
Expression and Function of ABC Transporters in Human Alveolar Epithelial Cells

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
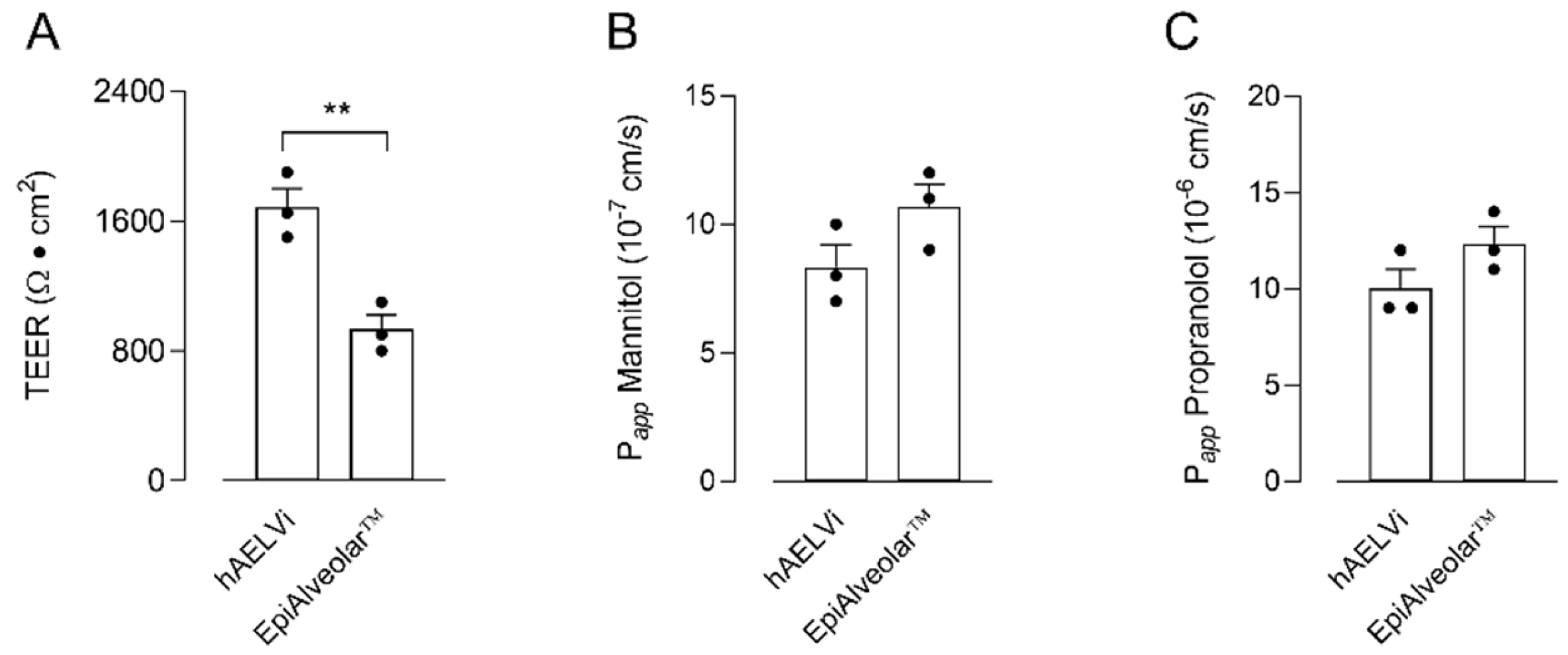
2.2. Transepithelial Electrical Resistance (TEER) and Cellular Permeability
2.3. Bidirectional Transport Studies
2.4. Calculation of Papp
2.5. Absolute Quantification of mRNA Expression
2.6. Western Blot Analysis
2.7. Immunocytochemistry
2.8. Statistical Analysis
2.9. Materials
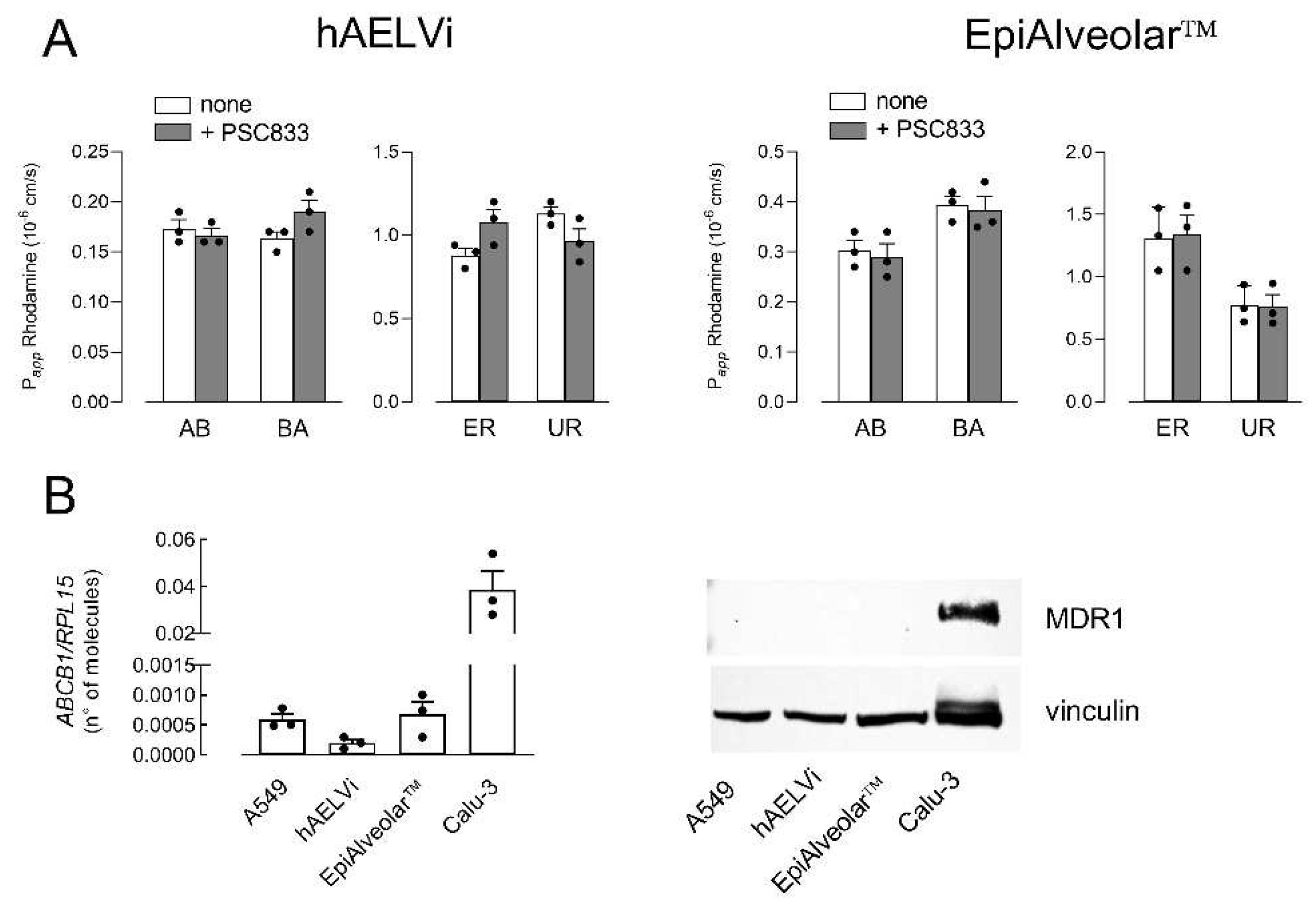
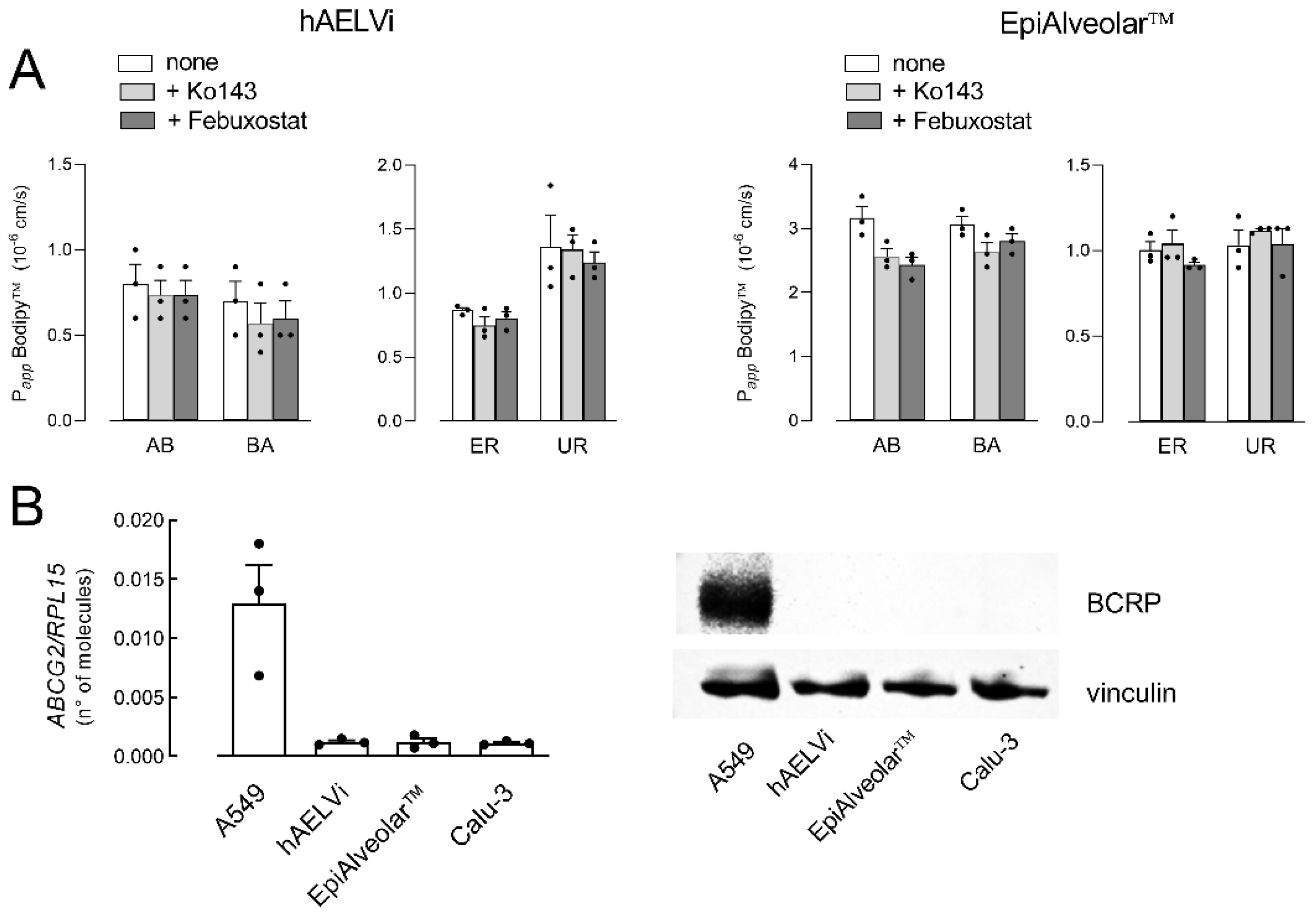
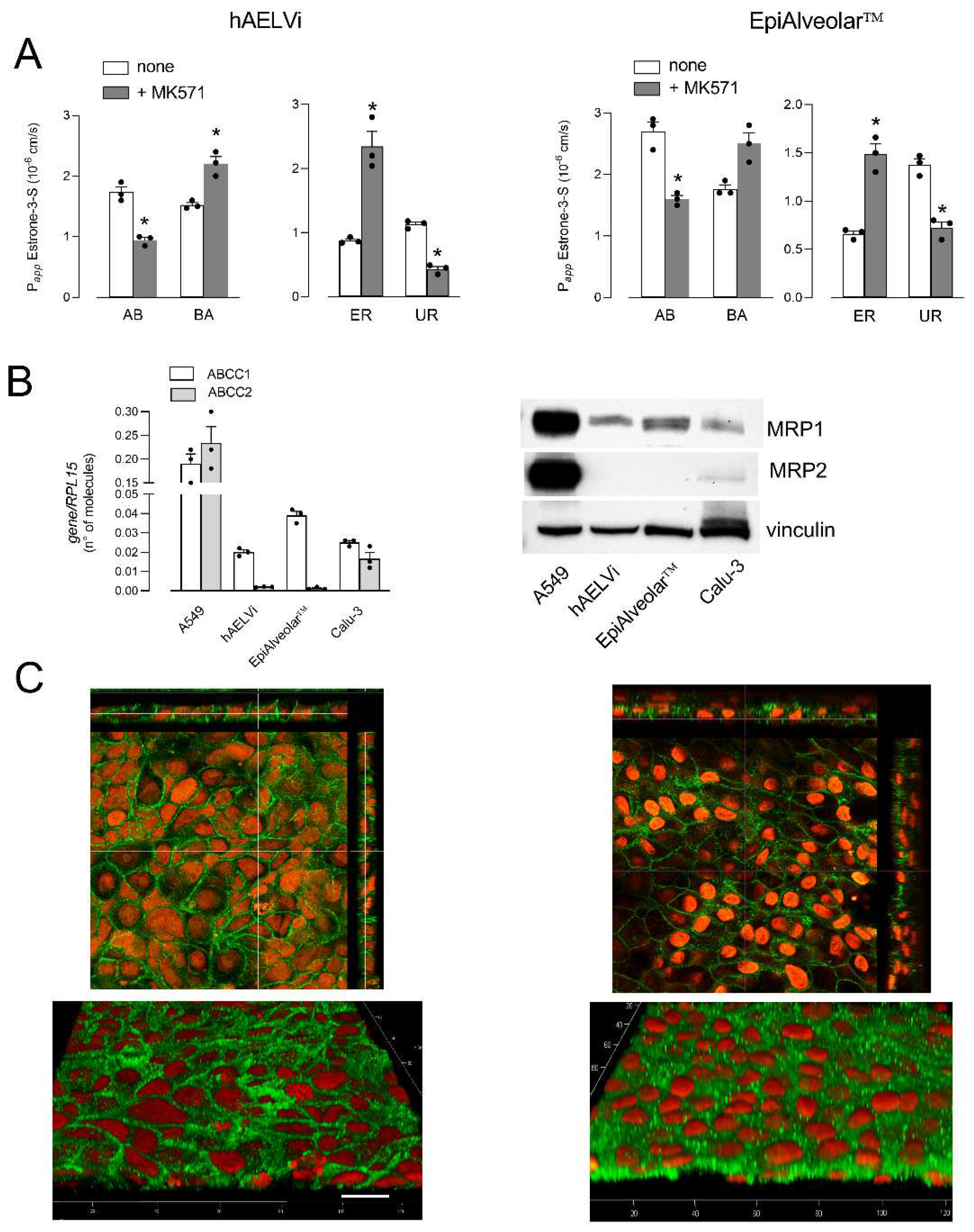
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human atp-binding cassette (abc) transporter superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef]
- Nickel, S.; Clerkin, C.G.; Selo, M.A.; Ehrhardt, C. Transport mechanisms at the pulmonary mucosa: Implications for drug delivery. Expert Opin. Drug Deliv. 2016, 13, 667–690. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Johnson, E.; Lewinson, O. Abc transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of abc transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Liu, X. Abc family transporters. Adv. Exp. Med. Biol. 2019, 1141, 13–100. [Google Scholar]
- Redente, E.F. Abc transporters: An overlooked mechanism of drug failure in our preclinical models? Am. J. Respir. Cell Mol. Biol. 2020, 62, 130–131. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, M.R.; Haffez, H. The potential roles of retinoids in combating drug resistance in cancer: Implications of atp-binding cassette (abc) transporters. Open Biol. 2022, 12, 220001. [Google Scholar] [CrossRef]
