
On the Prevalence and Roles of Proteins Undergoing Liquid–Liquid Phase Separation in the Biogenesis of PML-Bodies

 , ,
, ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
- -
- Viral DNA-containing PML bodies;
- -
- Giant PML bodies containing satellite DNA;
- -
- Alternative lengthening of telomeres (ALT)-associated PML bodies.
2. Materials and Methods
2.1. Study Design
- (i).
- An analysis of the PML-body proteome and the PML protein interactome in the BIOGRID (v.4.4) databases https://thebiogrid.org/ (accessed on 10 October 2023), and resources https://amigo.geneontology.org/amigo/term/GO:0016605 (accessed on 10 October 2023) (GO:0016605) (PML NB/ND10) and SL-0465 (https://www.uniprot.org/locations/SL-0465 (accessed on 10 October 2023);
- (ii).
- An analysis of data from the literature.
2.2. Intrinsic Disorder Prediction
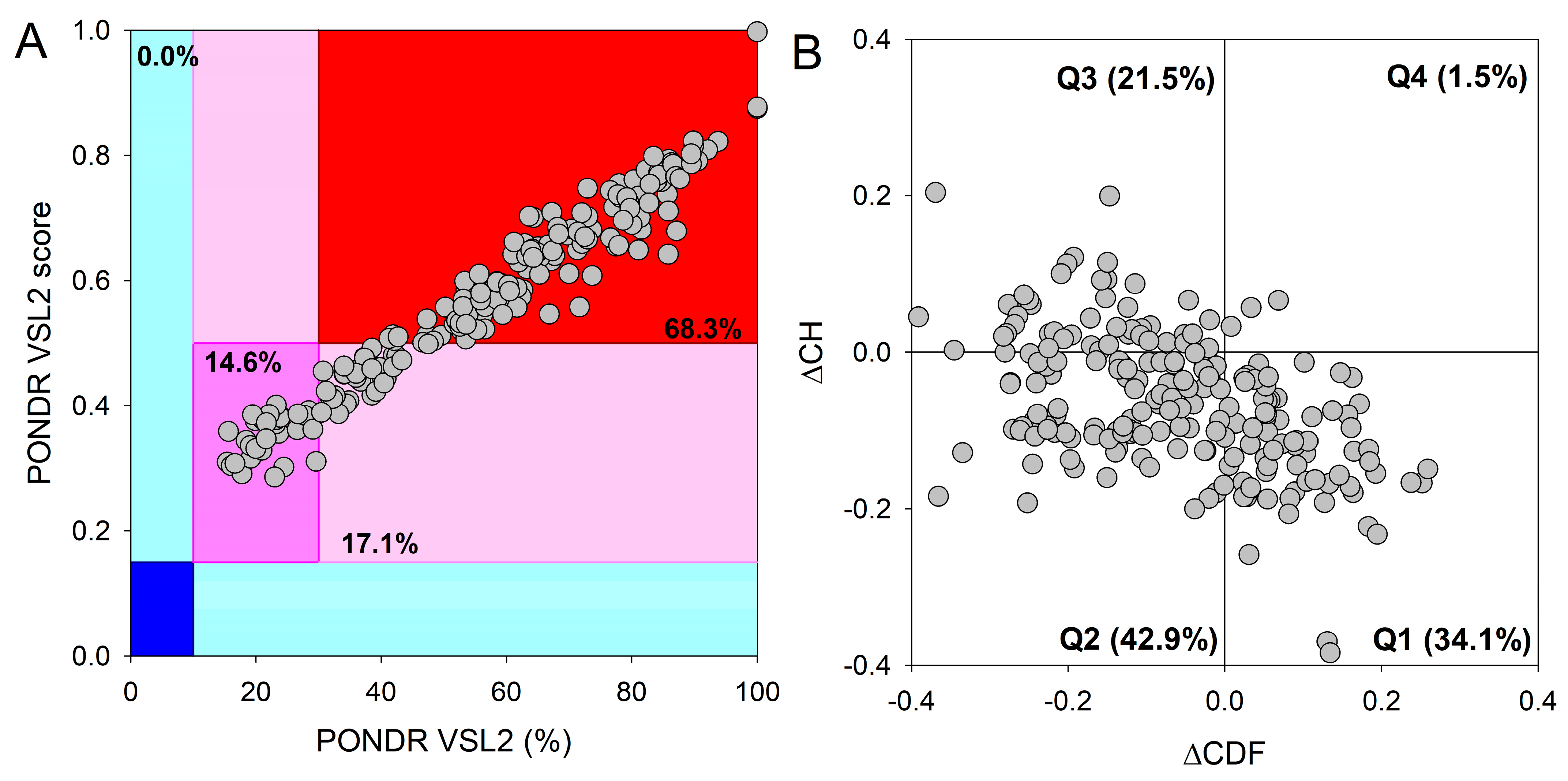
2.3. LLPS Predisposition Analysis
2.4. Evaluation of Protein Charge and Hydrophobicity
2.5. Determination of Proteins Targeted for SUMOylation
2.6. Aggregation Propensity Prediction
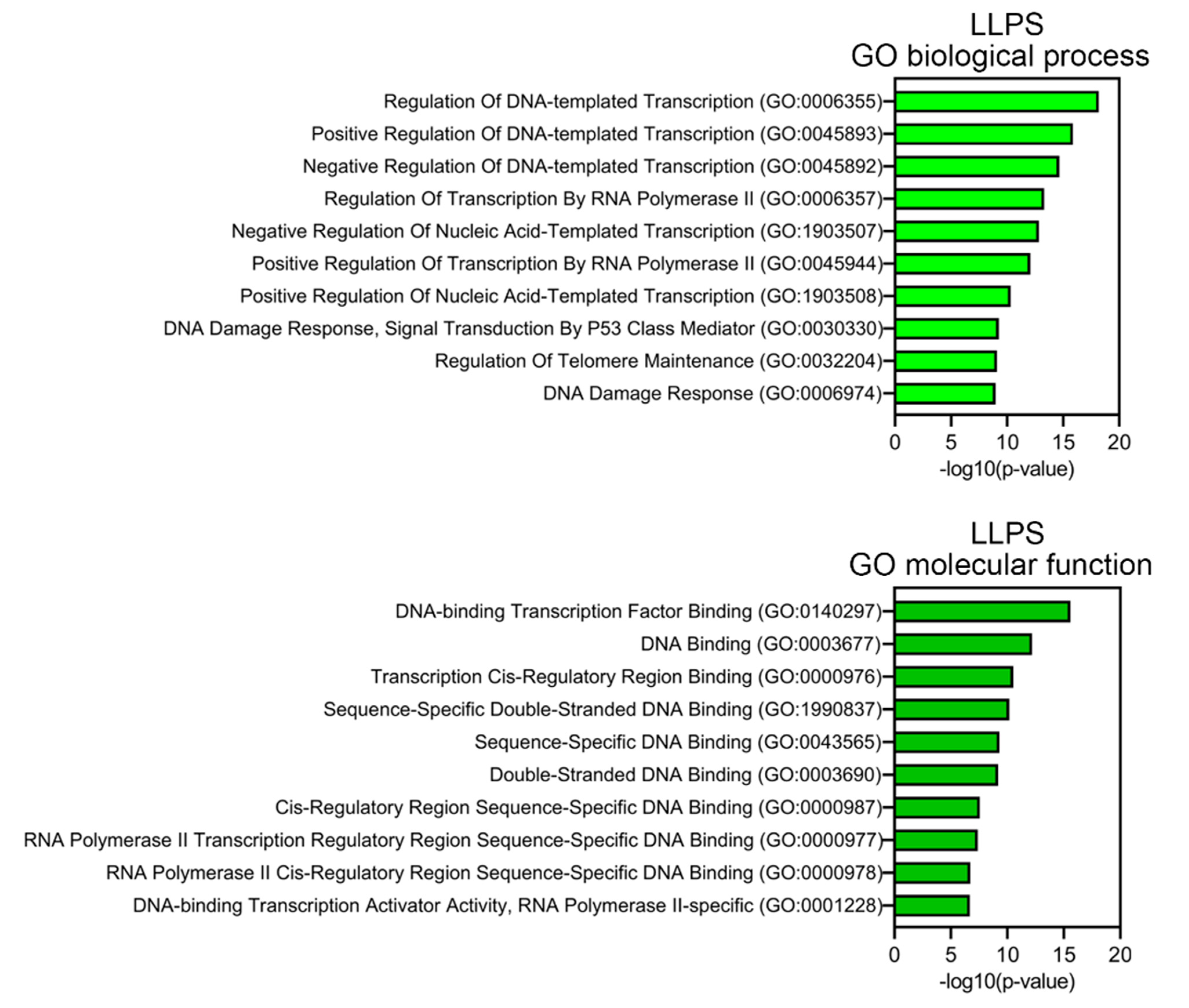
2.7. Determination of the Biological Processes and Molecular Functions of Proteins
3. Results and Discussion
3.1. Finding Proteins Included in the PML-Body Proteome
3.2. PML Bodies’ Scaffold Proteins
3.3. Proteome Analysis of PML Bodies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, B.; Zhang, L.; Dai, T.; Qin, Z.; Lu, H.; Zhang, L.; Zhou, F. Liquid-liquid phase separation in human health and diseases. Signal Transduct. Target. Ther. 2021, 6, 290. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Brangwynne, C.P. Liquid phase condensation in cell physiology and disease. Science 2017, 357, eaaf4382. [Google Scholar] [CrossRef] [PubMed]
- Antifeeva, I.A.; Fonin, A.V.; Fefilova, A.S.; Stepanenko, O.V.; Povarova, O.I.; Silonov, S.A.; Kuznetsova, I.M.; Uversky, V.N.; Turoverov, K.K. Liquid-liquid phase separation as an organizing principle of intracellular space: Overview of the evolution of the cell compartmentalization concept. Cell. Mol. Life Sci. 2022, 79, 251. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.; Weber, S.C.; Vaidya, N.; Haataja, M.; Brangwynne, C.P. RNA transcription modulates phase transition-driven nuclear body assembly. Proc. Natl. Acad. Sci. USA 2015, 112, E5237–E5245. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, D.; Battich, N.; Pelkmans, L. A Systems-Level Study Reveals Regulators of Membrane-less Organelles in Human Cells. Mol. Cell 2018, 72, 1035–1049.e5. [Google Scholar] [CrossRef]
- Brangwynne, C.P.; Tompa, P.; Pappu, R.V. Polymer physics of intracellular phase transitions. Nat. Phys. 2015, 11, 899–904. [Google Scholar] [CrossRef]
- Dignon, G.L.; Best, R.B.; Mittal, J. Biomolecular Phase Separation: From Molecular Driving Forces to Macroscopic Properties. Annu. Rev. Phys. Chem. 2020, 71, 53–75. [Google Scholar] [CrossRef]
- Adame-Arana, O.; Weber, C.A.; Zaburdaev, V.; Prost, J.; Jülicher, F. Liquid Phase Separation Controlled by pH. Biophys. J. 2020, 119, 1590–1605. [Google Scholar] [CrossRef]
- Corpet, A.; Kleijwegt, C.; Roubille, S.; Juillard, F.; Jacquet, K.; Texier, P.; Lomonte, P. PML nuclear bodies and chromatin dynamics: Catch me if you can! Nucleic Acids Res. 2020, 48, 11890–11912. [Google Scholar] [CrossRef]
- Niwa-Kawakita, M.; Wu, H.C.; Thé, H.; Lallemand-Breitenbach, V. PML nuclear bodies, membrane-less domains acting as ROS sensors? Semin. Cell Dev. Biol. 2018, 80, 29–34. [Google Scholar] [CrossRef]
- Lallemand-Breitenbach, V. PML nuclear bodies. Cold Spring Harb. Perspect. Biol. 2010, 2, a000661. [Google Scholar] [CrossRef] [PubMed]
- Prikrylova, T.; Pachernik, J.; Kozubek, S.; Bartova, E. Epigenetics and chromatin plasticity in embryonic stem cells. World J. Stem Cells 2013, 5, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, F.M.; Hendzel, M.J.; Bazett-Jones, D.P. Promyelocytic leukemia (PML) nuclear bodies are protein structures that do not accumulate RNA. J. Cell Biol. 2000, 148, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Hands, K.J.; Cuchet-Lourenco, D.; Everett, R.D.; Hay, R.T. PML isoforms in response to arsenic: High-resolution analysis of PML body structure and degradation characteristics. J. Cell Sci. 2014, 127, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Guan, D.; Kao, H.Y. The function, regulation and therapeutic implications of the tumor suppressor protein, PML. Cell Biosci. 2015, 5, 60. [Google Scholar] [CrossRef] [PubMed]
- Fonin, A.V.; Silonov, S.A.; Shpironok, O.G.; Antifeeva, I.A.; Petukhov, A.V.; Romanovich, A.E.; Kuznetsova, I.M.; Uversky, V.N.; Turoverov, K.K. The Role of Non-Specific Interactions in Canonical and ALT-Associated PML-Bodies Formation and Dynamics. Int. J. Mol. Sci. 2021, 22, 5821. [Google Scholar] [CrossRef]
- Li, Y.; Ma, X.; Wu, W.; Chen, Z.; Meng, G. PML Nuclear Body Biogenesis, Carcinogenesis, and Targeted Therapy. Trends Cancer 2020, 6, 889–906. [Google Scholar] [CrossRef]
- Sahin, U.; De Thé, H.; Lallemand-Breitenbach, V. PML nuclear bodies: Assembly and oxidative stress-sensitive sumoylation. Nucleus 2014, 5, 499–507. [Google Scholar] [CrossRef]
- Fonin, A.V.; Silonov, S.A.; Fefilova, A.S.; Stepanenko, O.V.; Gavrilova, A.A.; Petukhov, A.V.; Romanovich, A.E.; Modina, A.L.; Zueva, T.S.; Nedelyaev, E.M.; et al. New Evidence of the Importance of Weak Interactions in the Formation of PML-Bodies. Int. J. Mol. Sci. 2022, 23, 1613. [Google Scholar] [CrossRef]
- Dayhoff, G.W., 2nd; Uversky, V.N. Rapid prediction and analysis of protein intrinsic disorder. Protein Sci. 2022, 31, e4496. [Google Scholar] [CrossRef]
- Meszaros, B.; Erdos, G.; Dosztanyi, Z. IUPred2A: Context-dependent prediction of protein disorder as a function of redox state and protein binding. Nucleic Acids Res. 2018, 46, W329–W337. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, Z.; Peng, K.; Vucetic, S.; Radivojac, P.; Dunker, A.K. Exploiting heterogeneous sequence properties improves prediction of protein disorder. Proteins Struct. Funct. Bioinform. 2005, 61, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Radivojac, P.; Vucetic, S.; Dunker, A.K.; Obradovic, Z. Length-dependent prediction of protein intrinsic disorder. BMC Bioinform. 2006, 7, 208. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Vucetic, S.; Radivojac, P.; Brown, C.J.; Dunker, A.K.; Obradovic, Z. Optimizing long intrinsic disorder predictors with protein evolutionary information. J. Bioinform. Comput. Biol. 2005, 3, 35–60. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.; Obradovic, Z.; Li, X.; Garner, E.C.; Brown, C.J.; Dunker, A.K. Sequence complexity of disordered protein. Proteins 2001, 42, 38–48. [Google Scholar] [CrossRef]
- Xue, B.; Dunbrack, R.L.; Williams, R.W.; Dunker, A.K.; Uversky, V.N. PONDR-FIT: A meta-predictor of intrinsically disordered amino acids. Biochim. Biophys. Acta 2010, 1804, 996–1010. [Google Scholar] [CrossRef]
- Rajagopalan, K.; Mooney, S.M.; Parekh, N.; Getzenberg, R.H.; Kulkarni, P. A majority of the cancer/testis antigens are intrinsically disordered proteins. J. Cell. Biochem. 2011, 112, 3256–3267. [Google Scholar] [CrossRef]
- Uversky, V.N. Analyzing IDPs in interactomes. In Intrinsically Disordered Proteins; Kragelund, B.B., Skriver, K., Eds.; Volume Methods in Molecular Biology; Humana: New York, NY, USA, 2020; pp. 895–945. [Google Scholar]
- Uversky, V.N.; Gillespie, J.R.; Fink, A.L. Why are “natively unfolded” proteins unstructured under physiologic conditions? Proteins 2000, 41, 415–427. [Google Scholar] [CrossRef]
- Oldfield, C.J.; Cheng, Y.; Cortese, M.S.; Brown, C.J.; Uversky, V.N.; Dunker, A.K. Comparing and combining predictors of mostly disordered proteins. Biochemistry 2005, 44, 1989–2000. [Google Scholar] [CrossRef]
- Mohan, A.; Sullivan, W.J., Jr.; Radivojac, P.; Dunker, A.K.; Uversky, V.N. Intrinsic disorder in pathogenic and non-pathogenic microbes: Discovering and analyzing the unfoldomes of early-branching eukaryotes. Mol. Biosyst. 2008, 4, 328–340. [Google Scholar] [CrossRef]
