
Identification of an l-Arabitol Transporter from Aspergillus niger


Abstract
1. Introduction
2. Materials and Methods
2.1. Strains, Media and Growth Conditions
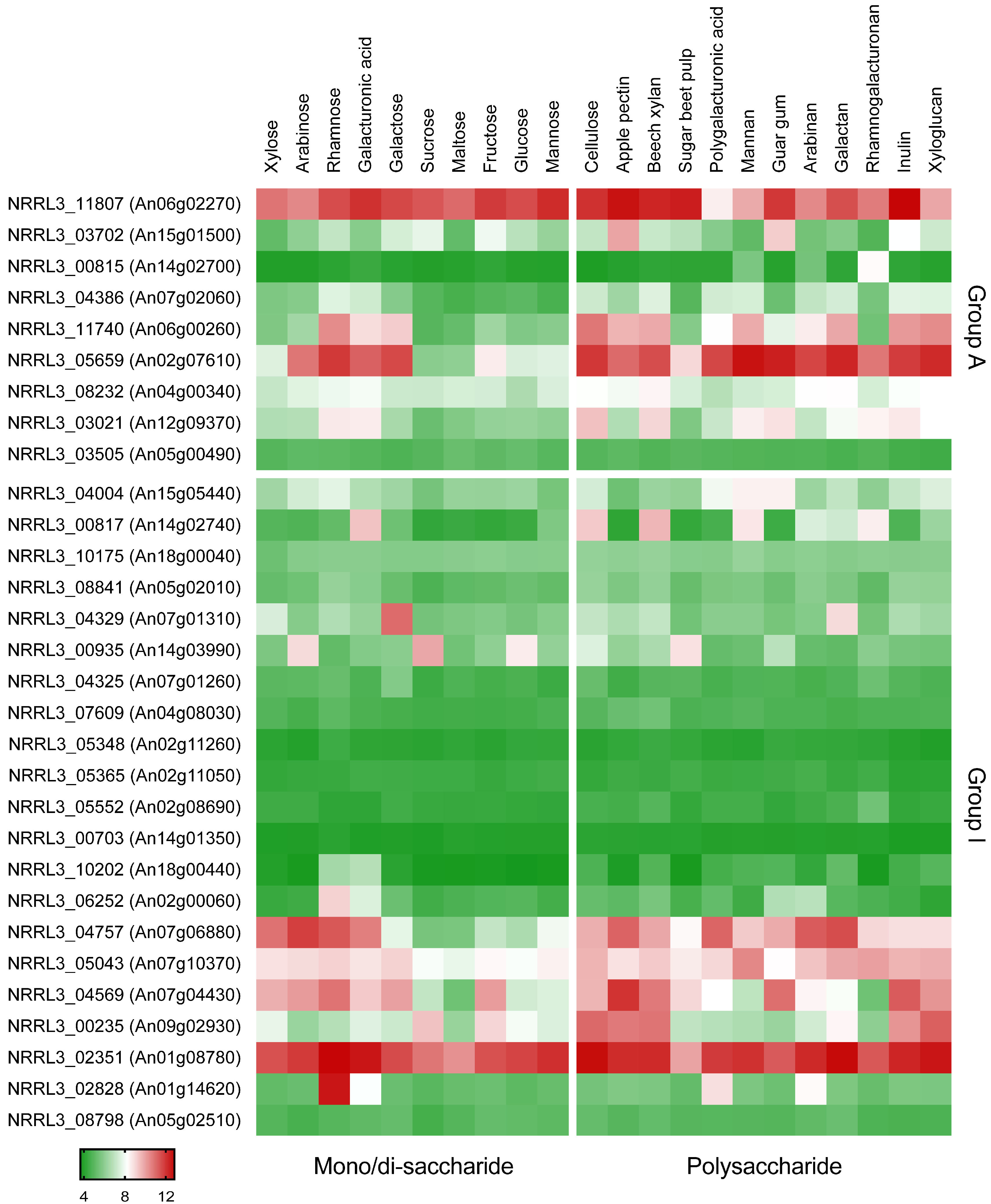
2.2. Expression Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | CBS Number | Genotype | Reference |
|---|---|---|---|
| N593 Δku70 | CBS 138852 | cspA1, kusA::amdS, pyrG¯ | [29] |
| ΔlatA | CBS 147737 | cspA1, kusA::amdS, pyrG¯, latA¯ | This study |
| Δ05659 | CBS 147735 | cspA1, kusA::amdS, pyrG¯, 05659¯ | This study |
| ΔladAΔxdhAΔsdhA | CBS 144672 | cspA1, kusA::amdS, pyrG¯, ladA¯, xdhA¯, sdhA¯ | [10] |
| ΔladAΔxdhAΔsdhAΔlatA | CBS 149003 | cspA1, kusA::amdS, pyrG¯, ladA¯, xdhA¯, sdhA¯, latA¯ | This study |
2.3. Transfer and Sugar Consumption Experiments
2.4. Quantification of Monosaccharides and Polyols
2.5. Phylogenetic Analysis
3. Results and Discussion
3.1. Identification of Putative l-Arabitol Transporters
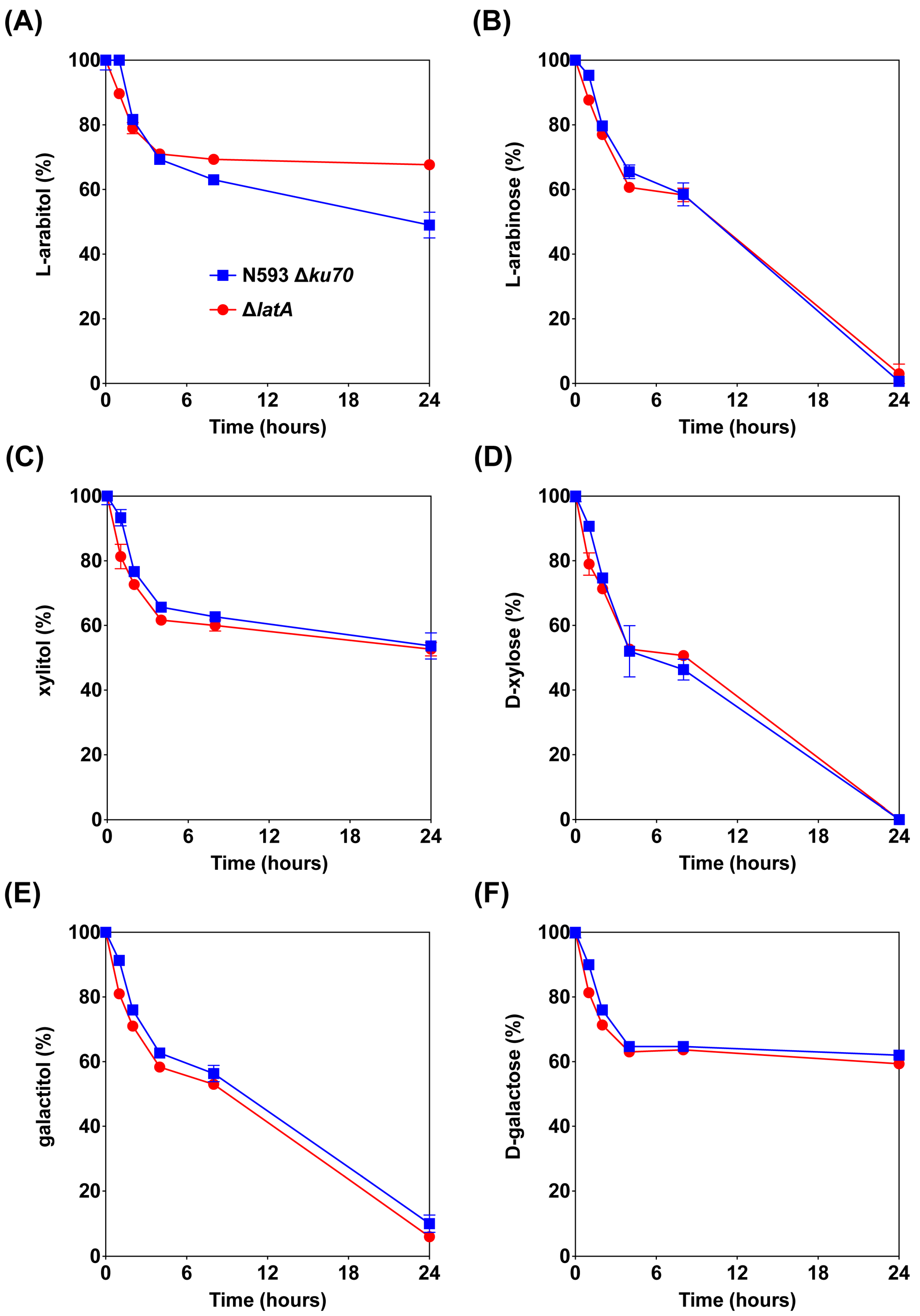
3.1.1. Deletion of latA Resulted in Impaired Growth on l-Arabitol
3.1.2. Deletion of latA Only Affected l-Arabitol Consumption
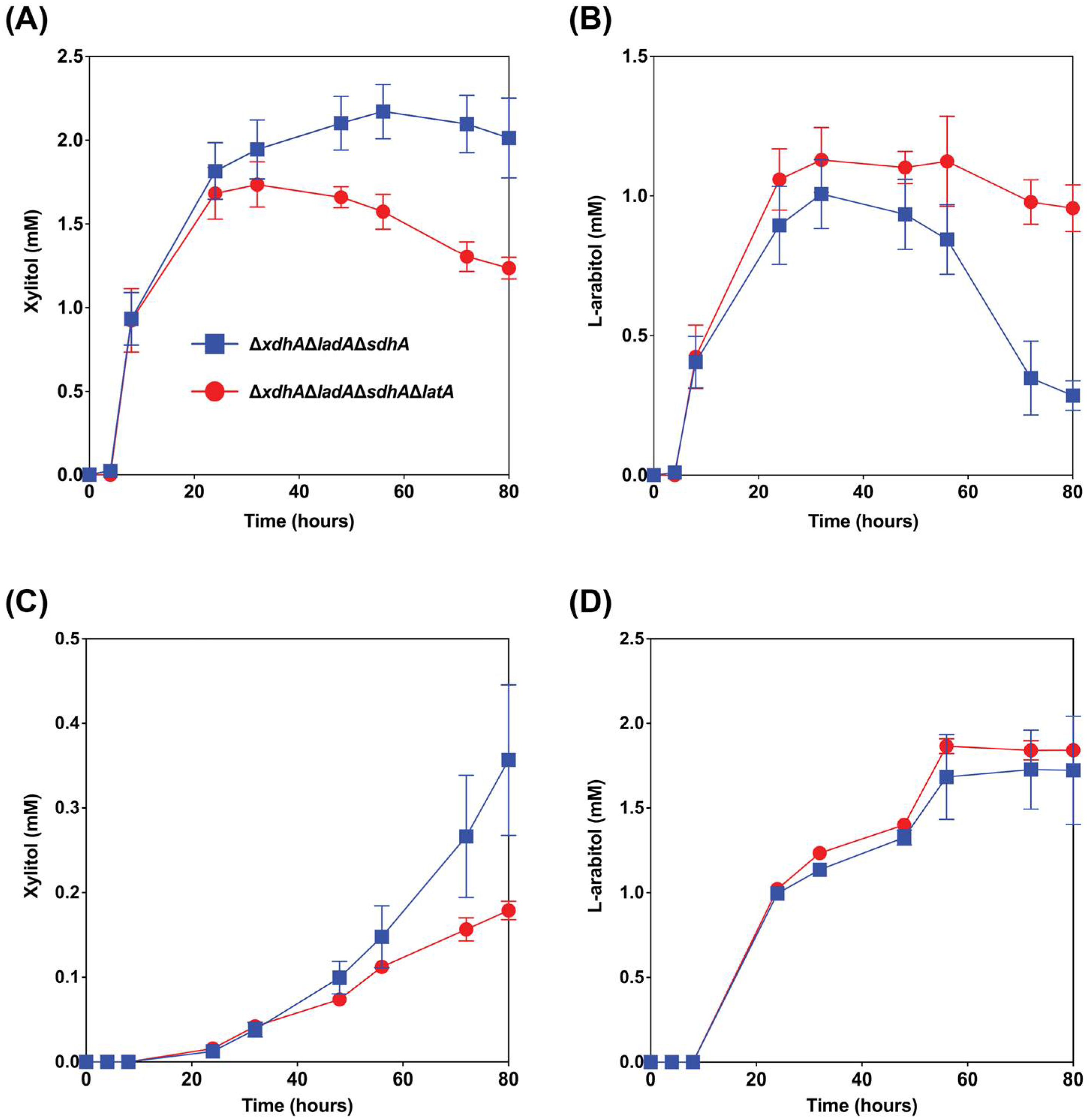
3.1.3. The deletion of latA Positively Affected l-Arabitol Production from Wheat Bran and Sugar Beet Pulp
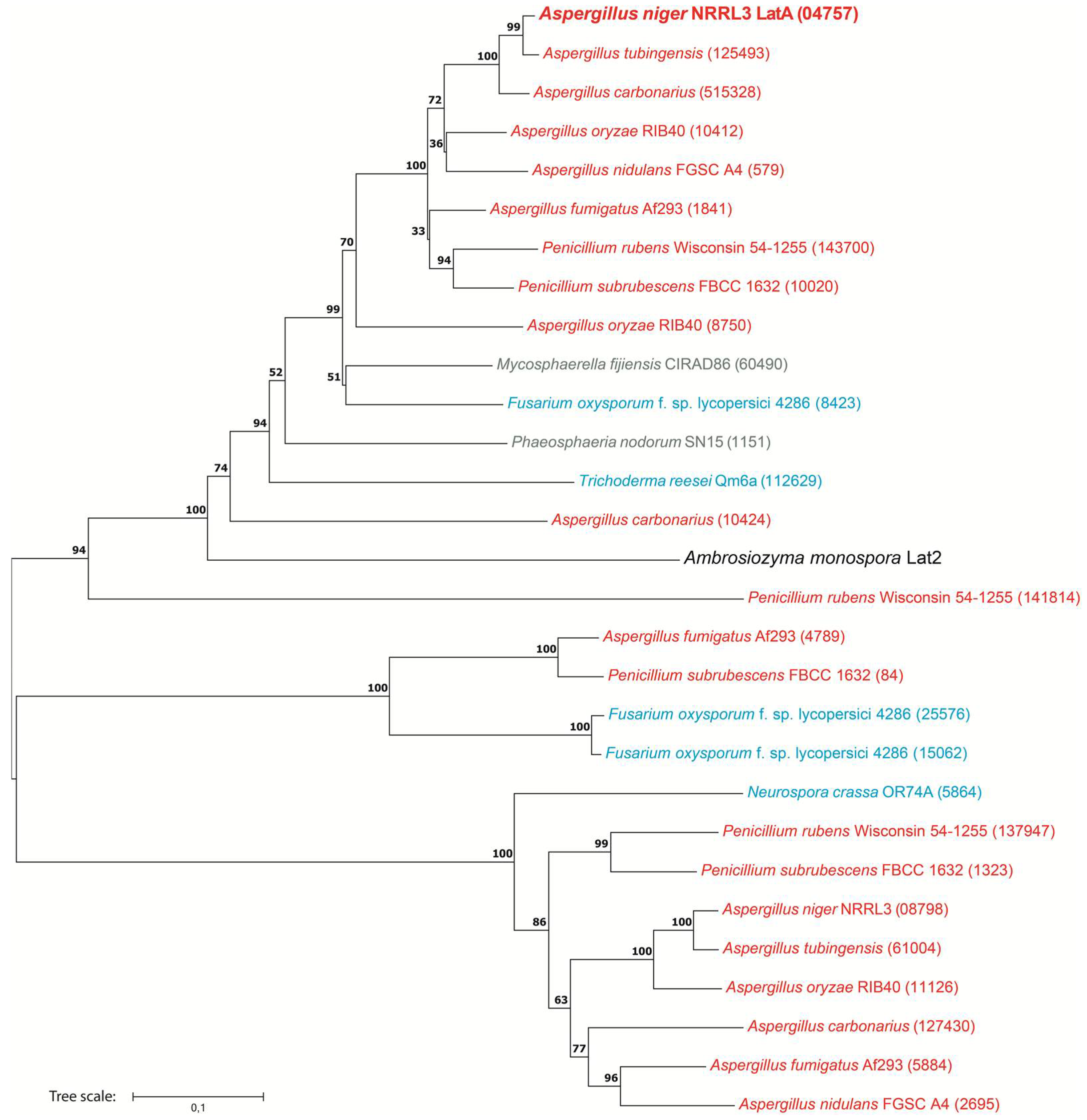
3.1.4. LatA Is Widely Present in Ascomycete Fungi
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jennings, D.B.; Ehrenshaft, M.; Pharr, D.M.; Williamson, J.D. Roles for mannitol and mannitol dehydrogenase in active oxygen-mediated plant defense. Proc. Natl. Acad. Sci. USA 1998, 95, 15129–15133. [Google Scholar] [CrossRef]