- Surowiak, P.; Pawelczyk, K.; Maciejczyk, A.; Pudelko, M.; Kolodziej, J.; Zabel, M. Positive correlation between cyclooxygenase 2 and the expression of abc transporters in non-small cell lung cancer. Anticancer Res. 2008, 28, 2967–2974. [Google Scholar] [PubMed]
- Xiao, H.; Zheng, Y.; Ma, L.; Tian, L.; Sun, Q. Clinically-relevant abc transporter for anti-cancer drug resistance. Front. Pharmacol. 2021, 12, 648407. [Google Scholar] [CrossRef]
- van der Deen, M.; de Vries, E.G.; Timens, W.; Scheper, R.J.; Timmer-Bosscha, H.; Postma, D.S. Atp-binding cassette (abc) transporters in normal and pathological lung. Respir. Res. 2005, 6, 59. [Google Scholar] [CrossRef]
- Bosquillon, C. Drug transporters in the lung--do they play a role in the biopharmaceutics of inhaled drugs? J. Pharm. Sci. 2010, 99, 2240–2255. [Google Scholar] [CrossRef]
- Han, J.Y.; Lim, H.S.; Yoo, Y.K.; Shin, E.S.; Park, Y.H.; Lee, S.Y. Associations of abcb1, abcc2, and abcg2 polymorphisms with irinotecan-pharmacokinetics and clinical outcome in patients with advanced non-small cell lung cancer. Cancer 2007, 110, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Vesel, M.; Rapp, J.; Feller, D.; Kiss, E.; Jaromi, L.; Meggyes, M. Abcb1 and abcg2 drug transporters are differentially expressed in non-small cell lung cancers (nsclc) and expression is modified by cisplatin treatment via altered wnt signaling. Respir. Res. 2017, 18, 52. [Google Scholar] [CrossRef]
- McElroy, M.C.; Kasper, M. The use of alveolar epithelial type i cell-selective markers to investigate lung injury and repair. Eur. Respir. J. 2004, 24, 664–673. [Google Scholar] [CrossRef]