- Huang, F.; Oldfield, C.; Meng, J.; Hsu, W.L.; Xue, B.; Uversky, V.N.; Romero, P.; Dunker, A.K. Subclassifying disordered proteins by the CH-CDF plot method. In Proceedings of the Pacific Symposium on Biocomputing, Kohala Coast, HI, USA, 3–7 January 2012; pp. 128–139. [Google Scholar]
- Hardenberg, M.; Horvath, A.; Ambrus, V.; Fuxreiter, M.; Vendruscolo, M. Widespread occurrence of the droplet state of proteins in the human proteome. Proc. Natl. Acad. Sci. USA 2020, 117, 33254–33262. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Sun, T.; Li, Q.; Xu, Y.; Zhang, Z.; Lai, L.; Pei, J. Prediction of liquid-liquid phase separating proteins using machine learning. BMC Bioinform. 2022, 23, 72. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Conchillo-Solé, O.; de Groot, N.S.; Avilés, F.X.; Vendrell, J.; Daura, X.; Ventura, S. AGGRESCAN: A server for the prediction and evaluation of “hot spots” of aggregation in polypeptides. BMC Bioinform. 2007, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Guan, D.; Lim, J.H.; Peng, L.; Liu, Y.; Lam, M.; Seto, E.; Kao, H.Y. Deacetylation of the tumor suppressor protein PML regulates hydrogen peroxide-induced cell death. Cell Death Dis. 2014, 5, e1340. [Google Scholar] [CrossRef]
- Lång, A.; Lång, E.; Bøe, S.O. PML Bodies in Mitosis. Cells 2019, 8, 893. [Google Scholar] [CrossRef]
- Missiroli, S.; Bonora, M.; Patergnani, S.; Poletti, F.; Perrone, M.; Gafà, R.; Magri, E.; Raimondi, A.; Lanza, G.; Tacchetti, C.; et al. PML at Mitochondria-Associated Membranes Is Critical for the Repression of Autophagy and Cancer Development. Cell Rep. 2016, 16, 2415–2427. [Google Scholar] [CrossRef]
- Giorgi, C.; Ito, K.; Lin, H.K.; Santangelo, C.; Wieckowski, M.R.; Lebiedzinska, M.; Bononi, A.; Bonora, M.; Duszynski, J.; Bernardi, R.; et al. PML regulates apoptosis at endoplasmic reticulum by modulating calcium release. Science 2010, 330, 1247–1251. [Google Scholar] [CrossRef]
- Cheng, X.; Kao, H.Y. Post-translational modifications of PML: Consequences and implications. Front. Oncol. 2012, 2, 210. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.; Laukens, K.; Dang, T.H.; Van Ostade, X. A manually curated network of the PML nuclear body interactome reveals an important role for PML-NBs in SUMOylation dynamics. Int. J. Biol. Sci. 2010, 6, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Gao, Y.; Lin, H.K. Cytoplasmic PML: From molecular regulation to biological functions. J. Cell. Biochem. 2014, 115, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.H.; Lamark, T.; Isakson, P.; Finley, K.; Larsen, K.B.; Brech, A.; Øvervatn, A.; Stenmark, H.; Bjørkøy, G.; Simonsen, A.; et al. p62/SQSTM1 and ALFY interact to facilitate the formation of p62 bodies/ALIS and their degradation by autophagy. Autophagy 2010, 6, 330–344. [Google Scholar] [CrossRef] [PubMed]
- Korioth, F.; Gieffers, C.; Maul, G.G.; Frey, J. Molecular characterization of NDP52, a novel protein of the nuclear domain 10, which is redistributed upon virus infection and interferon treatment. J. Cell Biol. 1995, 130, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ahmady, E.; Deeke, S.A.; Rabaa, S.; Kouri, L.; Kenney, L.; Stewart, A.F.; Burgon, P.G. Identification of a novel muscle A-type lamin-interacting protein (MLIP). J. Biol. Chem. 2011, 286, 19702–19713. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Luo, Z.; Jiang, S.; Li, F.; Han, X.; Hu, Y.; Wang, D.; Zhao, Y.; Ma, W.; Liu, D.; et al. The telomere-associated homeobox-containing protein TAH1/HMBOX1 participates in telomere maintenance in ALT cells. J. Cell Sci. 2013, 126, 3982–3989. [Google Scholar] [CrossRef]
- Lopez, P.; Vidal, F.; Martin, L.; Lopez-Fernandez, L.A.; Rual, J.F.; Rosen, B.S.; Cuzin, F.; Rassoulzadegan, M. Gene control in germinal differentiation: RNF6, a transcription regulatory protein in the mouse sertoli cell. Mol. Cell. Biol. 2002, 22, 3488–3496. [Google Scholar] [CrossRef]
- Cheung, B.B.; Bell, J.; Raif, A.; Bohlken, A.; Yan, J.; Roediger, B.; Poljak, A.; Smith, S.; Lee, M.; Thomas, W.D.; et al. The estrogen-responsive B box protein is a novel regulator of the retinoid signal. J. Biol. Chem. 2006, 281, 18246–18256. [Google Scholar] [CrossRef]
- Gao, C.; Cheng, X.; Lam, M.; Liu, Y.; Liu, Q.; Chang, K.S.; Kao, H.Y. Signal-dependent regulation of transcription by histone deacetylase 7 involves recruitment to promyelocytic leukemia protein nuclear bodies. Mol. Biol. Cell 2008, 19, 3020–3027. [Google Scholar] [CrossRef][Green Version]
- Green, L.M.; Wagner, K.J.; Campbell, H.A.; Addison, K.; Roberts, S.G. Dynamic interaction between WT1 and BASP1 in transcriptional regulation during differentiation. Nucleic Acids Res. 2009, 37, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.C.; Lee, C.C.; Yao, Y.L.; Lai, C.C.; Schmitz, M.L.; Yang, W.M. SUMO5, a Novel Poly-SUMO Isoform, Regulates PML Nuclear Bodies. Sci. Rep. 2016, 6, 26509. [Google Scholar] [CrossRef] [PubMed]
- Tse, W.T.; Tang, J.; Jin, O.; Korsgren, C.; John, K.M.; Kung, A.L.; Gwynn, B.; Peters, L.L.; Lux, S.E. A new spectrin, beta IV, has a major truncated isoform that associates with promyelocytic leukemia protein nuclear bodies and the nuclear matrix. J. Biol. Chem. 2001, 276, 23974–23985. [Google Scholar] [CrossRef] [PubMed]
- Roussigne, M.; Cayrol, C.; Clouaire, T.; Amalric, F.; Girard, J.P. THAP1 is a nuclear proapoptotic factor that links prostate-apoptosis-response-4 (Par-4) to PML nuclear bodies. Oncogene 2003, 22, 2432–2442. [Google Scholar] [CrossRef]
- Boisvert, F.M.; Hendzel, M.J.; Masson, J.Y.; Richard, S. Methylation of MRE11 regulates its nuclear compartmentalization. Cell Cycle 2005, 4, 981–989. [Google Scholar] [CrossRef]
- Lee, J.; Lee, Y.; Lee, M.J.; Park, E.; Kang, S.H.; Chung, C.H.; Lee, K.H.; Kim, K. Dual modification of BMAL1 by SUMO2/3 and ubiquitin promotes circadian activation of the CLOCK/BMAL1 complex. Mol. Cell. Biol. 2008, 28, 6056–6065. [Google Scholar] [CrossRef]
- Karvonen, U.; Jääskeläinen, T.; Rytinki, M.; Kaikkonen, S.; Palvimo, J.J. ZNF451 is a novel PML body- and SUMO-associated transcriptional coregulator. J. Mol. Biol. 2008, 382, 585–600. [Google Scholar] [CrossRef]
- Yang, S.; Kuo, C.; Bisi, J.E.; Kim, M.K. PML-dependent apoptosis after DNA damage is regulated by the checkpoint kinase hCds1/Chk2. Nat. Cell Biol. 2002, 4, 865–870. [Google Scholar] [CrossRef]
- Sano, M.; Tokudome, S.; Shimizu, N.; Yoshikawa, N.; Ogawa, C.; Shirakawa, K.; Endo, J.; Katayama, T.; Yuasa, S.; Ieda, M.; et al. Intramolecular control of protein stability, subnuclear compartmentalization, and coactivator function of peroxisome proliferator-activated receptor gamma coactivator 1alpha. J. Biol. Chem. 2007, 282, 25970–25980. [Google Scholar] [CrossRef]
- Miki, T.; Xu, Z.; Chen-Goodspeed, M.; Liu, M.; Van Oort-Jansen, A.; Rea, M.A.; Zhao, Z.; Lee, C.C.; Chang, K.S. PML regulates PER2 nuclear localization and circadian function. EMBO J. 2012, 31, 1427–1439. [Google Scholar] [CrossRef]
- Li, Q.; He, Y.; Wei, L.; Wu, X.; Wu, D.; Lin, S.; Wang, Z.; Ye, Z.; Lin, S.C. AXIN is an essential co-activator for the promyelocytic leukemia protein in p53 activation. Oncogene 2011, 30, 1194–1204. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kaytor, M.D.; Duvick, L.A.; Skinner, P.J.; Koob, M.D.; Ranum, L.P.; Orr, H.T. Nuclear localization of the spinocerebellar ataxia type 7 protein, ataxin-7. Hum. Mol. Genet. 1999, 8, 1657–1664. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jang, S.M.; Nathans, J.F.; Fu, H.; Redon, C.E.; Jenkins, L.M.; Thakur, B.L.; Pongor, L.S.; Baris, A.M.; Gross, J.M.; O’Neill, M.J.; et al. The RepID-CRL4 ubiquitin ligase complex regulates metaphase to anaphase transition via BUB3 degradation. Nat. Commun. 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Marcello, A.; Ferrari, A.; Pellegrini, V.; Pegoraro, G.; Lusic, M.; Beltram, F.; Giacca, M. Recruitment of human cyclin T1 to nuclear bodies through direct interaction with the PML protein. EMBO J. 2003, 22, 2156–2166. [Google Scholar] [CrossRef]
