Gene therapy. Angew. Chem. Int. Edit. 2020, 59, 21648–21655. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Han, P.; Chen, W.; Gao, C.; Wang, S.; Song, T.; Niu, M.; Rodriguez-Paton, A. MARPPI: Boosting prediction of protein-protein interactions with multi-scale architecture residual network. Brief. Bioinform. 2022, 24, bbac524. [Google Scholar] [CrossRef]
- Zou, C.Y.; Zhang, Q.; Zhou, C.J.; Cao, W.Y. A nonlinear neural network based on an analog DNA toehold mediated strand displacement reaction circuit. Nanoscale 2022, 14, 6585–6599. [Google Scholar] [CrossRef]
- Xiong, X.W.; Zhu, T.; Zhu, Y.; Cao, M.Y.; Xiao, J.; Li, L.; Wang, F.; Fan, C.H.; Pei, H. Molecular convolutional neural networks with DNA regulatory circuits. Nat. Mach. Intell. 2022, 4, 625–635. [Google Scholar] [CrossRef]
- Topal, M.D.; Fresco, J.R. Complementary base pairing and the origin of substitution mutations. Nature 1976, 263, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Zheng, Y.; Wang, B.; Zhang, Q. Design of Constraint Coding Sets for Archive DNA Storage. IEEE/ACM Trans. Comput. Biol. Bioinform. 2021, 19, 3384–3394. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Huang, Z.; Liao, S.; Zhou, C.; Yan, S.; Chen, G. Improved Bare Bones Particle Swarm Optimization for DNA Sequence Design. IEEE Trans. Nanobioscience 2022, 1. [Google Scholar] [CrossRef]
- Cao, B.; Shi, P.; Zheng, Y.; Zhang, Q. FMG: An observable DNA storage coding method based on frequency matrix game graphs. Comput. Biol. Med. 2022, 151, 106269. [Google Scholar] [CrossRef]
- Lv, Y.F.; Hu, R.; Zhu, G.Z.; Zhang, X.B.; Mei, L.; Liu, Q.L.; Qiu, L.P.; Wu, C.C.; Tan, W.H. Preparation and biomedical applications of programmable and multifunctional DNA nanoflowers. Nat. Protoc. 2015, 10, 1508–1524. [Google Scholar] [CrossRef]
- Lyu, Y.F.; Wu, C.C.; Heinke, C.; Han, D.; Cai, R.; Teng, I.T.; Liu, Y.; Liu, H.; Zhang, X.B.; Liu, Q.L.; et al. Constructing Smart Protocells with Built-In DNA Computational Core to Eliminate Exogenous Challenge. J. Am. Chem. Soc. 2018, 140, 6912–6920. [Google Scholar] [CrossRef]
- Amodio, A.; Del Grosso, E.; Troina, A.; Placidi, E.; Ricci, F. Remote Electronic Control of DNA-Based Reactions and Nanostructure Assembly. Nano Lett. 2018, 18, 2918–2923. [Google Scholar] [CrossRef]
- Simmel, F.C.; Yurke, B.; Singh, H.R. Principles and Applications of Nucleic Acid Strand Displacement Reactions. Chem. Rev. 2019, 119, 6326–6369. [Google Scholar] [CrossRef]
- Lu, C.H.; Qi, X.J.; Orbach, R.; Yang, H.H.; Mironi-Harpaz, I.; Seliktar, D.; Willner, I. Switchable Catalytic Acrylamide Hydrogels Cross-Linked by Hemin/G-Quadruplexes. Nano Lett. 2013, 13, 1298–1302. [Google Scholar] [CrossRef]
- Miyake, Y.; Togashi, H.; Tashiro, M.; Yamaguchi, H.; Oda, S.; Kudo, M.; Tanaka, Y.; Kondo, Y.; Sawa, R.; Fujimoto, T.; et al. Mercury(II)-mediated formation of thymine-Hg-II-thymine base pairs in DNA duplexes. J. Am. Chem. Soc. 2006, 128, 2172–2173. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; O’Hagan, M.P.; Li, Z.Y.; Zhang, J.J.; Ma, X.; Tian, H.; Willner, I. Photoresponsive DNA materials and their applications. Chem. Soc. Rev. 2022, 51, 720–760. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, Y.; Asanuma, H. Light-Driven DNA Nanomachine with a Photoresponsive Molecular Engine. Accounts Chem. Res. 2014, 47, 1663–1672. [Google Scholar] [CrossRef]
- Gehring, K.; Leroy, J.L.; Gueron, M. A tetrameric DNA structure with protonated cytosine.cytosine base pairs. Nature 1993, 363, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.H.; Zhou, K.; Liu, L.; Wu, H.C. Construction of a pH-Mediated Single-Molecule Switch with a Nanopore-DNA Complex. Small 2022, 18, 8. [Google Scholar] [CrossRef]
