

The Cd/Zn Axis: Emerging Concepts in Cellular Fate and Cytotoxicity


{kind=link}
Abstract
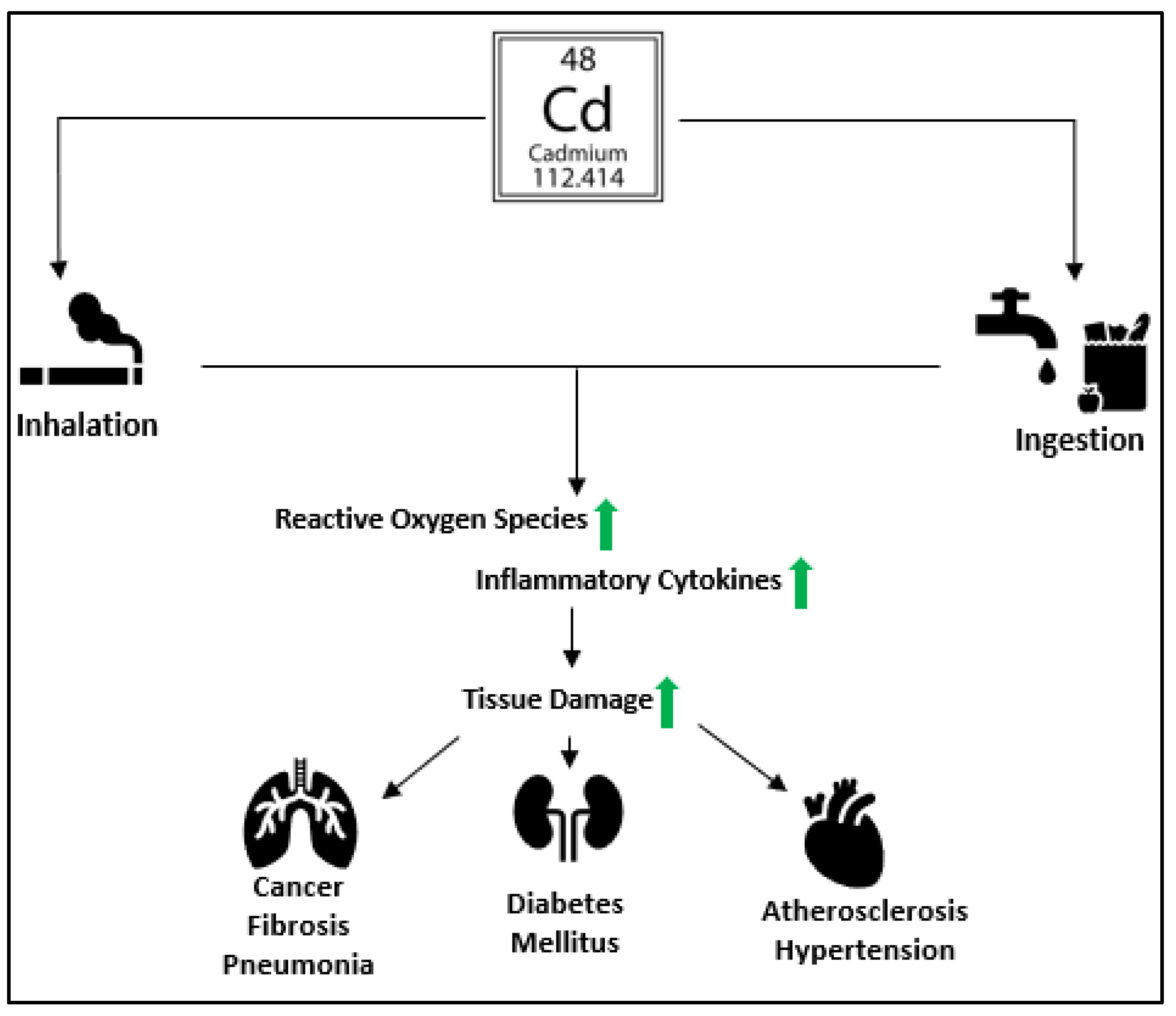
1. Introduction
2. Impacts on Immune Homeostasis
3. Oxidative Stress & Organelle Dysregulation
4. Tissue Retention and Competitive Binding
5. Oxidative Damage Therapies
6. Zn and Cd
7. Zn Is a Promising Therapeutic
8. An Emerging Concept of Cellular Fate
9. Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Compliance with Ethical Standards:
References
- Alghasham, A.; Salem, T.A.; Meki, A.R. Effect of cadmium-polluted water on plasma levels of tumor necrosis factor-α, interleukin-6 and oxidative status biomarkers in rats: Protective effect of curcumin. Food Chem. Toxicol. 2013, 59, 160–164. [Google Scholar] [CrossRef]
- Kim, Y.N.; Kim, J.S.; Seo, S.G.; Lee, Y.; Baek, S.W.; Kim, I.S.; Yoon, H.S.; Kim, K.R.; Kim, S.H.; Kim, K.H. Cadmium resistance in tobacco plants expressing the MuSI gene. Plant. Biotechnol. Rep. 2011, 5, 323–329. [Google Scholar] [CrossRef]
- Kulas, J.; Ninkov, M.; Tucovic, D.; Popov Aleksandrov, A.; Ukropina, M.; Cakic Milosevic, M.; Mutic, J.; Kataranovski, M.; Mikrov, I. Subchronic Oral Cadmium Exposure Exerts both Stimulatory and Suppressive Effects on Pulmonary Inflammation/Immune Reactivity in Rats. Biomed. Environ. Sci. 2019, 32, 508–519. [Google Scholar] [CrossRef]
- Baba, H.; Tsuneyama, K.; Yazaki, M.; Nagata, K.; Minamisaka, T.; Tsuda, T.; Nomoto, K.; Hayashi, S.; Miwa, S.; Nakajima, T.; et al. The liver in itai-itai disease (chronic cadmium poisoning): Pathological features and metallothionein expression. Mod. Pathol. 2013, 26, 1228–1234. [Google Scholar] [CrossRef]
- Mirkov, I.; Popov Aleksandrov, A.; Ninkov, M.; Tucovic, D.; Kulas, J.; Zeljkovic, M.; Popovic, D.; Kataranovski, M. Immunotoxicology of cadmium: Cells of the immune system as targets and effectors of cadmium toxicity. Food Chem. Toxicol. 2021, 149, 112026. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, X.; Sun, S.; Li, Q.; Xie, Y.; Li, Q.; Zhao, Y.; Pei, J.; Zhang, W.; Xue, P.; et al. Cadmium modulates hematopoietic stem and progenitor cells and skews toward myelopoiesis in mice. Toxicol. Appl. Pharm. 2016, 313, 24–34. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Q.; Yang, Z.; Shao, Y.; Xue, P.; Qu, W.; Jia, X.; Cheng, L.; He, M.; He, R.; et al. Cadmium Activates Noncanonical Wnt Signaling to Impair Hematopoietic Stem Cell Function in Mice. Toxicol. Sci. 2018, 165, 254–266. [Google Scholar] [CrossRef]
- Beryllium, cadmium, mercury, and exposures in the glass manufacturing industry. IARC Monogr. Eval. Carcinog. Risks Hum. 1993, 58, 1–415.
- Zhang, X.; Ma, L.; Tang, Y.; Han, J.; Qi, Y.; Huang, D. Low-dose cadmium exposure facilitates cell proliferation by promoter hypermethylation of RASSF1A and DAPK1 genes. Environ. Toxicol. 2021, 36, 2313–2321. [Google Scholar] [CrossRef]