- Wu, G.; Lee, W.H.; Chen, P.L. NBS1 and TRF1 colocalize at promyelocytic leukemia bodies during late S/G2 phases in immortalized telomerase-negative cells. Implication of NBS1 in alternative lengthening of telomeres. J. Biol. Chem. 2000, 275, 30618–30622. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Nomura, T.; Kim, H.; Kaul, S.C.; Wadhwa, R.; Shinagawa, T.; Ichikawa-Iwata, E.; Zhong, S.; Pandolfi, P.P.; Ishii, S. Role of PML and PML-RARalpha in Mad-mediated transcriptional repression. Mol. Cell 2001, 7, 1233–1243. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Conn, K.L.; Wasson, P.; Charman, M.; Tong, L.; Grant, K.; McFarlane, S.; Boutell, C. SUMO Ligase Protein Inhibitor of Activated STAT1 (PIAS1) Is a Constituent Promyelocytic Leukemia Nuclear Body Protein That Contributes to the Intrinsic Antiviral Immune Response to Herpes Simplex Virus 1. J. Virol. 2016, 90, 5939–5952. [Google Scholar] [CrossRef]
- Li, C.; McManus, F.P.; Plutoni, C.; Pascariu, C.M.; Nelson, T.; Alberici Delsin, L.E.; Emery, G.; Thibault, P. Quantitative SUMO proteomics identifies PIAS1 substrates involved in cell migration and motility. Nat. Commun. 2020, 11, 834. [Google Scholar] [CrossRef]
- Xu, G.L.; Pan, Y.K.; Wang, B.Y.; Huang, L.; Tian, L.; Xue, J.L.; Chen, J.Z.; Jia, W. TTRAP is a novel PML nuclear bodies-associated protein. Biochem. Biophys. Res. Commun. 2008, 375, 395–398. [Google Scholar] [CrossRef]
- Vilotti, S.; Biagioli, M.; Foti, R.; Dal Ferro, M.; Lavina, Z.S.; Collavin, L.; Del Sal, G.; Zucchelli, S.; Gustincich, S. The PML nuclear bodies-associated protein TTRAP regulates ribosome biogenesis in nucleolar cavities upon proteasome inhibition. Cell Death Differ. 2012, 19, 488–500. [Google Scholar] [CrossRef]
- Nayak, A.; Glöckner-Pagel, J.; Vaeth, M.; Schumann, J.E.; Buttmann, M.; Bopp, T.; Schmitt, E.; Serfling, E.; Berberich-Siebelt, F. Sumoylation of the transcription factor NFATc1 leads to its subnuclear relocalization and interleukin-2 repression by histone deacetylase. J. Biol. Chem. 2009, 284, 10935–10946. [Google Scholar] [CrossRef]
- Cairo, S.; De Falco, F.; Pizzo, M.; Salomoni, P.; Pandolfi, P.P.; Meroni, G. PML interacts with Myc, and Myc target gene expression is altered in PML-null fibroblasts. Oncogene 2005, 24, 2195–2203. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cobb, A.M.; De Silva, S.A.; Hayward, R.; Sek, K.; Ulferts, S.; Grosse, R.; Shanahan, C.M. Filamentous nuclear actin regulation of PML NBs during the DNA damage response is deregulated by prelamin A. Cell Death Dis. 2022, 13, 1042. [Google Scholar] [CrossRef] [PubMed]
- Fogal, V.; Gostissa, M.; Sandy, P.; Zacchi, P.; Sternsdorf, T.; Jensen, K.; Pandolfi, P.P.; Will, H.; Schneider, C.; Del Sal, G. Regulation of p53 activity in nuclear bodies by a specific PML isoform. EMBO J. 2000, 19, 6185–6195. [Google Scholar] [CrossRef] [PubMed]
- Marks, D.; Heinen, N.; Bachmann, L.; Meermeyer, S.; Werner, M.; Gallego, L.; Hemmerich, P.; Bader, V.; Winklhofer, K.F.; Schröder, E.; et al. Amyloid precursor protein elevates fusion of promyelocytic leukemia nuclear bodies in human hippocampal areas with high plaque load. Acta Neuropathol. Commun. 2021, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Alcalay, M.; Tomassoni, L.; Colombo, E.; Stoldt, S.; Grignani, F.; Fagioli, M.; Szekely, L.; Helin, K.; Pelicci, P.G. The promyelocytic leukemia gene product (PML) forms stable complexes with the retinoblastoma protein. Mol. Cell. Biol. 1998, 18, 1084–1093. [Google Scholar] [CrossRef]
- Cohen, N.; Sharma, M.; Kentsis, A.; Perez, J.M.; Strudwick, S.; Borden, K.L. PML RING suppresses oncogenic transformation by reducing the affinity of eIF4E for mRNA. EMBO J. 2001, 20, 4547–4559. [Google Scholar] [CrossRef]
- Kentsis, A.; Dwyer, E.C.; Perez, J.M.; Sharma, M.; Chen, A.; Pan, Z.Q.; Borden, K.L. The RING domains of the promyelocytic leukemia protein PML and the arenaviral protein Z repress translation by directly inhibiting translation initiation factor eIF4E. J. Mol. Biol. 2001, 312, 609–623. [Google Scholar] [CrossRef]
- Dahle, Ø.; Bakke, O.; Gabrielsen, O.S. c-Myb associates with PML in nuclear bodies in hematopoietic cells. Exp. Cell Res. 2004, 297, 118–126. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Minami, T.; Tojo, M.; Honda, Y.; Uchimura, Y.; Saitoh, H.; Yasuda, H.; Nagahiro, S.; Saya, H.; Nakao, M. Serum response factor is modulated by the SUMO-1 conjugation system. Biochem. Biophys. Res. Commun. 2003, 306, 32–38. [Google Scholar] [CrossRef]
- Gupta, P.; Ho, P.C.; Ha, S.G.; Lin, Y.W.; Wei, L.N. HDAC3 as a molecular chaperone for shuttling phosphorylated TR2 to PML: A novel deacetylase activity-independent function of HDAC3. PLoS ONE 2009, 4, e4363. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Duprez, E.; Borden, K.L.; Freemont, P.S.; Etkin, L.D. Ret finger protein is a normal component of PML nuclear bodies and interacts directly with PML. J. Cell Sci. 1998, 111 Pt 10, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.S.; Xu, Z.X.; Ran, R.; Meng, F.; Chang, K.S. Promyelocytic leukemia protein PML inhibits Nur77-mediated transcription through specific functional interactions. Oncogene 2002, 21, 3925–3933. [Google Scholar] [CrossRef] [PubMed]
- Dent, A.L.; Yewdell, J.; Puvion-Dutilleul, F.; Koken, M.H.; de The, H.; Staudt, L.M. LYSP100-associated nuclear domains (LANDs): Description of a new class of subnuclear structures and their relationship to PML nuclear bodies. Blood 1996, 88, 1423–1426. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.T.; Tajsic, T.; Mellad, J.A.; Searles, R.; Zhang, Q.; Shanahan, C.M. Novel nuclear nesprin-2 variants tether active extracellular signal-regulated MAPK1 and MAPK2 at promyelocytic leukemia protein nuclear bodies and act to regulate smooth muscle cell proliferation. J. Biol. Chem. 2010, 285, 1311–1320. [Google Scholar] [CrossRef]
- Ulbricht, T.; Alzrigat, M.; Horch, A.; Reuter, N.; von Mikecz, A.; Steimle, V.; Schmitt, E.; Krämer, O.H.; Stamminger, T.; Hemmerich, P. PML promotes MHC class II gene expression by stabilizing the class II transactivator. J. Cell Biol. 2012, 199, 49–63. [Google Scholar] [CrossRef]
- Dhordain, P.; Albagli, O.; Honore, N.; Guidez, F.; Lantoine, D.; Schmid, M.; The, H.D.; Zelent, A.; Koken, M.H. Colocalization and heteromerization between the two human oncogene POZ/zinc finger proteins, LAZ3 (BCL6) and PLZF. Oncogene 2000, 19, 6240–6250. [Google Scholar] [CrossRef]
- Buchberger, E.; El Harchi, M.; Payrhuber, D.; Zommer, A.; Schauer, D.; Simonitsch-Klupp, I.; Bilban, M.; Brostjan, C. Overexpression of the transcriptional repressor complex BCL-6/BCoR leads to nuclear aggregates distinct from classical aggresomes. PLoS ONE 2013, 8, e76845. [Google Scholar] [CrossRef]
- Peche, L.Y.; Scolz, M.; Ladelfa, M.F.; Monte, M.; Schneider, C. MageA2 restrains cellular senescence by targeting the function of PMLIV/p53 axis at the PML-NBs. Cell Death Differ. 2012, 19, 926–936. [Google Scholar] [CrossRef]
- Johnson, F.B.; Lombard, D.B.; Neff, N.F.; Mastrangelo, M.A.; Dewolf, W.; Ellis, N.A.; Marciniak, R.A.; Yin, Y.; Jaenisch, R.; Guarente, L. Association of the Bloom syndrome protein with topoisomerase IIIalpha in somatic and meiotic cells. Cancer Res. 2000, 60, 1162–1167. [Google Scholar]