- Lewis, D.H.; Smith, D.C. Sugar alcohols (polyols) in fungi and green plants. I. Distribution, physiology and metabolism. New Phytol. 1967, 66, 143–184. [Google Scholar] [CrossRef]
- Monedero, V.; Perez-Martinez, G.; Yebra, M.J. Perspectives of engineering lactic acid bacteria for biotechnological polyol production. Appl. Microbiol. Biotechnol. 2010, 86, 1003–1015. [Google Scholar] [CrossRef]
- Mortlock, R.P.; Fossitt, D.D.; Wood, W.A. A basis for utlization of unnatural pentoses and pentitols by Aerobacter aerogenes. Proc. Natl. Acad. Sci. USA 1965, 54, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Kordowska-Wiater, M. Production of arabitol by yeasts: Current status and future prospects. J. Appl. Microbiol. 2015, 119, 303–314. [Google Scholar] [CrossRef] [PubMed]
- de Groot, M.J.L.; van den Dool, C.; Wosten, H.A.B.; Levisson, M.; vanKuyk, P.A.; Ruijter, G.J.G.; de Vries, R.P. Regulation of pentose catabolic pathway genes of Aspergillus niger. Food Technol. Biotechnol. 2007, 45, 134–138. [Google Scholar]
- Mojzita, D.; Penttila, M.; Richard, P. Identification of an l-arabinose reductase gene in Aspergillus niger and its role in l-arabinose catabolism. J. Biol. Chem. 2010, 285, 23622–23628. [Google Scholar] [CrossRef]
- Battaglia, E.; Hansen, S.F.; Leendertse, A.; Madrid, S.; Mulder, H.; Nikolaev, I.; de Vries, R.P. Regulation of pentose utilisation by AraR, but not XlnR, differs in Aspergillus nidulans and Aspergillus niger. Appl. Microbiol. Biotechnol. 2011, 91, 387–397. [Google Scholar] [CrossRef]