- Aspal, M.; Zemans, R.L. Mechanisms of atii-to-ati cell differentiation during lung regeneration. Int. J. Mol. Sci. 2020, 21, 3188. [Google Scholar] [CrossRef]
- Foster, K.A.; Oster, C.G.; Mayer, M.M.; Avery, M.L.; Audus, K.L. Characterization of the a549 cell line as a type ii pulmonary epithelial cell model for drug metabolism. Exp. Cell Res. 1998, 243, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Hermanns, M.I.; Unger, R.E.; Kehe, K.; Peters, K.; Kirkpatrick, C.J. Lung epithelial cell lines in coculture with human pulmonary microvascular endothelial cells: Development of an alveolo-capillary barrier in vitro. Lab. Invest. 2004, 84, 736–752. [Google Scholar] [CrossRef]
- Ren, H.; Birch, N.P.; Suresh, V. An optimised human cell culture model for alveolar epithelial transport. PLoS ONE 2016, 11, e0165225. [Google Scholar] [CrossRef] [PubMed]
- Rucka, Z.; Vanhara, P.; Koutna, I.; Tesarova, L.; Potesilova, M.; Stejskal, S. Differential effects of insulin and dexamethasone on pulmonary surfactant-associated genes and proteins in a549 and h441 cells and lung tissue. Int. J. Mol. Med. 2013, 32, 211–218. [Google Scholar] [CrossRef]
- Salomon, J.J.; Muchitsch, V.E.; Gausterer, J.C.; Schwagerus, E.; Huwer, H.; Daum, N. The cell line ncl-h441 is a useful in vitro model for transport studies of human distal lung epithelial barrier. Mol. Pharm. 2014, 11, 995–1006. [Google Scholar] [CrossRef]
- Kuehn, A.; Kletting, S.; de Souza Carvalho-Wodarz, C.; Repnik, U.; Griffiths, G.; Fischer, U. Human alveolar epithelial cells expressing tight junctions to model the air-blood barrier. ALTEX 2016, 33, 251–260. [Google Scholar] [CrossRef]