- Kruse, M.L.; Arlt, A.; Sieke, A.; Grohmann, F.; Grossmann, M.; Minkenberg, J.; Fölsch, U.R.; Schäfer, H. Immediate early gene X1 (IEX-1) is organized in subnuclear structures and partially co-localizes with promyelocytic leukemia protein in HeLa cells. J. Biol. Chem. 2005, 280, 24849–24856. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.; Adamovich, Y.; Reuven, N.; Shaul, Y. The Yes-associated protein 1 stabilizes p73 by preventing Itch-mediated ubiquitination of p73. Cell Death Differ. 2007, 14, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Strano, S.; Monti, O.; Pediconi, N.; Baccarini, A.; Fontemaggi, G.; Lapi, E.; Mantovani, F.; Damalas, A.; Citro, G.; Sacchi, A.; et al. The transcriptional coactivator Yes-associated protein drives p73 gene-target specificity in response to DNA Damage. Mol. Cell 2005, 18, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Basei, F.L.; Meirelles, G.V.; Righetto, G.L.; Dos Santos Migueleti, D.L.; Smetana, J.H.; Kobarg, J. New interaction partners for Nek4.1 and Nek4.2 isoforms: From the DNA damage response to RNA splicing. Proteome Sci. 2015, 13, 11. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.M.; Proytcheva, M.; Ellis, N.A.; Holloman, W.K.; German, J. BLM, the Bloom’s syndrome protein, varies during the cell cycle in its amount, distribution, and co-localization with other nuclear proteins. Cytogenet. Cell Genet. 2000, 91, 217–223. [Google Scholar] [CrossRef]
- Luciani, J.J.; Depetris, D.; Usson, Y.; Metzler-Guillemain, C.; Mignon-Ravix, C.; Mitchell, M.J.; Megarbane, A.; Sarda, P.; Sirma, H.; Moncla, A.; et al. PML nuclear bodies are highly organised DNA-protein structures with a function in heterochromatin remodelling at the G2 phase. J. Cell Sci. 2006, 119, 2518–2531. [Google Scholar] [CrossRef]
- Zhang, Y.; Shao, A.W.; Tang, J.; Geng, Y. PML-II recruits ataxin-3 to PML-NBs and inhibits its deubiquitinating activity. Biochem. Biophys. Res. Commun. 2021, 554, 186–192. [Google Scholar] [CrossRef]
- Fu, C.; Ahmed, K.; Ding, H.; Ding, X.; Lan, J.; Yang, Z.; Miao, Y.; Zhu, Y.; Shi, Y.; Zhu, J.; et al. Stabilization of PML nuclear localization by conjugation and oligomerization of SUMO-3. Oncogene 2005, 24, 5401–5413. [Google Scholar] [CrossRef]
- Rabellino, A.; Carter, B.; Konstantinidou, G.; Wu, S.Y.; Rimessi, A.; Byers, L.A.; Heymach, J.V.; Girard, L.; Chiang, C.M.; Teruya-Feldstein, J.; et al. The SUMO E3-ligase PIAS1 regulates the tumor suppressor PML and its oncogenic counterpart PML-RARA. Cancer Res. 2012, 72, 2275–2284. [Google Scholar] [CrossRef]
- Duprez, E.; Saurin, A.J.; Desterro, J.M.; Lallemand-Breitenbach, V.; Howe, K.; Boddy, M.N.; Solomon, E.; de Thé, H.; Hay, R.T.; Freemont, P.S. SUMO-1 modification of the acute promyelocytic leukaemia protein PML: Implications for nuclear localisation. J. Cell Sci. 1999, 112 Pt 3, 381–393. [Google Scholar] [CrossRef]
- Barroso-Gomila, O.; Trulsson, F.; Muratore, V.; Canosa, I.; Merino-Cacho, L.; Cortazar, A.R.; Pérez, C.; Azkargorta, M.; Iloro, I.; Carracedo, A.; et al. Identification of proximal SUMO-dependent interactors using SUMO-ID. Nat. Commun. 2021, 12, 6671. [Google Scholar] [CrossRef] [PubMed]
- Häkli, M.; Karvonen, U.; Jänne, O.A.; Palvimo, J.J. SUMO-1 promotes association of SNURF (RNF4) with PML nuclear bodies. Exp. Cell Res. 2005, 304, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Shastrula, P.K.; Sierra, I.; Deng, Z.; Keeney, F.; Hayden, J.E.; Lieberman, P.M.; Janicki, S.M. PML is recruited to heterochromatin during S phase and represses DAXX-mediated histone H3.3 chromatin assembly. J. Cell Sci. 2019, 132, jcs.220970. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Rogers, R.; Matunis, M.J.; Mayhew, C.N.; Goodson, M.L.; Park-Sarge, O.K.; Sarge, K.D. Regulation of heat shock transcription factor 1 by stress-induced SUMO-1 modification. J. Biol. Chem. 2001, 276, 40263–40267. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.S.; Xu, Z.X.; Hittelman, W.N.; Salomoni, P.; Pandolfi, P.P.; Chang, K.S. Promyelocytic leukemia protein sensitizes tumor necrosis factor alpha-induced apoptosis by inhibiting the NF-kappaB survival pathway. J. Biol. Chem. 2003, 278, 12294–12304. [Google Scholar] [CrossRef] [PubMed]
- Giogha, C.; Lung, T.W.; Mühlen, S.; Pearson, J.S.; Hartland, E.L. Substrate recognition by the zinc metalloprotease effector NleC from enteropathogenic Escherichia coli. Cell. Microbiol. 2015, 17, 1766–1778. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, A.; Wier, E.M.; Fu, K.; Sun, X.; Yu, H.; Zheng, W.; Sham, H.P.; Johnson, K.; Bailey, S.; Vallance, B.A.; et al. Metalloprotease NleC suppresses host NF-κB/inflammatory responses by cleaving p65 and interfering with the p65/RPS3 interaction. PLoS Pathog. 2015, 11, e1004705. [Google Scholar] [CrossRef]
- Koken, M.H.; Reid, A.; Quignon, F.; Chelbi-Alix, M.K.; Davies, J.M.; Kabarowski, J.H.; Zhu, J.; Dong, S.; Chen, S.; Chen, Z.; et al. Leukemia-associated retinoic acid receptor alpha fusion partners, PML and PLZF, heterodimerize and colocalize to nuclear bodies. Proc. Natl. Acad. Sci. USA 1997, 94, 10255–10260. [Google Scholar] [CrossRef]
- Xu, D.; Holko, M.; Sadler, A.J.; Scott, B.; Higashiyama, S.; Berkofsky-Fessler, W.; McConnell, M.J.; Pandolfi, P.P.; Licht, J.D.; Williams, B.R. Promyelocytic leukemia zinc finger protein regulates interferon-mediated innate immunity. Immunity 2009, 30, 802–816. [Google Scholar] [CrossRef]
- Wang, X.; He, S.; Sun, J.M.; Delcuve, G.P.; Davie, J.R. Selective association of peroxiredoxin 1 with genomic DNA and COX-2 upstream promoter elements in estrogen receptor negative breast cancer cells. Mol. Biol. Cell 2010, 21, 2987–2995. [Google Scholar] [CrossRef]
- Sachini, N.; Arampatzi, P.; Klonizakis, A.; Nikolaou, C.; Makatounakis, T.; Lam, E.W.; Kretsovali, A.; Papamatheakis, J. Promyelocytic leukemia protein (PML) controls breast cancer cell proliferation by modulating Forkhead transcription factors. Mol. Oncol. 2019, 13, 1369–1387. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liao, L.; Deng, X.; Chen, R.; Gray, N.S.; Yates, J.R., 3rd; Lee, J.D. BMK1 is involved in the regulation of p53 through disrupting the PML-MDM2 interaction. Oncogene 2013, 32, 3156–3164. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yoshida, N.; Murakami, N.; Kawata, K.; Ishizaki, H.; Tanaka-Okamoto, M.; Miyoshi, J.; Zinn, A.R.; Shime, H.; Inoue, N. Dynamic regulation of p53 subnuclear localization and senescence by MORC3. Mol. Biol. Cell 2007, 18, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Sloan, E.; Orr, A.; Everett, R.D. MORC3, a Component of PML Nuclear Bodies, Has a Role in Restricting Herpes Simplex Virus 1 and Human Cytomegalovirus. J. Virol. 2016, 90, 8621–8633. [Google Scholar] [CrossRef] [PubMed]
- Mimura, Y.; Takahashi, K.; Kawata, K.; Akazawa, T.; Inoue, N. Two-step colocalization of MORC3 with PML nuclear bodies. J. Cell Sci. 2010, 123, 2014–2024. [Google Scholar] [CrossRef] [PubMed]
- Renner, F.; Moreno, R.; Schmitz, M.L. SUMOylation-dependent localization of IKKepsilon in PML nuclear bodies is essential for protection against DNA-damage-triggered cell death. Mol. Cell 2010, 37, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Park, J.S.; Kang, Y.K. Dual functions of histone-lysine N-methyltransferase Setdb1 protein at promyelocytic leukemia-nuclear body (PML-NB): Maintaining PML-NB structure and regulating the expression of its associated genes. J. Biol. Chem. 2011, 286, 41115–41124. [Google Scholar] [CrossRef]
- Schmidt, J.A.; Danielson, K.G.; Duffner, E.R.; Radecki, S.G.; Walker, G.T.; Shelton, A.; Wang, T.; Knepper, J.E. Regulation of the oncogenic phenotype by the nuclear body protein ZC3H8. BMC Cancer 2018, 18, 759. [Google Scholar] [CrossRef]
- Kleijwegt, C.; Bressac, F.; Seurre, C.; Bouchereau, W.; Cohen, C.; Texier, P.; Simonet, T.; Schaeffer, L.; Lomonte, P.; Corpet, A. Interplay between PML NBs and HIRA for H3.3 dynamics following type I interferon stimulus. eLife 2023, 12, e80156. [Google Scholar] [CrossRef]
- Potts, P.R.; Yu, H. The SMC5/6 complex maintains telomere length in ALT cancer cells through SUMOylation of telomere-binding proteins. Nat. Struct. Mol. Biol. 2007, 14, 581–590. [Google Scholar] [CrossRef]
- Brouwer, A.K.; Schimmel, J.; Wiegant, J.C.; Vertegaal, A.C.; Tanke, H.J.; Dirks, R.W. Telomeric DNA mediates de novo PML body formation. Mol. Biol. Cell 2009, 20, 4804–4815. [Google Scholar] [CrossRef] [PubMed]