- Battaglia, E.; Visser, L.; Nijssen, A.; van Veluw, J.; Wösten, H.A.B.; de Vries, R.P. Analysis of regulation of pentose utilisation in Aspergillus niger reveals evolutionary adaptations in the Eurotiales. Stud. Mycol. 2011, 69, 31–38. [Google Scholar] [CrossRef]
- Chroumpi, T.; Peng, M.; Aguilar-Pontes, M.V.; Muller, A.; Wang, M.; Yan, J.; Lipzen, A.; Ng, V.; Grigoriev, I.V.; Makela, M.R.; et al. Revisiting a ‘simple’ fungal metabolic pathway reveals redundancy, complexity and diversity. Microb. Biotechnol. 2021, 14, 2525–2537. [Google Scholar] [CrossRef]
- Witteveen, C.F.B.; Busink, R.; van de Vondervoort, P.; Dijkema, C.; Swart, K.; Visser, J. l-arabinose and d-xylose catabolism in Aspergillus niger. J. Gen. Microbiol. 1989, 135, 2163–2171. [Google Scholar] [CrossRef][Green Version]
- Meng, J.; Chroumpi, T.; Mäkelä, M.R.; de Vries, R.P. Xylitol production from plant biomass by Aspergillus niger through metabolic engineering. Bioresour. Technol 2021, 344, 126199. [Google Scholar] [CrossRef] [PubMed]
- Noiraud, N.; Maurousset, L.; Lemoine, R. Transport of polyols in higher plants. Plant Physiol. Biochem. 2001, 39, 717–728. [Google Scholar] [CrossRef]
- Reinders, A.; Panshyshyn, J.A.; Ward, J.W. Analysis of transport activity of Arabidopsis sugar alcohol permease homolog AtPLT5. J. Biol. Chem. 2005, 280, 1594–1602. [Google Scholar] [CrossRef] [PubMed]
- Schilling, S.; Oesterhelt, C. Structurally reduced monosaccharide transporters in an evolutionarily conserved red alga. Biochem. J. 2007, 406, 325–331. [Google Scholar] [CrossRef]