- Delie, F.; Rubas, W. A human colonic cell line sharing similarities with enterocytes as a model to examine oral absorption: Advantages and limitations of the caco-2 model. Crit. Rev. Ther. Drug Carrier Syst. 1997, 14, 221–286. [Google Scholar] [CrossRef]
- Hidalgo, I.J. Assessing the absorption of new pharmaceuticals. Curr Top. Med. Chem. 2001, 1, 385–401. [Google Scholar] [CrossRef]
- Pires, C.L.; Praca, C.; Martins, P.A.T.; Batista de Carvalho, A.L.M.; Ferreira, L.; Marques, M.P.M. Re-use of caco-2 monolayers in permeability assays-validation regarding cell monolayer integrity. Pharmaceutics 2021, 13, 1563. [Google Scholar] [CrossRef] [PubMed]
- Adson, A.; Raub, T.J.; Burton, P.S.; Barsuhn, C.L.; Hilgers, A.R.; Audus, K.L. Quantitative approaches to delineate paracellular diffusion in cultured epithelial cell monolayers. J. Pharm. Sci. 1994, 83, 1529–1536. [Google Scholar] [CrossRef]
- Keemink, J.; Bergstrom, C.A.S. Caco-2 cell conditions enabling studies of drug absorption from digestible lipid-based formulations. Pharm. Res. 2018, 35, 74. [Google Scholar] [CrossRef]
- Barilli, A.; Rotoli, B.M.; Visigalli, R.; Ingoglia, F.; Cirlini, M.; Prandi, B. Gliadin-mediated production of polyamines by raw264.7 macrophages modulates intestinal epithelial permeability in vitro. Biochim. Biophys. Acta 2015, 1852, 1779–1786. [Google Scholar] [CrossRef]