- Kulkarni, P.; Dasgupta, P.; Bhat, N.S.; Hashimoto, Y.; Saini, S.; Shahryari, V.; Yamamura, S.; Shiina, M.; Tanaka, Y.; Dahiya, R.; et al. Role of the PI3K/Akt pathway in cadmium induced malignant transformation of normal prostate epithelial cells. Toxicol. Appl. Pharm. 2020, 409, 115308. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, X.; Sun, D.; Cao, J.; Zhang, Y.; Huo, X. Alteration of the number and percentage of innate immune cells in preschool children from an e-waste recycling area. Ecotoxicol. Environ. Saf. 2017, 145, 615–622. [Google Scholar] [CrossRef]
- Goyal, T.; Mitra, P.; Singh, P.; Sharma, S.; Purohit, P.; Sharma, P. Effect of occupational co-exposure to lead and cadmium on selected immunomodulatory cytokines. Toxicol. Ind. Health 2022, 38, 1–10. [Google Scholar] [CrossRef]
- Chen, X.; Bi, M.; Yang, J.; Cai, J.; Zhang, H.; Zhu, Y.; Zheng, Y.; Liu, Q.; Shi, G.; Zhang, Z. Cadmium exposure triggers oxidative stress, necroptosis, Th1/Th2 imbalance and promotes inflammation through the TNF-α/NF-κB pathway in swine small intestine. J. Hazard. Mater. 2022, 421, 126704. [Google Scholar] [CrossRef]
- Zheng, X.; Xu, X.; Lu, F.; Wang, Q.; Zeng, Z.; Huo, X. High serum IgG subclass concentrations in children with e-waste Pb and Cd exposure. Sci. Total Environ. 2021, 764, 142806. [Google Scholar] [CrossRef]
- Zahedi, A.; Hassanvand, M.S.; Jaafarzadeh, N.; Ghadiri, A.; Shamsipour, M.; Dehcheshmeh, M.G. Increased allergic and asthmatic risks in children residing in industrial areas by surveying the pre-inflammatory (IgE, IL-4 and IL-13) biomarkers. J. Environ. Health Sci. Eng. 2022, 20, 609–615. [Google Scholar] [CrossRef]
- Kim, E.; Bonnegarde-Bernard, A.; Opiyo, S.O.; Joldrichsen, M.R.; Attia, Z.; Ahmer, B.H.; Cormet-Boyaka, E.; Boyaka, P.N. Pollutants enhance IgE sensitization in the gut via local alteration of vitamin D-metabolizing enzymes. Mucosal. Immunol. 2022, 15, 143–153. [Google Scholar] [CrossRef]
- Knoell, D.L.; Wyatt, T.A. The adverse impact of cadmium on immune function and lung host defense. Semin. Cell Dev. Biol. 2021, 115, 70–76. [Google Scholar] [CrossRef]
- Alkharashi, N.A.O.; Periasamy, V.S.; Athinarayanan, J.; Alshatwi, A.A. Cadmium triggers mitochondrial oxidative stress in human peripheral blood lymphocytes and monocytes: Analysis using in vitro and system toxicology approaches. J. Trace Elem. Med. Biol. 2017, 42, 117–128. [Google Scholar] [CrossRef]
- Sarmiento-Ortega, V.E.; Moroni-González, D.; Diaz, A.; Brambila, E.; Treviño, S. ROS and ERK Pathway Mechanistic Approach on Hepatic Insulin Resistance After Chronic Oral Exposure to Cadmium NOAEL Dose. Biol. Trace Elem. Res. 2022. [Google Scholar] [CrossRef]