- Heuel, H.; Shakeri-Garakani, A.; Turgut, S.; Lengeler, J.W. Genes for d-arabinitol and ribitol catabolism from Klebsiella pneumoniae. Microbiology 1998, 144 Pt 6, 1631–1639. [Google Scholar] [CrossRef]
- Jordan, P.; Choe, J.Y.; Boles, E.; Oreb, M. Hxt13, Hxt15, Hxt16 and Hxt17 from Saccharomyces cerevisiae represent a novel type of polyol transporters. Sci. Rep. 2016, 6, 23502. [Google Scholar] [CrossRef]
- Londesborough, J.; Richard, P.; Valkonen, M.; Viljanen, K. Effect of C-terminal Protein tags on pentitol and l-arabinose transport by Ambrosiozyma monospora Lat1 and Lat2 transporters in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2014, 80, 2737–2745. [Google Scholar] [CrossRef]
- Verho, R.; Penttila, M.; Richard, P. Cloning of two genes (LAT1,2) encoding specific l-arabinose transporters of the l-arabinose fermenting yeast Ambrosiozyma monospora. Appl. Biochem. Biotechnol. 2011, 164, 604–611. [Google Scholar] [CrossRef]
- Pereira, I.; Madeira, A.; Prista, C.; Loureiro-Dias, M.C.; Leandro, M.J. Characterization of new polyol/H+ symporters in Debaryomyces hansenii. PLoS ONE 2014, 9, e88180. [Google Scholar] [CrossRef]
- Peng, M.; Aguilar-Pontes, M.V.; de Vries, R.P.; Mäkelä, M.R. In silico analysis of putative sugar transporter genes in Aspergillus niger using phylogeny and comparative transcriptomics. Front. Microbiol. 2018, 9, 1045. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lin, L.; Li, H.; Tian, C.; Ma, Y. Transcriptional comparison of the filamentous fungus Neurospora crassa growing on three major monosaccharides d-glucose, d-xylose and l-arabinose. Biotechnol. Biofuels 2014, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, S.; Zhao, H. Discovery and characterization of novel d-xylose-specific transporters from Neurospora crassa and Pichia stipitis. Mol. Biosyst. 2010, 6, 2150–2156. [Google Scholar] [CrossRef]
- Ferreira, C.; van Voorst, F.; Martins, A.; Neves, L.; Oliveira, R.; Kielland-Brandt, M.C.; Lucas, C.; Brandt, A. A member of the sugar transporter family, Stl1p is the glycerol/H+ symporter in Saccharomyces cerevisiae. Mol. Biol. Cell. 2005, 16, 2068–2076. [Google Scholar] [CrossRef]
- Song, L.; Ouedraogo, J.P.; Kolbusz, M.; Nguyen, T.T.M.; Tsang, A. Efficient genome editing using tRNA promoter-driven CRISPR/Cas9 gRNA in Aspergillus niger. PLoS ONE 2018, 13, e0202868. [Google Scholar] [CrossRef]
- Kowalczyk, J.E.; Lubbers, R.J.M.; Peng, M.; Battaglia, E.; Visser, J.; de Vries, R.P. Combinatorial control of gene expression in Aspergillus niger grown on sugar beet pectin. Sci. Rep. 2017, 7, 12356. [Google Scholar] [CrossRef]
- de Vries, R.; Burgers, K.; van de Vondervoort, P.; Frisvad, J.; Samson, R.; Visser, J. A new black Aspergillus species, A. vadensis, is a promising host for homologous and heterologous protein production. Appl. Environ. Microbiol. 2004, 70, 3954–3959. [Google Scholar] [CrossRef] [PubMed]
- Gruben, B.S.; Mäkelä, M.R.; Kowalczyk, J.E.; Zhou, M.M.; Benoit-Gelber, I.; De Vries, R.P. Expression-based clustering of CAZyme-encoding genes of Aspergillus niger. BMC Genom. 2017, 18, 900. [Google Scholar] [CrossRef]