- González-Prieto, R.; Cuijpers, S.A.; Luijsterburg, M.S.; van Attikum, H.; Vertegaal, A.C. SUMOylation and PARylation cooperate to recruit and stabilize SLX4 at DNA damage sites. EMBO Rep. 2015, 16, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Forlani, G.; Tosi, G.; Turrini, F.; Poli, G.; Vicenzi, E.; Accolla, R.S. Tripartite Motif-Containing Protein 22 Interacts with Class II Transactivator and Orchestrates Its Recruitment in Nuclear Bodies Containing TRIM19/PML and Cyclin T1. Front. Immunol. 2017, 8, 564. [Google Scholar] [CrossRef] [PubMed]
- Ivanschitz, L.; Takahashi, Y.; Jollivet, F.; Ayrault, O.; Le Bras, M.; de Thé, H. PML IV/ARF interaction enhances p53 SUMO-1 conjugation, activation, and senescence. Proc. Natl. Acad. Sci. USA 2015, 112, 14278–14283. [Google Scholar] [CrossRef] [PubMed]
- Oravcová, M.; Nie, M.; Zilio, N.; Maeda, S.; Jami-Alahmadi, Y.; Lazzerini-Denchi, E.; Wohlschlegel, J.A.; Ulrich, H.D.; Otomo, T.; Boddy, M.N. The Nse5/6-like SIMC1-SLF2 complex localizes SMC5/6 to viral replication centers. eLife 2022, 11, e79676. [Google Scholar] [CrossRef]
- Shiio, Y.; Rose, D.W.; Aur, R.; Donohoe, S.; Aebersold, R.; Eisenman, R.N. Identification and characterization of SAP25, a novel component of the mSin3 corepressor complex. Mol. Cell. Biol. 2006, 26, 1386–1397. [Google Scholar] [CrossRef]
- Pearson, M.; Carbone, R.; Sebastiani, C.; Cioce, M.; Fagioli, M.; Saito, S.; Higashimoto, Y.; Appella, E.; Minucci, S.; Pandolfi, P.P.; et al. PML regulates p53 acetylation and premature senescence induced by oncogenic Ras. Nature 2000, 406, 207–210. [Google Scholar] [CrossRef]
- Doucas, V. The promyelocytic (PML) nuclear compartment and transcription control. Biochem. Pharmacol. 2000, 60, 1197–1201. [Google Scholar] [CrossRef]
- Wu, Q.; Hu, H.; Lan, J.; Emenari, C.; Wang, Z.; Chang, K.S.; Huang, H.; Yao, X. PML3 Orchestrates the Nuclear Dynamics and Function of TIP60. J. Biol. Chem. 2009, 284, 8747–8759. [Google Scholar] [CrossRef]
- Everett, R.D.; Meredith, M.; Orr, A.; Cross, A.; Kathoria, M.; Parkinson, J. A novel ubiquitin-specific protease is dynamically associated with the PML nuclear domain and binds to a herpesvirus regulatory protein. EMBO J. 1997, 16, 1519–1530. [Google Scholar] [CrossRef]
- Song, M.S.; Salmena, L.; Carracedo, A.; Egia, A.; Lo-Coco, F.; Teruya-Feldstein, J.; Pandolfi, P.P. The deubiquitinylation and localization of PTEN are regulated by a HAUSP-PML network. Nature 2008, 455, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Langley, E.; Pearson, M.; Faretta, M.; Bauer, U.M.; Frye, R.A.; Minucci, S.; Pelicci, P.G.; Kouzarides, T. Human SIR2 deacetylates p53 and antagonizes PML/p53-induced cellular senescence. EMBO J. 2002, 21, 2383–2396. [Google Scholar] [CrossRef] [PubMed]
- Hattersley, N.; Shen, L.; Jaffray, E.G.; Hay, R.T. The SUMO protease SENP6 is a direct regulator of PML nuclear bodies. Mol. Biol. Cell 2011, 22, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Tatemichi, Y.; Shibazaki, M.; Yasuhira, S.; Kasai, S.; Tada, H.; Oikawa, H.; Suzuki, Y.; Takikawa, Y.; Masuda, T.; Maesawa, C. Nucleus accumbens associated 1 is recruited within the promyelocytic leukemia nuclear body through SUMO modification. Cancer Sci. 2015, 106, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wang, F.; Cao, W.B.; Lv, X.X.; Hua, F.; Cui, B.; Yu, J.J.; Zhang, X.W.; Shang, S.; Liu, S.S.; et al. TRIB3 Promotes APL Progression through Stabilization of the Oncoprotein PML-RARα and Inhibition of p53-Mediated Senescence. Cancer Cell 2017, 31, 697–710.e7. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Livingston, C.M.; Li, L.; Beran, R.K.; Daffis, S.; Ramakrishnan, D.; Burdette, D.; Peiser, L.; Salas, E.; Ramos, H.; et al. The Smc5/6 Complex Restricts HBV when Localized to ND10 without Inducing an Innate Immune Response and Is Counteracted by the HBV X Protein Shortly after Infection. PLoS ONE 2017, 12, e0169648. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Park, Y.J.; Hwang, I.Y.; Youdim, M.B.; Park, K.S.; Oh, Y.J. Nuclear translocation of DJ-1 during oxidative stress-induced neuronal cell death. Free Radic. Biol. Med. 2012, 53, 936–950. [Google Scholar] [CrossRef]
- Suico, M.A.; Yoshida, H.; Seki, Y.; Uchikawa, T.; Lu, Z.; Shuto, T.; Matsuzaki, K.; Nakao, M.; Li, J.D.; Kai, H. Myeloid Elf-1-like factor, an ETS transcription factor, up-regulates lysozyme transcription in epithelial cells through interaction with promyelocytic leukemia protein. J. Biol. Chem. 2004, 279, 19091–19098. [Google Scholar] [CrossRef]
- Fukuyo, Y.; Mogi, K.; Tsunematsu, Y.; Nakajima, T. E2FBP1/hDril1 modulates cell growth through downregulation of promyelocytic leukemia bodies. Cell Death Differ. 2004, 11, 747–759. [Google Scholar] [CrossRef]
- Tatematsu, K.; Yoshimoto, N.; Koyanagi, T.; Tokunaga, C.; Tachibana, T.; Yoneda, Y.; Yoshida, M.; Okajima, T.; Tanizawa, K.; Kuroda, S. Nuclear-cytoplasmic shuttling of a RING-IBR protein RBCK1 and its functional interaction with nuclear body proteins. J. Biol. Chem. 2005, 280, 22937–22944. [Google Scholar] [CrossRef]
- He, W.; Hu, C.X.; Hou, J.K.; Fan, L.; Xu, Y.W.; Liu, M.H.; Yan, S.Y.; Chen, G.Q.; Huang, Y. Microtubule-associated protein 1 light chain 3 interacts with and contributes to growth inhibiting effect of PML. PLoS ONE 2014, 9, e113089. [Google Scholar] [CrossRef] [PubMed]
- Harhouri, K.; Navarro, C.; Depetris, D.; Mattei, M.G.; Nissan, X.; Cau, P.; De Sandre-Giovannoli, A.; Lévy, N. MG132-induced progerin clearance is mediated by autophagy activation and splicing regulation. EMBO Mol. Med. 2017, 9, 1294–1313. [Google Scholar] [CrossRef] [PubMed]
- Zannini, L.; Buscemi, G.; Fontanella, E.; Lisanti, S.; Delia, D. REGgamma/PA28gamma proteasome activator interacts with PML and Chk2 and affects PML nuclear bodies number. Cell Cycle 2009, 8, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, B.C.; Kim, S.J.; Choi, K.S. Role of MAP kinases and their cross-talk in TGF-beta1-induced apoptosis in FaO rat hepatoma cell line. Hepatology 2002, 35, 1360–1371. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.G.; Möller, A.; Sirma, H.; Zentgraf, H.; Taya, Y.; Dröge, W.; Will, H.; Schmitz, M.L. Regulation of p53 activity by its interaction with homeodomain-interacting protein kinase-2. Nat. Cell Biol. 2002, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Banumathy, G.; Somaiah, N.; Zhang, R.; Tang, Y.; Hoffmann, J.; Andrake, M.; Ceulemans, H.; Schultz, D.; Marmorstein, R.; Adams, P.D. Human UBN1 is an ortholog of yeast Hpc2p and has an essential role in the HIRA/ASF1a chromatin-remodeling pathway in senescent cells. Mol. Cell. Biol. 2009, 29, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Chakarova, C.F.; Papaioannou, M.G.; Khanna, H.; Lopez, I.; Waseem, N.; Shah, A.; Theis, T.; Friedman, J.; Maubaret, C.; Bujakowska, K.; et al. Mutations in TOPORS cause autosomal dominant retinitis pigmentosa with perivascular retinal pigment epithelium atrophy. Am. J. Hum. Genet. 2007, 81, 1098–1103. [Google Scholar] [CrossRef]
- Ji, L.; Huo, X.; Zhang, Y.; Yan, Z.; Wang, Q.; Wen, B. TOPORS, a tumor suppressor protein, contributes to the maintenance of higher-order chromatin architecture. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194518. [Google Scholar] [CrossRef]
- Hariharasudhan, G.; Jeong, S.Y.; Kim, M.J.; Jung, S.M.; Seo, G.; Moon, J.R.; Lee, S.; Chang, I.Y.; Kee, Y.; You, H.J.; et al. TOPORS-mediated RAD51 SUMOylation facilitates homologous recombination repair. Nucleic Acids Res. 2022, 50, 1501–1516. [Google Scholar] [CrossRef]
- Salsman, J.; Pinder, J.; Tse, B.; Corkery, D.; Dellaire, G. The translation initiation factor 3 subunit eIF3K interacts with PML and associates with PML nuclear bodies. Exp. Cell Res. 2013, 319, 2554–2565. [Google Scholar] [CrossRef]