- Skalny, A.V.; Lima, T.R.R.; Ke, T.; Zhou, J.C.; Bornhorst, J.; Alekseenko, S.I.; Aaseth, J.; Anesti, O.; Sarigiannis, D.A.; Tsatsakis, A.; et al. Corrigendum to “Toxic metal exposure as a possible risk factor for COVID-19 and other respiratory infectious diseases” [Food Chem. Toxicol. 146 111809]. Food Chem. Toxicol. 2021, 149, 111999. [Google Scholar] [CrossRef]
- Branca, J.J.V.; Pacini, A.; Gulisano, M.; Taddei, N.; Fiorillo, C.; Becatti, M. Cadmium-Induced Cytotoxicity: Effects on Mitochondrial Electron Transport Chain. Front. Cell Dev. Biol. 2020, 8, 604377. [Google Scholar] [CrossRef]
- Wang, Y.; Chi, H.; Xu, F.; He, Z.; Li, Z.; Wu, F.; Li, Y.; Zhang, G.; Peng, X.; Yu, S.; et al. Cadmium chloride-induced apoptosis of HK-2 cells via interfering with mitochondrial respiratory chain. Ecotoxicol. Environ. Saf. 2022, 236, 113494. [Google Scholar] [CrossRef]
- Tian, J.; Li, Z.; Wang, L.; Qiu, D.; Zhang, X.; Xin, X.; Cai, Z.; Lei, B. Metabolic signatures for safety assessment of low-level cadmium exposure on human osteoblast-like cells. Ecotoxicol. Environ. Saf. 2021, 207, 111257. [Google Scholar] [CrossRef]
- Yuan, J.; Zhao, Y.; Bai, Y.; Gu, J.; Yuan, Y.; Liu, X.; Liu, Z.; Zou, H.; Bian, J. Cadmium induces endosomal/lysosomal enlargement and blocks autophagy flux in rat hepatocytes by damaging microtubules. Ecotoxicol. Environ. Saf. 2021, 228, 112993. [Google Scholar] [CrossRef]
- Gong, Z.G.; Zhao, Y.; Wang, Z.Y.; Fan, R.F.; Liu, Z.P.; Wang, L. Epigenetic regulator BRD4 is involved in cadmium-induced acute kidney injury via contributing to lysosomal dysfunction, autophagy blockade and oxidative stress. J. Hazard. Mater. 2022, 423, 127110. [Google Scholar] [CrossRef]
- Pi, H.; Li, M.; Tian, L.; Yang, Z.; Yu, Z.; Zhou, Z. Enhancing lysosomal biogenesis and autophagic flux by activating the transcription factor EB protects against cadmium-induced neurotoxicity. Sci. Rep. 2017, 7, 43466. [Google Scholar] [CrossRef]
- Cheng, J.; Liu, D.; Zhao, L.; Zhao, Q.; Zhang, X.; Wang, B.; Bai, D. Potentilla anserine L. polysaccharide inhibits cadmium-induced neurotoxicity by attenuating autophagy. Neurochem. Int. 2021, 147, 105045. [Google Scholar] [CrossRef]
- Ng, C.S.; Wan, S.; Yim, A.P. Pulmonary ischaemia-reperfusion injury: Role of apoptosis. Eur. Respir J. 2005, 25, 356–363. [Google Scholar] [CrossRef]
- Fujishiro, H.; Yano, Y.; Takada, Y.; Tanihara, M.; Himeno, S. Roles of ZIP8, ZIP14, and DMT1 in transport of cadmium and manganese in mouse kidney proximal tubule cells. Metallomics 2012, 4, 700–708. [Google Scholar] [CrossRef]
- Fujie, T.; Ito, K.; Ozaki, Y.; Takahashi, S.; Yamamoto, C.; Kaji, T. Induction of ZIP8, a ZIP transporter, via NF-κB signaling by the activation of IκBα and JNK signaling in cultured vascular endothelial cells exposed to cadmium. Toxicol. Appl. Pharm. 2022, 434, 115802. [Google Scholar] [CrossRef]
- Fujishiro, H.; Miyamoto, S.; Sumi, D.; Kambe, T.; Himeno, S. Effects of individual amino acid mutations of zinc transporter ZIP8 on manganese- and cadmium-transporting activity. Biochem. Biophys. Res. Commun. 2022, 616, 26–32. [Google Scholar] [CrossRef]