- Sun, L.J.; Cao, B.; Liu, Y.; Shi, P.J.; Zheng, Y.F.; Wang, B.; Zhang, Q. TripDesign: A DNA Triplex Design Approach Based on Interaction Forces. J. Phys. Chem. B 2022, 126, 8708–8719. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.K.; Zhang, Q.; Liu, Y.; Wang, B.; Zhou, S.H. A molecular device: A DNA molecular lock driven by the nicking enzymes. Comp. Struct. Biotechnol. J. 2020, 18, 2107–2116. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Turberfield, A.J.; Yurke, B.; Winfree, E. Engineering entropy-driven reactions and networks catalyzed by DNA. Science 2007, 318, 1121–1125. [Google Scholar] [CrossRef]
- Liu, C.J.; Liu, Y.; Zhu, E.Q.; Zhang, Q.; Wei, X.P.; Wang, B. Cross-Inhibitor: A time-sensitive molecular circuit based on DNA strand displacement. Nucleic Acids Res. 2020, 48, 10691–10701. [Google Scholar] [CrossRef]
- Dong, J.T.; Ouyang, Y.; Wang, J.B.; O’Hagan, M.P.; Willner, I. Assembly of Dynamic Gated and Cascaded Transient DNAzyme Networks. ACS Nano 2022, 16, 6153–6164. [Google Scholar] [CrossRef]
- Yao, C.; Zhang, R.; Tang, J.P.; Yang, D.Y. Rolling circle amplification (RCA)-based DNA hydrogel. Nat. Protoc. 2021, 16, 5460–5483. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H.M.; Willner, I.; Wang, F.A. High-performance biosensing based on autonomous enzyme-free DNA circuits. Top. Curr. Chem. 2020, 378, 32. [Google Scholar] [CrossRef]
- Dirks, R.M.; Pierce, N.A. Triggered amplification by hybridization chain reaction. Proc. Natl. Acad. Sci. USA 2004, 101, 15275–15278. [Google Scholar] [CrossRef]
- Li, C.C.; Zhang, J.J.; Gao, Y.S.; Luo, S.S.; Wu, Z.S. Nonenzymatic Autonomous Assembly of Cross-Linked Network Structures from Only Two Palindromic DNA Components for Intracellular Fluorescence Imaging of miRNAs. ACS Sens. 2022, 7, 601–611. [Google Scholar] [CrossRef]
- Song, X.L.; Ding, Q.; Zhang, J.; Sun, R.L.; Yin, L.H.; Wei, W.; Pu, Y.P.; Liu, S.Q. Smart Catalyzed Hairpin Assembly-Induced DNAzyme Nanosystem for Intracellular UDG Imaging. Anal. Chem. 2021, 93, 13687–13693. [Google Scholar] [CrossRef] [PubMed]
- He, S.Z.; Yu, S.S.; Li, R.M.; Chen, Y.Y.; Wang, Q.; He, Y.Q.; Liu, X.Q.; Wang, F.A. On-Site Non-enzymatic Orthogonal Activation of a Catalytic DNA Circuit for Self-Reinforced In Vivo MicroRNA Imaging. Angew. Chem. Int. Edit. 2022, 61, 9. [Google Scholar] [CrossRef]
- Yin, P.; Choi, H.M.T.; Calvert, C.R.; Pierce, N.A. Programming biomolecular self-assembly pathways. Nature 2008, 451, 318–322. [Google Scholar] [CrossRef]
- Wang, J.; Shimada, N.; Maruyama, A. Cationic Copolymer-Augmented DNA Hybridization Chain Reaction. ACS Appl. Mater. Interfaces 2022, 14, 39396–39403. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, Q.; Chen, Y.Q.; Liu, X.Q.; Wang, F.; Zhou, X. Amplified MicroRNA Detection and Intracellular Imaging Based on an Autonomous and Catalytic Assembly of DNAzyme. ACS Sens. 2019, 4, 110–117. [Google Scholar] [CrossRef]
- Song, J.; Mou, H.Z.; Li, X.Q.; Liu, Y.; Yang, X.J.; Chen, H.Y.; Xu, J.J. Self-assembled DNA/RNA nanospheres with cascade signal amplification for intracellular MicroRNA imaging. Sens. Actuator B Chem. 2022, 360, 8. [Google Scholar] [CrossRef]
- Dong, C.; Fang, X.; Xiong, J.; Zhang, J.; Gan, H.; Song, C.; Wang, L. Simultaneous Visualization of Dual Intercellular Signal Transductions via SERS Imaging of Membrane Proteins Dimerization on Single Cells. ACS Nano 2022, 16, 14055–14065. [Google Scholar] [CrossRef]