- Kunapuli, P.; Kasyapa, C.S.; Chin, S.F.; Caldas, C.; Cowell, J.K. ZNF198, a zinc finger protein rearranged in myeloproliferative disease, localizes to the PML nuclear bodies and interacts with SUMO-1 and PML. Exp. Cell Res. 2006, 312, 3739–3751. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y.; Shima, T.; Chiba, T.; Irimura, T.; Pandolfi, P.P.; Kitabayashi, I. PML activates transcription by protecting HIPK2 and p300 from SCFFbx3-mediated degradation. Mol. Cell. Biol. 2008, 28, 7126–7138. [Google Scholar] [CrossRef]
- Milovic-Holm, K.; Krieghoff, E.; Jensen, K.; Will, H.; Hofmann, T.G. FLASH links the CD95 signaling pathway to the cell nucleus and nuclear bodies. EMBO J. 2007, 26, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Chen, Y.; Yan, K.; Shao, Y.; Zhang, Q.C.; Lin, Y.; Xi, Q. Recruitment of TRIM33 to cell-context specific PML nuclear bodies regulates nodal signaling in mESCs. EMBO J. 2023, 42, e112058. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.Q.; Nguyen, A.; Cao, Y.; Chang, A.C.; Reddel, R.R. HP1-mediated formation of alternative lengthening of telomeres-associated PML bodies requires HIRA but not ASF1a. PLoS ONE 2011, 6, e17036. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Block, S.; Scribano, C.M.; Thiry, R.; Esbona, K.; Audhya, A.; Weaver, B.A. Mad1 destabilizes p53 by preventing PML from sequestering MDM2. Nat. Commun. 2019, 10, 1540. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Seo, T.; Kim, H.; Choe, J. Sumoylation of the novel protein hRIP{beta} is involved in replication protein A deposition in PML nuclear bodies. Mol. Cell. Biol. 2005, 25, 8202–8214. [Google Scholar] [CrossRef]
- Sohn, S.Y.; Hearing, P. Adenoviral strategies to overcome innate cellular responses to infection. FEBS Lett. 2019, 593, 3484–3495. [Google Scholar] [CrossRef] [PubMed]
- Barr, S.M.; Leung, C.G.; Chang, E.E.; Cimprich, K.A. ATR kinase activity regulates the intranuclear translocation of ATR and RPA following ionizing radiation. Curr. Biol. 2003, 13, 1047–1051. [Google Scholar] [CrossRef]
- Bloch, D.B.; Chiche, J.D.; Orth, D.; de la Monte, S.M.; Rosenzweig, A.; Bloch, K.D. Structural and functional heterogeneity of nuclear bodies. Mol. Cell. Biol. 1999, 19, 4423–4430. [Google Scholar] [CrossRef]
- Bloch, D.B.; de la Monte, S.M.; Guigaouri, P.; Filippov, A.; Bloch, K.D. Identification and characterization of a leukocyte-specific component of the nuclear body. J. Biol. Chem. 1996, 271, 29198–29204. [Google Scholar] [CrossRef] [PubMed]
- Bloch, D.B.; Nakajima, A.; Gulick, T.; Chiche, J.D.; Orth, D.; de La Monte, S.M.; Bloch, K.D. Sp110 localizes to the PML-Sp100 nuclear body and may function as a nuclear hormone receptor transcriptional coactivator. Mol. Cell. Biol. 2000, 20, 6138–6146. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chu, J.; Yucer, N.; Leng, M.; Wang, S.Y.; Chen, B.P.; Hittelman, W.N.; Wang, Y. RING finger and WD repeat domain 3 (RFWD3) associates with replication protein A (RPA) and facilitates RPA-mediated DNA damage response. J. Biol. Chem. 2011, 286, 22314–22322. [Google Scholar] [CrossRef] [PubMed]
- Lunardi, A.; Chiacchiera, F.; D’Este, E.; Carotti, M.; Dal Ferro, M.; Di Minin, G.; Del Sal, G.; Collavin, L. The evolutionary conserved gene C16orf35 encodes a nucleo-cytoplasmic protein that interacts with p73. Biochem. Biophys. Res. Commun. 2009, 388, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Deiss, K.; Lockwood, N.; Howell, M.; Segeren, H.A.; Saunders, R.E.; Chakravarty, P.; Soliman, T.N.; Martini, S.; Rocha, N.; Semple, R.; et al. A genome-wide RNAi screen identifies the SMC5/6 complex as a non-redundant regulator of a Topo2a-dependent G2 arrest. Nucleic Acids Res. 2019, 47, 2906–2921. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, M.; Gong, R.; Wang, Z.; Lu, L.; Peng, S.; Duan, Z.; Feng, Y.; Liu, Y.; Gao, B. Forkhead O Transcription Factor 4 Restricts HBV Covalently Closed Circular DNA Transcription and HBV Replication through Genetic Downregulation of Hepatocyte Nuclear Factor 4 Alpha and Epigenetic Suppression of Covalently Closed Circular DNA via Interacting with Promyelocytic Leukemia Protein. J. Virol. 2022, 96, e0054622. [Google Scholar] [CrossRef] [PubMed]
- Borden, K.L.; Campbelldwyer, E.J.; Carlile, G.W.; Djavani, M.; Salvato, M.S. Two RING finger proteins, the oncoprotein PML and the arenavirus Z protein, colocalize with the nuclear fraction of the ribosomal P proteins. J. Virol. 1998, 72, 3819–3826. [Google Scholar] [CrossRef]
- Bernassola, F.; Salomoni, P.; Oberst, A.; Di Como, C.J.; Pagano, M.; Melino, G.; Pandolfi, P.P. Ubiquitin-dependent degradation of p73 is inhibited by PML. J. Exp. Med. 2004, 199, 1545–1557. [Google Scholar] [CrossRef]
- Lee, Y.R.; Yuan, W.C.; Ho, H.C.; Chen, C.H.; Shih, H.M.; Chen, R.H. The Cullin 3 substrate adaptor KLHL20 mediates DAPK ubiquitination to control interferon responses. EMBO J. 2010, 29, 1748–1761. [Google Scholar] [CrossRef]
- Martin, N.; Benhamed, M.; Nacerddine, K.; Demarque, M.D.; van Lohuizen, M.; Dejean, A.; Bischof, O. Physical and functional interaction between PML and TBX2 in the establishment of cellular senescence. EMBO J. 2012, 31, 95–109. [Google Scholar] [CrossRef]
- Oh, W.; Ghim, J.; Lee, E.W.; Yang, M.R.; Kim, E.T.; Ahn, J.H.; Song, J. PML-IV functions as a negative regulator of telomerase by interacting with TERT. J. Cell Sci. 2009, 122, 2613–2622. [Google Scholar] [CrossRef] [PubMed]
- Salomoni, P.; Bernardi, R.; Bergmann, S.; Changou, A.; Tuttle, S.; Pandolfi, P.P. The promyelocytic leukemia protein PML regulates c-Jun function in response to DNA damage. Blood 2005, 105, 3686–3690. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, J.; Zou, W.X.; Chang, K.S. Inhibition of Sp1 functions by its sequestration into PML nuclear bodies. PLoS ONE 2014, 9, e94450. [Google Scholar] [CrossRef] [PubMed]
- Kordon, M.M.; Szczurek, A.; Berniak, K.; Szelest, O.; Solarczyk, K.; Tworzydło, M.; Wachsmann-Hogiu, S.; Vaahtokari, A.; Cremer, C.; Pederson, T.; et al. PML-like subnuclear bodies, containing XRCC1, juxtaposed to DNA replication-based single-strand breaks. FASEB J. 2019, 33, 2301–2313. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, L.Q.; Yu, W.; Zhao, Z.G.; Xie, X.M.; Wang, W.T.; Xiong, J.; Li, M.; Xue, Z.; Wang, X.; et al. PML4 facilitates erythroid differentiation by enhancing the transcriptional activity of GATA-1. Blood 2014, 123, 261–270. [Google Scholar] [CrossRef]
- Rao, V.A.; Fan, A.M.; Meng, L.; Doe, C.F.; North, P.S.; Hickson, I.D.; Pommier, Y. Phosphorylation of BLM, dissociation from topoisomerase IIIalpha, and colocalization with gamma-H2AX after topoisomerase I-induced replication damage. Mol. Cell. Biol. 2005, 25, 8925–8937. [Google Scholar] [CrossRef]
- Carbone, R.; Pearson, M.; Minucci, S.; Pelicci, P.G. PML NBs associate with the hMre11 complex and p53 at sites of irradiation induced DNA damage. Oncogene 2002, 21, 1633–1640. [Google Scholar] [CrossRef]
- Gibb, S.L.; Boston-Howes, W.; Lavina, Z.S.; Gustincich, S.; Brown, R.H., Jr.; Pasinelli, P.; Trotti, D. A caspase-3-cleaved fragment of the glial glutamate transporter EAAT2 is sumoylated and targeted to promyelocytic leukemia nuclear bodies in mutant SOD1-linked amyotrophic lateral sclerosis. J. Biol. Chem. 2007, 282, 32480–32490. [Google Scholar] [CrossRef]
- Alsheich-Bartok, O.; Haupt, S.; Alkalay-Snir, I.; Saito, S.; Appella, E.; Haupt, Y. PML enhances the regulation of p53 by CK1 in response to DNA damage. Oncogene 2008, 27, 3653–3661. [Google Scholar] [CrossRef]
- Gialitakis, M.; Arampatzi, P.; Makatounakis, T.; Papamatheakis, J. Gamma interferon-dependent transcriptional memory via relocalization of a gene locus to PML nuclear bodies. Mol. Cell. Biol. 2010, 30, 2046–2056. [Google Scholar] [CrossRef]
- Kurki, S.; Latonen, L.; Laiho, M. Cellular stress and DNA damage invoke temporally distinct Mdm2, p53 and PML complexes and damage-specific nuclear relocalization. J. Cell Sci. 2003, 116, 3917–3925. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.A.; Sun, Y.; Song, J.; Chen, Y.; Krontiris, T.G.; Durrin, L.K. SUMO conjugation to the matrix attachment region-binding protein, special AT-rich sequence-binding protein-1 (SATB1), targets SATB1 to promyelocytic nuclear bodies where it undergoes caspase cleavage. J. Biol. Chem. 2008, 283, 18124–18134. [Google Scholar] [CrossRef] [PubMed]