- Fujishiro, H.; Sumino, M.; Sumi, D.; Umemoto, H.; Tsuneyama, K.; Matsukawa, T.; Yokoyama, K.; Himeno, S. Spatial localization of cadmium and metallothionein in the kidneys of mice at the early phase of cadmium accumulation. J. Toxicol. Sci. 2022, 47, 507–517. [Google Scholar] [CrossRef]
- Wang, X.; Gong, B.; He, E.; Peijnenburg, W.; Qiu, H. Subcellular localization and compartment-specific toxicokinetics of cadmium, arsenic, and zinc in brandling worm Eisenia fetida. Chemosphere 2022, 308, 136482. [Google Scholar] [CrossRef]
- Jan, S.; Anna, C.; Antonín, K.; Jiří, Š.; Jan, B.; Tereza, L.; Pavel, K. Intracellular sequestration of cadmium and zinc in ectomycorrhizal fungus Amanita muscaria (Agaricales, Amanitaceae) and characterization of its metallothionein gene. Fungal. Genet. Biol. 2022, 162, 103717. [Google Scholar] [CrossRef]
- Nasiadek, M.; Stragierowicz, J.; Kilanowicz, A. An Assessment of Metallothionein-Cadmium Binding in Rat Uterus after Subchronic Exposure Using a Long-Term Observation Model. Int. J. Mol. Sci. 2022, 23, 5154. [Google Scholar] [CrossRef]
- Cadmium; OSHA, OSaHA, Eds.; United States Department of Labor: Washington, DC, USA, 1992; Volume 1910.1027(l)(3)(iii)(A). [Google Scholar]
- Kim, J.J.; Kim, Y.S.; Kumar, V. Heavy metal toxicity: An update of chelating therapeutic strategies. J. Trace Elem. Med. Biol. 2019, 54, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.J.; Chisolm, J., Jr. Lead, zinc and copper decorporation during calcium disodium ethylenediamine tetraacetate treatment of lead-poisoned children. J. Pharm. Exp. 1986, 239, 829–835. [Google Scholar]
- Innico, G.; Gobbi, L.; Bertoldi, G.; Rigato, M.; Basso, A.; Bonfante, L.; Calò, L.A. Oxidative stress, inflammation, and peritoneal dialysis: A molecular biology approach. Artif. Organs. 2021, 45, 1202–1207. [Google Scholar] [CrossRef]
- Eder, K.; Siebers, M.; Most, E.; Scheibe, S.; Weissmann, N.; Gessner, D.K. An excess dietary vitamin E concentration does not influence Nrf2 signaling in the liver of rats fed either soybean oil or salmon oil. Nutr. Metab. 2017, 14, 71. [Google Scholar] [CrossRef]
- Tandon, S.K.; Singh, S.; Dhawan, M. Preventive effect of vitamin E in cadmium intoxication. Biomed. Environ. Sci. 1992, 5, 39–45. [Google Scholar]
- Fang, J.; Xie, S.; Chen, Z.; Wang, F.; Chen, K.; Zuo, Z.; Cui, H.; Guo, H.; Ouyang, P.; Chen, Z.; et al. Protective Effect of Vitamin E on Cadmium-Induced Renal Oxidative Damage and Apoptosis in Rats. Biol. Trace Elem. Res. 2021, 199, 4675–4687. [Google Scholar] [CrossRef]
- ElMahdy, M.K.; Zaki, M.O.; Al-Karmalawy, A.A.; Abdo, W.; Alnasser, S.M.; Antar, S.A. Glimepiride ameliorates renal toxicity induced by cadmium in mice: Modulation of Jun N terminal kinase (JNK)/nuclear factor kappa B (NF-κB) and phosphatidylinositol 3-kinases (PI3K)/protein kinase (AKT) pathways. Life Sci. 2022, 311, 121184. [Google Scholar] [CrossRef]