- Meyer, V.; Arentshorst, M.; El-Ghezal, A.; Drews, A.C.; Kooistra, R.; van den Hondel, C.A.; Ram, A.F. Highly efficient gene targeting in the Aspergillus niger kusA mutant. J. Biotechnol. 2007, 128, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Mäkelä, M.R.; Bouzid, O.; Robl, D.; Post, H.; Peng, M.; Heck, A.; Altelaar, M.; de Vries, R.P. Cultivation of Podospora anserina on soybean hulls results in an efficient enzyme cocktail for plant biomass hydrolysis. New Biotechnol. 2017, 37, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2015, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Sloothaak, J.; Odoni, D.I.; de Graaff, L.H.; Martins Dos Santos, V.A.P.; Schaap, P.J.; Tamayo-Ramos, J.A. Aspergillus niger membrane-associated proteome analysis for the identification of glucose transporters. Biotechnol. Biofuels 2015, 8, 150. [Google Scholar] [CrossRef] [PubMed]
- Sloothaak, J.; Tamayo-Ramos, J.A.; Odoni, D.I.; Laothanachareon, T.; Derntl, C.; Mach-Aigner, A.R.; Martins Dos Santos, V.A.P.; Schaap, P.J. Identification and functional characterization of novel xylose transporters from the cell factories Aspergillus niger and Trichoderma reesei. Biotechnol. Biofuels 2016, 9, 148. [Google Scholar] [CrossRef] [PubMed]
- vanKuyk, P.A.; Diderich, J.A.; MacCabe, A.P.; Hererro, O.; Ruijter, G.J.G.; Visser, J. Aspergillus niger mstA encodes a high-affinity sugar/H+ symporter which is regulated in response to extracellular pH. Biochem. J. 2004, 379, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Ozturkoglu Budak, S.; Zhou, M.; Brouwer, C.; Wiebenga, A.; Benoit, I.; Di Falco, M.; Tsang, A.; de Vries, R.P. A genomic survey of proteases in Aspergilli. BMC Genom. 2014, 15, 523. [Google Scholar] [CrossRef]