- Sapetschnig, A.; Rischitor, G.; Braun, H.; Doll, A.; Schergaut, M.; Melchior, F.; Suske, G. Transcription factor Sp3 is silenced through SUMO modification by PIAS1. EMBO J. 2002, 21, 5206–5215. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.; Best, J.L.; Zon, L.I.; Gill, G. SUMO-1 modification represses Sp3 transcriptional activation and modulates its subnuclear localization. Mol. Cell 2002, 10, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.J.; Meloni, A.R.; Nevins, J.R. The Rb-related p130 protein controls telomere lengthening through an interaction with a Rad50-interacting protein, RINT-1. Mol. Cell 2006, 22, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.I.; Kitamura, T.; Kruse, J.P.; Raum, J.C.; Stein, R.; Gu, W.; Accili, D. FoxO1 protects against pancreatic beta cell failure through NeuroD and MafA induction. Cell Metab. 2005, 2, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Voisset, E.; Moravcsik, E.; Stratford, E.W.; Jaye, A.; Palgrave, C.J.; Hills, R.K.; Salomoni, P.; Kogan, S.C.; Solomon, E.; Grimwade, D. Pml nuclear body disruption cooperates in APL pathogenesis and impairs DNA damage repair pathways in mice. Blood 2018, 131, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Aster, J.C.; Blacklow, S.C.; Lake, R.; Artavanis-Tsakonas, S.; Griffin, J.D. MAML1, a human homologue of Drosophila mastermind, is a transcriptional co-activator for NOTCH receptors. Nat. Genet. 2000, 26, 484–489. [Google Scholar] [CrossRef]
- Takahashi, H.; Hatakeyama, S.; Saitoh, H.; Nakayama, K.I. Noncovalent SUMO-1 binding activity of thymine DNA glycosylase (TDG) is required for its SUMO-1 modification and colocalization with the promyelocytic leukemia protein. J. Biol. Chem. 2005, 280, 5611–5621. [Google Scholar] [CrossRef]
- Blander, G.; Zalle, N.; Daniely, Y.; Taplick, J.; Gray, M.D.; Oren, M. DNA damage-induced translocation of the Werner helicase is regulated by acetylation. J. Biol. Chem. 2002, 277, 50934–50940. [Google Scholar] [CrossRef]
- El-Asmi, F.; El-Mchichi, B.; Maroui, M.A.; Dianoux, L.; Chelbi-Alix, M.K. TGF-β induces PML SUMOylation, degradation and PML nuclear body disruption. Cytokine 2019, 120, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Osterwald, S.; Deeg, K.I.; Chung, I.; Parisotto, D.; Wörz, S.; Rohr, K.; Erfle, H.; Rippe, K. PML induces compaction, TRF2 depletion and DNA damage signaling at telomeres and promotes their alternative lengthening. J. Cell Sci. 2015, 128, 1887–1900. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Park, B.; Cho, S.; Lee, S.; Kim, Y.; Lee, S.O.; Cho, K.; Lee, S.; Jin, B.S.; Ahn, J.H.; et al. Promyelocytic leukemia is a direct inhibitor of SAPK2/p38 mitogen-activated protein kinase. J. Biol. Chem. 2004, 279, 40994–41003. [Google Scholar] [CrossRef] [PubMed]
- Merkl, P.E.; Orzalli, M.H.; Knipe, D.M. Mechanisms of Host IFI16, PML, and Daxx Protein Restriction of Herpes Simplex Virus 1 Replication. J. Virol. 2018, 92, e00057-18. [Google Scholar] [CrossRef] [PubMed]
- Sasai, N.; Saitoh, N.; Saitoh, H.; Nakao, M. The transcriptional cofactor MCAF1/ATF7IP is involved in histone gene expression and cellular senescence. PLoS ONE 2013, 8, e68478. [Google Scholar] [CrossRef]
- Sun, H.; Liu, Y.; Hunter, T. Multiple Arkadia/RNF111 structures coordinate its Polycomb body association and transcriptional control. Mol. Cell. Biol. 2014, 34, 2981–2995. [Google Scholar] [CrossRef]
- Schmitz, M.L.; Grishina, I. Regulation of the tumor suppressor PML by sequential post-translational modifications. Front. Oncol. 2012, 2, 204. [Google Scholar] [CrossRef][Green Version]
- Rauth, S.; Karmakar, S.; Shah, A.; Seshacharyulu, P.; Nimmakayala, R.K.; Ganguly, K.; Bhatia, R.; Muniyan, S.; Kumar, S.; Dutta, S.; et al. SUMO Modification of PAF1/PD2 Enables PML Interaction and Promotes Radiation Resistance in Pancreatic Ductal Adenocarcinoma. Mol. Cell. Biol. 2021, 41, e0013521. [Google Scholar] [CrossRef]
- Xu, Z.X.; Timanova-Atanasova, A.; Zhao, R.X.; Chang, K.S. PML colocalizes with and stabilizes the DNA damage response protein TopBP1. Mol. Cell. Biol. 2003, 23, 4247–4256. [Google Scholar] [CrossRef]
- Rokudai, S.; Laptenko, O.; Arnal, S.M.; Taya, Y.; Kitabayashi, I.; Prives, C. MOZ increases p53 acetylation and premature senescence through its complex formation with PML. Proc. Natl. Acad. Sci. USA 2013, 110, 3895–3900. [Google Scholar] [CrossRef]
- Guan, D.; Factor, D.; Liu, Y.; Wang, Z.; Kao, H.Y. The epigenetic regulator UHRF1 promotes ubiquitination-mediated degradation of the tumor-suppressor protein promyelocytic leukemia protein. Oncogene 2013, 32, 3819–3828. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, J.; Dong, F.N.; Dong, J.T. Characterization of nuclear localization and SUMOylation of the ATBF1 transcription factor in epithelial cells. PLoS ONE 2014, 9, e92746. [Google Scholar] [CrossRef] [PubMed]
- Georges, A.; Benayoun, B.A.; Marongiu, M.; Dipietromaria, A.; L’Hôte, D.; Todeschini, A.L.; Auer, J.; Crisponi, L.; Veitia, R.A. SUMOylation of the Forkhead transcription factor FOXL2 promotes its stabilization/activation through transient recruitment to PML bodies. PLoS ONE 2011, 6, e25463. [Google Scholar] [CrossRef] [PubMed]
- Shire, K.; Wong, A.I.; Tatham, M.H.; Anderson, O.F.; Ripsman, D.; Gulstene, S.; Moffat, J.; Hay, R.T.; Frappier, L. Identification of RNF168 as a PML nuclear body regulator. J. Cell Sci. 2016, 129, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Imrichova, T.; Hubackova, S.; Kucerova, A.; Kosla, J.; Bartek, J.; Hodny, Z.; Vasicova, P. Dynamic PML protein nucleolar associations with persistent DNA damage lesions in response to nucleolar stress and senescence-inducing stimuli. Aging 2019, 11, 7206–7235. [Google Scholar] [CrossRef] [PubMed]
- Lång, A.; Eriksson, J.; Schink, K.O.; Lång, E.; Blicher, P.; Połeć, A.; Brech, A.; Dalhus, B.; Bøe, S.O. Visualization of PML nuclear import complexes reveals FG-repeat nucleoporins at cargo retrieval sites. Nucleus 2017, 8, 404–420. [Google Scholar] [CrossRef] [PubMed]
- Lallemand-Breitenbach, V.; Zhu, J.; Puvion, F.; Koken, M.; Honoré, N.; Doubeikovsky, A.; Duprez, E.; Pandolfi, P.P.; Puvion, E.; Freemont, P.; et al. Role of promyelocytic leukemia (PML) sumolation in nuclear body formation, 11S proteasome recruitment, and As2O3-induced PML or PML/retinoic acid receptor alpha degradation. J. Exp. Med. 2001, 193, 1361–1371. [Google Scholar] [CrossRef]
- Tang, J.; Xie, W.; Yang, X. Association of caspase-2 with the promyelocytic leukemia protein nuclear bodies. Cancer Biol. Ther. 2005, 4, 645–649. [Google Scholar] [CrossRef]
- Scheper, G.C.; Parra, J.L.; Wilson, M.; Van Kollenburg, B.; Vertegaal, A.C.; Han, Z.G.; Proud, C.G. The N and C termini of the splice variants of the human mitogen-activated protein kinase-interacting kinase Mnk2 determine activity and localization. Mol. Cell. Biol. 2003, 23, 5692–5705. [Google Scholar] [CrossRef]
- Sabò, A.; Lusic, M.; Cereseto, A.; Giacca, M. Acetylation of conserved lysines in the catalytic core of cyclin-dependent kinase 9 inhibits kinase activity and regulates transcription. Mol. Cell. Biol. 2008, 28, 2201–2212. [Google Scholar] [CrossRef]
- Tomasini, R.; Samir, A.A.; Carrier, A.; Isnardon, D.; Cecchinelli, B.; Soddu, S.; Malissen, B.; Dagorn, J.C.; Iovanna, J.L.; Dusetti, N.J. TP53INP1s and homeodomain-interacting protein kinase-2 (HIPK2) are partners in regulating p53 activity. J. Biol. Chem. 2003, 278, 37722–37729. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Kim, D.U.; Kehrl, J.H. RGS14 is a centrosomal and nuclear cytoplasmic shuttling protein that traffics to promyelocytic leukemia nuclear bodies following heat shock. J. Biol. Chem. 2005, 280, 805–814. [Google Scholar] [CrossRef]