- Ben Mrid, R.; Ennoury, A.; Roussi, Z.; Naboulsi, I.; Benmrid, B.; Kchikich, A.; El Omari, R.; Nhiri, M.; Yasri, A. Thymoquinone Alleviates Cadmium Induced Stress in Germinated Lens culinaris Seeds by Reducing Oxidative Stress and Increasing Antioxidative Activities. Life 2022, 12, 1779. [Google Scholar] [CrossRef]
- Martin Molinero, G.D.; Boldrini, G.G.; Pérez Chaca, M.V.; Moyano, M.F.; Armonelli Fiedler, S.; Giménez, M.S.; Gómez, N.N.; López, P.H.H.; Álvarez, S.M. A soybean based-diet prevents Cadmium access to rat cerebellum, maintaining trace elements homeostasis and avoiding morphological alterations. Biometals 2022, 36, 67–96. [Google Scholar] [CrossRef]
- Szuster-Ciesielska, A.; Stachura, A.; Słotwińska, M.; Kamińska, T.; Sniezko, R.; Paduch, R.; Abramczyk, D.; Filar, J.; Kandefer-Szerszeń, M. The inhibitory effect of zinc on cadmium-induced cell apoptosis and reactive oxygen species (ROS) production in cell cultures. Toxicology 2000, 145, 159–171. [Google Scholar] [CrossRef]
- Ebaid, H.; Hassan, I.; Bashandy, S.; Taha, N.A.; Mahmood, A.; Alomar, S.; Alhazza, I.; Mashaly, A.; Rady, A. Zinc improves the immune function and the proliferation of lymphocytes in Cadmium-treated rats. Cent. Eur. J. Immunol. 2014, 39, 441–448. [Google Scholar] [CrossRef]
- Pan, J.; Huang, X.; Li, Y.; Li, M.; Yao, N.; Zhou, Z.; Li, X. Zinc protects against cadmium-induced toxicity by regulating oxidative stress, ions homeostasis and protein synthesis. Chemosphere 2017, 188, 265–273. [Google Scholar] [CrossRef]
- Arao, Y.; Korach, K.S. The physiological role of estrogen receptor functional domains. Essays. Biochem. 2021, 65, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Tarhonska, K.; Lesicka, M.; Janasik, B.; Roszak, J.; Reszka, E.; Braun, M.; Kołacińska-Wow, A.; Jabłońska, E. Cadmium and breast cancer—Current state and research gaps in the underlying mechanisms. Toxicol. Lett. 2022, 361, 29–42. [Google Scholar] [CrossRef]
- Liu, W.B.; Zhu, H.L.; Xiong, Y.W.; Lv, J.; Huang, Y.C.; Wang, H. Environmental cadmium exposure during gestation impairs fetal brain and cognitive function of adult offspring via reducing placenta-derived E2 level. Chemosphere 2022, 307, 135668. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Moniuszko-Jakoniuk, J. Interactions between cadmium and zinc in the organism. Food Chem. Toxicol. 2001, 39, 967–980. [Google Scholar] [CrossRef]
- Earley, B.J.; Cubillas, C.; Warnhoff, K.; Ahmad, R.; Alcantar, A.; Lyon, M.D.; Schneider, D.L.; Kornfeld, K. Cadmium hijacks the high zinc response by binding and activating the HIZR-1 nuclear receptor. Proc. Natl. Acad. Sci. USA 2021, 118, e2022649118. [Google Scholar] [CrossRef]