- Rotoli, B.M.; Barilli, A.; Visigalli, R.; Ferrari, F.; Dall’Asta, V. Y+lat1 and y+lat2 contribution to arginine uptake in different human cell models: Implications in the pathophysiology of lysinuric protein intolerance. J. Cell Mol. Med. 2020, 24, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Barilli, A.; Visigalli, R.; Ferrari, F.; Di Lascia, M.; Riccardi, B.; Puccini, P. Organic cation transporters (octs/octns) in human primary alveolar epithelial cells. Biochem. Biophys. Res. Commun. 2021, 576, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Archinal-Mattheis, A.; Rzepka, R.W.; Watanabe, T.; Kokubu, N.; Itoh, Y.; Combates, N.J. Analysis of the interactions of sdz psc 833 ([3’-keto-bmt1]-val2]-cyclosporine), a multidrug resistance modulator, with p-glycoprotein. Oncol. Res. 1995, 7, 603–610. [Google Scholar] [PubMed]
- Marsousi, N.; Doffey-Lazeyras, F.; Rudaz, S.; Desmeules, J.A.; Daali, Y. Intestinal permeability and p-glycoprotein-mediated efflux transport of ticagrelor in caco-2 monolayer cells. Fundam. Clin. Pharmacol. 2016, 30, 577–584. [Google Scholar] [CrossRef]
- Rotoli, B.M.; Barilli, A.; Visigalli, R.; Ferrari, F.; Frati, C.; Lagrasta, C.A. Characterization of abc transporters in epiairway, a cellular model of normal human bronchial epithelium. Int. J. Mol. Sci. 2020, 21, 3190. [Google Scholar] [CrossRef]
- Miyata, H.; Takada, T.; Toyoda, Y.; Matsuo, H.; Ichida, K.; Suzuki, H. Identification of febuxostat as a new strong abcg2 inhibitor: Potential applications and risks in clinical situations. Front. Pharmacol. 2016, 7, 518. [Google Scholar] [CrossRef]
- Allen, J.D.; van Loevezijn, A.; Lakhai, J.M.; van der Valk, M.; van Tellingen, O.; Reid, G. Potent and specific inhibition of the breast cancer resistance protein multidrug transporter in vitro and in mouse intestine by a novel analogue of fumitremorgin c. Mol. Cancer Ther. 2002, 1, 417–425. [Google Scholar] [PubMed]