- Messaoudi, L.; Yang, Y.G.; Kinomura, A.; Stavreva, D.A.; Yan, G.; Bortolin-Cavaillé, M.L.; Arakawa, H.; Buerstedde, J.M.; Hainaut, P.; Cavaillé, J.; et al. Subcellular distribution of human RDM1 protein isoforms and their nucleolar accumulation in response to heat shock and proteotoxic stress. Nucleic Acids Res. 2007, 35, 6571–6587. [Google Scholar] [CrossRef] [PubMed]
- Gongora, C.; David, G.; Pintard, L.; Tissot, C.; Hua, T.D.; Dejean, A.; Mechti, N. Molecular cloning of a new interferon-induced PML nuclear body-associated protein. J. Biol. Chem. 1997, 272, 19457–19463. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S.; Reed, J.C. ZIP kinase triggers apoptosis from nuclear PML oncogenic domains. Mol. Cell. Biol. 2003, 23, 6174–6186. [Google Scholar] [CrossRef]
- Sharma, P.; Murillas, R.; Zhang, H.; Kuehn, M.R. N4BP1 is a newly identified nucleolar protein that undergoes SUMO-regulated polyubiquitylation and proteasomal turnover at promyelocytic leukemia nuclear bodies. J. Cell Sci. 2010, 123, 1227–1234. [Google Scholar] [CrossRef]
- Oughtred, R.; Rust, J.; Chang, C.; Breitkreutz, B.J.; Stark, C.; Willems, A.; Boucher, L.; Leung, G.; Kolas, N.; Zhang, F.; et al. The BioGRID database: A comprehensive biomedical resource of curated protein, genetic, and chemical interactions. Protein Sci. 2021, 30, 187–200. [Google Scholar] [CrossRef]
- Xu, P.; Roizman, B. The SP100 component of ND10 enhances accumulation of PML and suppresses replication and the assembly of HSV replication compartments. Proc. Natl. Acad. Sci. USA 2017, 114, E3823–E3829. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Wang, P.; Benhenda, S.; Wu, H.; Lallemand-Breitenbach, V.; Zhen, T.; Jollivet, F.; Peres, L.; Li, Y.; Chen, S.J.; Chen, Z.; et al. RING tetramerization is required for nuclear body biogenesis and PML sumoylation. Nat. Commun. 2018, 9, 1277. [Google Scholar] [CrossRef]
- Lang, M.; Jegou, T.; Chung, I.; Richter, K.; Munch, S.; Udvarhelyi, A.; Cremer, C.; Hemmerich, P.; Engelhardt, J.; Hell, S.W.; et al. Three-dimensional organization of promyelocytic leukemia nuclear bodies. J. Cell Sci. 2010, 123, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Liebl, M.C.; Hofmann, T.G. Regulating the p53 Tumor Suppressor Network at PML Biomolecular Condensates. Cancers 2022, 14, 4549. [Google Scholar] [CrossRef] [PubMed]
- Willms, A.; Schupp, H.; Poelker, M.; Adawy, A.; Debus, J.F.; Hartwig, T.; Krichel, T.; Fritsch, J.; Singh, S.; Walczak, H.; et al. TRAIL-receptor 2-a novel negative regulator of p53. Cell Death Dis. 2021, 12, 757. [Google Scholar] [CrossRef] [PubMed]
- Kwek, S.S.; Derry, J.; Tyner, A.L.; Shen, Z.; Gudkov, A.V. Functional analysis and intracellular localization of p53 modified by SUMO-1. Oncogene 2001, 20, 2587–2599. [Google Scholar] [CrossRef] [PubMed]
- Sha, Z.; Blyszcz, T.; Gonzalez-Prieto, R.; Vertegaal, A.C.O.; Goldberg, A.L. Inhibiting ubiquitination causes an accumulation of SUMOylated newly synthesized nuclear proteins at PML bodies. J. Biol. Chem. 2019, 294, 15218–15234. [Google Scholar] [CrossRef] [PubMed]
- Erker, Y.; Neyret-Kahn, H.; Seeler, J.S.; Dejean, A.; Atfi, A.; Levy, L. Arkadia, a novel SUMO-targeted ubiquitin ligase involved in PML degradation. Mol. Cell. Biol. 2013, 33, 2163–2177. [Google Scholar] [CrossRef] [PubMed]
- Bregnard, T.; Ahmed, A.; Semenova, I.V.; Weller, S.K.; Bezsonova, I. The B-box1 domain of PML mediates SUMO E2-E3 complex formation through an atypical interaction with UBC9. Biophys. Chem. 2022, 287, 106827. [Google Scholar] [CrossRef] [PubMed]
- Matt, S.; Hofmann, T.G. Crosstalk between p53 modifiers at PML bodies. Mol. Cell. Oncol. 2018, 5, e1074335. [Google Scholar] [CrossRef]
- Ye, M.; Tang, Y.; Tang, S.; Liu, J.; Wu, K.; Yao, S.; Sun, Y.; Zhou, L.; Deng, T.; Chen, Y.; et al. STIP is a critical nuclear scaffolding protein linking USP7 to p53-Mdm2 pathway regulation. Oncotarget 2015, 6, 34718–34731. [Google Scholar] [CrossRef]
- Nagasaka, M.; Miyajima, C.; Aoki, H.; Aoyama, M.; Morishita, D.; Inoue, Y.; Hayashi, H. Insights into Regulators of p53 Acetylation. Cells 2022, 11, 3825. [Google Scholar] [CrossRef]
- Moller, A.; Sirma, H.; Hofmann, T.G.; Staege, H.; Gresko, E.; Ludi, K.S.; Klimczak, E.; Droge, W.; Will, H.; Schmitz, M.L. Sp100 is important for the stimulatory effect of homeodomain-interacting protein kinase-2 on p53-dependent gene expression. Oncogene 2003, 22, 8731–8737. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mohammed, A.S.; Uversky, V.N. Intrinsic Disorder as a Natural Preservative: High Levels of Intrinsic Disorder in Proteins Found in the 2600-Year-Old Human Brain. Biology 2022, 11, 1704. [Google Scholar] [CrossRef] [PubMed]
- Eskiw, C.H.; Dellaire, G.; Bazett-Jones, D.P. Chromatin contributes to structural integrity of promyelocytic leukemia bodies through a SUMO-1-independent mechanism. J. Biol. Chem. 2004, 279, 9577–9585. [Google Scholar] [CrossRef] [PubMed]
- Torok, D.; Ching, R.W.; Bazett-Jones, D.P. PML nuclear bodies as sites of epigenetic regulation. Front. Biosci. 2009, 14, 1325–1336. [Google Scholar] [CrossRef]
- Delbarre, E.; Ivanauskiene, K.; Spirkoski, J.; Shah, A.; Vekterud, K.; Moskaug, J.O.; Boe, S.O.; Wong, L.H.; Kuntziger, T.; Collas, P. PML protein organizes heterochromatin domains where it regulates histone H3.3 deposition by ATRX/DAXX. Genome Res. 2017, 27, 913–921. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name (Gene Name) | UniProt ID | PER (VSL2B) Disorder | LLPS | DPR Number, Length aa | AHS Number, Length aa | Role in LLPS |
|---|---|---|---|---|---|---|
| Cellular tumor antigen p53 (P53) | P04637 | IDP | Yes | 4.2 | 7.7 | Driver |
| Nuclear autoantigen Sp-100 (SP100) | P23497 | IDP | Yes | 7.5 | 16.1 | Driver |
| Homeodomain-interacting protein kinase 2 (HIPK2) | Q9H2X6 | IDP | Yes | 6.3 | 8.9 | Driver |
| CREB-binding protein (CREBB) | Q92793 | IDP | Yes | 12.2 | 30.3 | Driver |
| Death domain-associated protein 6 (DAXX) | Q9UER7 | IDP | Yes | 6.5 | 14.1 | Driver |
| Small ubiquitin-related modifier 1 (SUMO1) | P63165 | IDP | No | 1.2 | 1.6 | Client |
| SUMO-conjugating enzyme UBC9 (UBE2I) | P63279 | Partial IDP | No | 0 | 0 | No LLPS |
| PML Isoform | UniProt ID | PER (VSL2B) Disorder | LLPS | DPR Number, Length aa | AHS Number, Length aa | Role in LLPS |
|---|---|---|---|---|---|---|
| PML-I | P29590-1 | IDP | Yes | 5.3 | 12.9 | Driver |
| PML-II | P29590-8 | IDP | Yes | 5.4 | 12.1 | Driver |
| PML-III | P29590-9 | IDP | Yes | 5.3 | 8.7 | Driver |
| PML-IV | P29590-5 | IDP | Yes | 4.2 | 9.8 | Driver |
| PML-V | P29590-2 | IDP | Yes | 4.2 | 7.8 | Driver |
| PML-VI | P29590-4 | IDP | Yes | 4.2 | 7.5 | Driver |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silonov, S.A.; Mokin, Y.I.; Nedelyaev, E.M.; Smirnov, E.Y.; Kuznetsova, I.M.; Turoverov, K.K.; Uversky, V.N.; Fonin, A.V. On the Prevalence and Roles of Proteins Undergoing Liquid–Liquid Phase Separation in the Biogenesis of PML-Bodies. Biomolecules 2023, 13, 1805. https://doi.org/10.3390/biom13121805
Silonov SA, Mokin YI, Nedelyaev EM, Smirnov EY, Kuznetsova IM, Turoverov KK, Uversky VN, Fonin AV. On the Prevalence and Roles of Proteins Undergoing Liquid–Liquid Phase Separation in the Biogenesis of PML-Bodies. Biomolecules. 2023; 13(12):1805. https://doi.org/10.3390/biom13121805
Chicago/Turabian StyleSilonov, Sergey A., Yakov I. Mokin, Eugene M. Nedelyaev, Eugene Y. Smirnov, Irina M. Kuznetsova, Konstantin K. Turoverov, Vladimir N. Uversky, and Alexander V. Fonin. 2023. "On the Prevalence and Roles of Proteins Undergoing Liquid–Liquid Phase Separation in the Biogenesis of PML-Bodies" Biomolecules 13, no. 12: 1805. https://doi.org/10.3390/biom13121805
APA StyleSilonov, S. A., Mokin, Y. I., Nedelyaev, E. M., Smirnov, E. Y., Kuznetsova, I. M., Turoverov, K. K., Uversky, V. N., & Fonin, A. V. (2023). On the Prevalence and Roles of Proteins Undergoing Liquid–Liquid Phase Separation in the Biogenesis of PML-Bodies. Biomolecules, 13(12), 1805. https://doi.org/10.3390/biom13121805