- Skalny, A.V.; Rink, L.; Ajsuvakova, O.P.; Aschner, M.; Gritsenko, V.A.; Alekseenko, S.I.; Svistunov, A.A.; Petrakis, D.; Spandidos, D.A.; Aaseth, J.; et al. Zinc and respiratory tract infections: Perspectives for COVID-19 (Review). Int. J. Mol. Med. 2020, 46, 17–26. [Google Scholar] [CrossRef]
- Knoell, D.L.; Smith, D.A.; Sapkota, M.; Heires, A.J.; Hanson, C.K.; Smith, L.M.; Poole, J.A.; Wyatt, T.A.; Romberger, D.J. Insufficient zinc intake enhances lung inflammation in response to agricultural organic dust exposure. J. Nutr. Biochem. 2019, 70, 56–64. [Google Scholar] [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Malireddi, R.K.S.; Kesavardhana, S.; Kanneganti, T.D. ZBP1 and TAK1: Master Regulators of NLRP3 Inflammasome/Pyroptosis, Apoptosis, and Necroptosis (PAN-optosis). Front. Cell Infect. Microbiol. 2019, 9, 406. [Google Scholar] [CrossRef]
- Samir, P.; Malireddi, R.K.S.; Kanneganti, T.D. The PANoptosome: A Deadly Protein Complex Driving Pyroptosis, Apoptosis, and Necroptosis (PANoptosis). Front. Cell. Infect. Microbiol. 2020, 10, 238. [Google Scholar] [CrossRef]
- Christgen, S.; Zheng, M.; Kesavardhana, S.; Karki, R.; Malireddi, R.K.S.; Banoth, B.; Place, D.E.; Briard, B.; Sharma, B.R.; Tuladhar, S.; et al. Identification of the PANoptosome: A Molecular Platform Triggering Pyroptosis, Apoptosis, and Necroptosis (PANoptosis). Front. Cell Infect. Microbiol. 2020, 10, 237. [Google Scholar] [CrossRef]
- Zhou, J.; Zeng, L.; Zhang, Y.; Wang, M.; Li, Y.; Jia, Y.; Wu, L.; Su, P. Cadmium exposure induces pyroptosis in testicular tissue by increasing oxidative stress and activating the AIM2 inflammasome pathway. Sci. Total Environ. 2022, 847, 157500. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, J.; Yang, J.; Cai, J.; Liu, Q.; Zhang, X.; Bao, J.; Zhang, Z. Cadmium induces apoptosis and autophagy in swine small intestine by downregulating the PI3K/Akt pathway. Environ. Sci. Pollut Res. Int. 2022, 29, 41207–41218. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johns, C.E.; Gattu, M.; Camilli, S.; Desaraju, A.; Kolliputi, N.; Galam, L. The Cd/Zn Axis: Emerging Concepts in Cellular Fate and Cytotoxicity. Biomolecules 2023, 13, 316. https://doi.org/10.3390/biom13020316
Johns CE, Gattu M, Camilli S, Desaraju A, Kolliputi N, Galam L. The Cd/Zn Axis: Emerging Concepts in Cellular Fate and Cytotoxicity. Biomolecules. 2023; 13(2):316. https://doi.org/10.3390/biom13020316
Chicago/Turabian StyleJohns, Colleen Elsa, Mrudula Gattu, Samuel Camilli, Apoorva Desaraju, Narasaiah Kolliputi, and Lakshmi Galam. 2023. "The Cd/Zn Axis: Emerging Concepts in Cellular Fate and Cytotoxicity" Biomolecules 13, no. 2: 316. https://doi.org/10.3390/biom13020316
APA StyleJohns, C. E., Gattu, M., Camilli, S., Desaraju, A., Kolliputi, N., & Galam, L. (2023). The Cd/Zn Axis: Emerging Concepts in Cellular Fate and Cytotoxicity. Biomolecules, 13(2), 316. https://doi.org/10.3390/biom13020316