- Gekeler, V.; Ise, W.; Sanders, K.H.; Ulrich, W.R.; Beck, J. The leukotriene ltd4 receptor antagonist mk571 specifically modulates mrp associated multidrug resistance. Biochem. Biophys. Res. Commun. 1995, 208, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.O.; Topp, E.; Makagiansar, I.; Siahaan, T.; Yazdanian, M.; Audus, K.L. Multidrug resistance-associated protein-1 functional activity in calu-3 cells. J. Pharmacol. Exp. Ther. 2001, 298, 1199–1205. [Google Scholar]
- Salomon, J.J.; Ehrhardt, C. Nanoparticles attenuate p-glycoprotein/mdr1 function in a549 human alveolar epithelial cells. Eur. J. Pharm. Biopharm. 2011, 77, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Togami, K.; Yamaguchi, K.; Tada, H.; Chono, S. Assessment of transporter-mediated efflux of nintedanib using in vitro cell line models of idiopathic pulmonary fibrosis. Pharmazie 2020, 75, 371–374. [Google Scholar] [PubMed]
- Endter, S.; Becker, U.; Daum, N.; Huwer, H.; Lehr, C.M.; Gumbleton, M. P-glycoprotein (mdr1) functional activity in human alveolar epithelial cell monolayers. Cell Tissue Res. 2007, 328, 77–84. [Google Scholar] [CrossRef]
- Park, J.K.; Coffey, N.J.; Bodine, S.P.; Zawatsky, C.N.; Jay, L.; Gahl, W.A. Bleomycin induces drug efflux in lungs. A pitfall for pharmacological studies of pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2020, 62, 178–190. [Google Scholar] [CrossRef]