- Cairns, M.J.; Hopkins, T.M.; Witherington, C.; Wang, L.; Sun, L.Q. Target site selection for an RNA-cleaving catalytic DNA. Nat. Biotechnol. 1999, 17, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, Y.; Liu, Z.L.; Lin, B.; Yi, H.B.; Xu, F.; Nie, Z.; Yao, S.Z. Insight into G-quadruplex-hemin DNAzyme/RNAzyme: Adjacent adenine as the intramolecular species for remarkable enhancement of enzymatic activity. Nucleic Acids Res. 2016, 44, 7373–7384. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.F.; Deng, F.; Liu, Z.; Shi, G.; Chen, J.H. Toehold-Mediated Cascade Catalytic Assembly for Mycotoxin Detection and Its Logic Applications. Anal. Chem. 2022, 94, 3693–3700. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Gong, J.M.; Li, S.Q.; Liu, C.Y.; Zhou, L.L.; Sheng, J.H.; Xu, Q.X. Enzyme-free dual-DNA walker based on catalytic hairpin assembled DNAzyme for sensing telomerase activity. Sens. Actuator B Chem. 2021, 329, 7. [Google Scholar] [CrossRef]
- Liu, X.Y.; Zhou, X.M.; Xia, X.Y.; Xiang, H. Catalytic hairpin assembly-based double-end DNAzyme cascade-feedback amplification for sensitive fluorescence detection of HIV-1 DNA. Anal. Chim. Acta 2020, 1096, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.R.; Weng, B.R.; Liu, S.J.; Kang, N.N.; Ran, J.B.; Deng, Z.S.; Wang, H.M.; Yang, C.Y.; Wang, F. Acid-improved DNAzyme-based chemiluminescence miRNA assay coupled with enzyme-free concatenated DNA circuit. Biosens. Bioelectron. 2022, 204, 7. [Google Scholar] [CrossRef]
- Huang, W.; Zhan, D.Y.; Xie, Y.M.; Li, X.; Lai, G.S. Dual CHA-mediated high-efficient formation of a tripedal DNA walker for constructing a novel proteinase-free dual-mode biosensing strategy. Biosens. Bioelectron. 2022, 197, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.X.; Brennan, J.D.; Li, Y.F. A Multi-component All-DNA Biosensing System Controlled by a DNAzyme. Angew. Chem.-Int. Edit. 2020, 59, 10401–10405. [Google Scholar] [CrossRef]
- Chuaychob, S.; Fujita, M.; Maeda, M. G-Quadruplex-Functionalized Gold Nanoparticles for a Real-Time Biomolecule Sensor with On-Demand Tunable Properties. Langmuir 2022, 38, 4870–4878. [Google Scholar] [CrossRef]
- Zheng, X.D.; Yang, J.; Zhou, C.J.; Zhang, C.; Zhang, Q.; Wei, X.P. Allosteric DNAzyme-based DNA logic circuit: Operations and dynamic analysis. Nucleic Acids Res. 2019, 47, 1097–1109. [Google Scholar] [CrossRef]
- Gao, Y.H.; Chen, Y.Y.; Shang, J.H.; Yu, S.S.; He, S.Z.; Cui, R.; Wang, F.A. Enzyme-Free Autocatalysis-Driven Feedback DNA Circuits for Amplified Aptasensing of Living Cells. ACS Appl. Mater. Interfaces 2022, 14, 5080–5089. [Google Scholar] [CrossRef] [PubMed]
- Harding, B.I.; Pollak, N.M.; Stefanovic, D.; Macdonald, J. Repeated Reuse of Deoxyribozyme-Based Logic Gates. Nano Lett. 2019, 19, 7655–7661. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, R.; Cui, S.; Zhang, X.; Shi, P.; Zhou, S.; Wang, B. Programming DNA Reaction Networks Using Allosteric DNA Hairpins. Biomolecules 2023, 13, 481. https://doi.org/10.3390/biom13030481
Qin R, Cui S, Zhang X, Shi P, Zhou S, Wang B. Programming DNA Reaction Networks Using Allosteric DNA Hairpins. Biomolecules. 2023; 13(3):481. https://doi.org/10.3390/biom13030481
Chicago/Turabian StyleQin, Rui, Shuang Cui, Xiaokang Zhang, Peijun Shi, Shihua Zhou, and Bin Wang. 2023. "Programming DNA Reaction Networks Using Allosteric DNA Hairpins" Biomolecules 13, no. 3: 481. https://doi.org/10.3390/biom13030481
APA StyleQin, R., Cui, S., Zhang, X., Shi, P., Zhou, S., & Wang, B. (2023). Programming DNA Reaction Networks Using Allosteric DNA Hairpins. Biomolecules, 13(3), 481. https://doi.org/10.3390/biom13030481