- Yoshino, K.; Yamamoto, K.; Hara, K.; Sonoda, M.; Yamamoto, Y.; Sakamoto, K. The conservation of polyol transporter proteins and their involvement in lichenized Ascomycota. Fungal Biol. 2019, 123, 318–329. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]





| Feedstock | Rha | Ara | Xyl | Man | Gal | Glc | UA | Total | Polysaccharides |
|---|---|---|---|---|---|---|---|---|---|
| Wheat bran | 0.0 | 16.5 | 34.6 | 1.4 | 1.7 | 42.5 | 3.3 | 53.7 | cellulose, arabinoxylan |
| Sugar beet pulp | 1.5 | 29.0 | 2.4 | 2.1 | 6.5 | 32.0 | 27.0 | 56.0 | cellulose, pectin, xyloglucan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, J.; Mäkelä, M.R.; de Vries, R.P. Identification of an l-Arabitol Transporter from Aspergillus niger. Biomolecules 2023, 13, 188. https://doi.org/10.3390/biom13020188
Meng J, Mäkelä MR, de Vries RP. Identification of an l-Arabitol Transporter from Aspergillus niger. Biomolecules. 2023; 13(2):188. https://doi.org/10.3390/biom13020188
Chicago/Turabian StyleMeng, Jiali, Miia R. Mäkelä, and Ronald P. de Vries. 2023. "Identification of an l-Arabitol Transporter from Aspergillus niger" Biomolecules 13, no. 2: 188. https://doi.org/10.3390/biom13020188
APA StyleMeng, J., Mäkelä, M. R., & de Vries, R. P. (2023). Identification of an l-Arabitol Transporter from Aspergillus niger. Biomolecules, 13(2), 188. https://doi.org/10.3390/biom13020188