- Nickel, S.; Selo, M.A.; Fallack, J.; Clerkin, C.G.; Huwer, H.; Schneider-Daum, N. Expression and activity of breast cancer resistance protein (bcrp/abcg2) in human distal lung epithelial cells in vitro. Pharm. Res. 2017, 34, 2477–2487. [Google Scholar] [CrossRef] [PubMed]
- Selo, M.A.; Delmas, A.S.; Springer, L.; Zoufal, V.; Sake, J.A.; Clerkin, C.G. Tobacco smoke and inhaled drugs alter expression and activity of multidrug resistance-associated protein-1 (mrp1) in human distal lung epithelial cells in vitro. Front. Bioeng. Biotechnol. 2020, 8, 1030. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visigalli, R.; Rotoli, B.M.; Ferrari, F.; Di Lascia, M.; Riccardi, B.; Puccini, P.; Dall’Asta, V.; Barilli, A. Expression and Function of ABC Transporters in Human Alveolar Epithelial Cells. Biomolecules 2022, 12, 1260. https://doi.org/10.3390/biom12091260
Visigalli R, Rotoli BM, Ferrari F, Di Lascia M, Riccardi B, Puccini P, Dall’Asta V, Barilli A. Expression and Function of ABC Transporters in Human Alveolar Epithelial Cells. Biomolecules. 2022; 12(9):1260. https://doi.org/10.3390/biom12091260
Chicago/Turabian StyleVisigalli, Rossana, Bianca Maria Rotoli, Francesca Ferrari, Maria Di Lascia, Benedetta Riccardi, Paola Puccini, Valeria Dall’Asta, and Amelia Barilli. 2022. "Expression and Function of ABC Transporters in Human Alveolar Epithelial Cells" Biomolecules 12, no. 9: 1260. https://doi.org/10.3390/biom12091260
APA StyleVisigalli, R., Rotoli, B. M., Ferrari, F., Di Lascia, M., Riccardi, B., Puccini, P., Dall’Asta, V., & Barilli, A. (2022). Expression and Function of ABC Transporters in Human Alveolar Epithelial Cells. Biomolecules, 12(9), 1260. https://doi.org/10.3390/biom12091260