
The Crystal Structure of the Defense Conferring Rice Protein OsJAC1 Reveals a Carbohydrate Binding Site on the Dirigent-like Domain

 , , and
, , and
Abstract
1. Introduction
2. Methods and Materials
2.1. Materials
2.2. Crystallization, Soaking Experiment and Data Collection
2.3. Structure Determination and Refinement
2.4. Ligand-Protein and Protein-Protein Docking
2.5. Hemagglutination Assay
2.6. Growth Inhibition Assay
3. Results
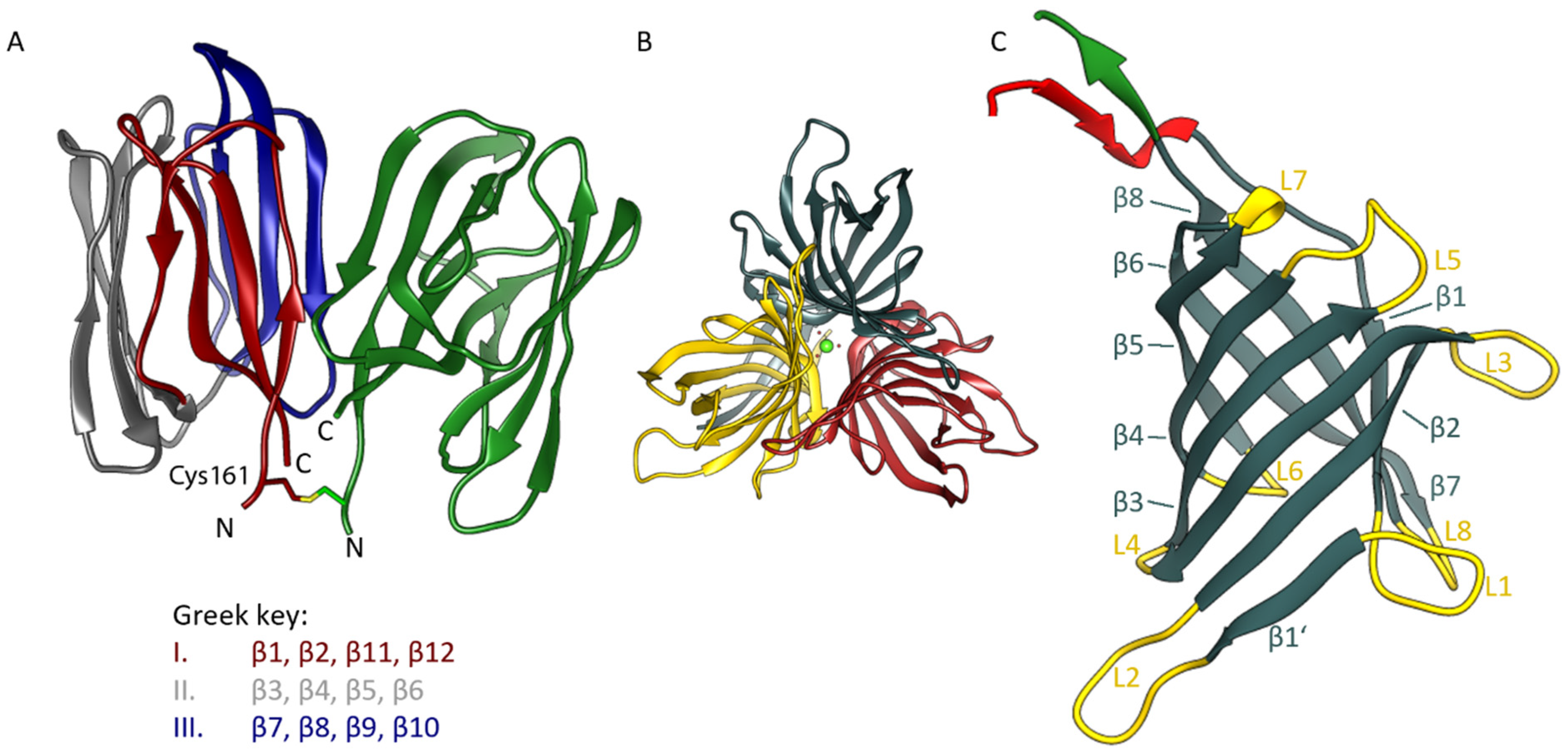
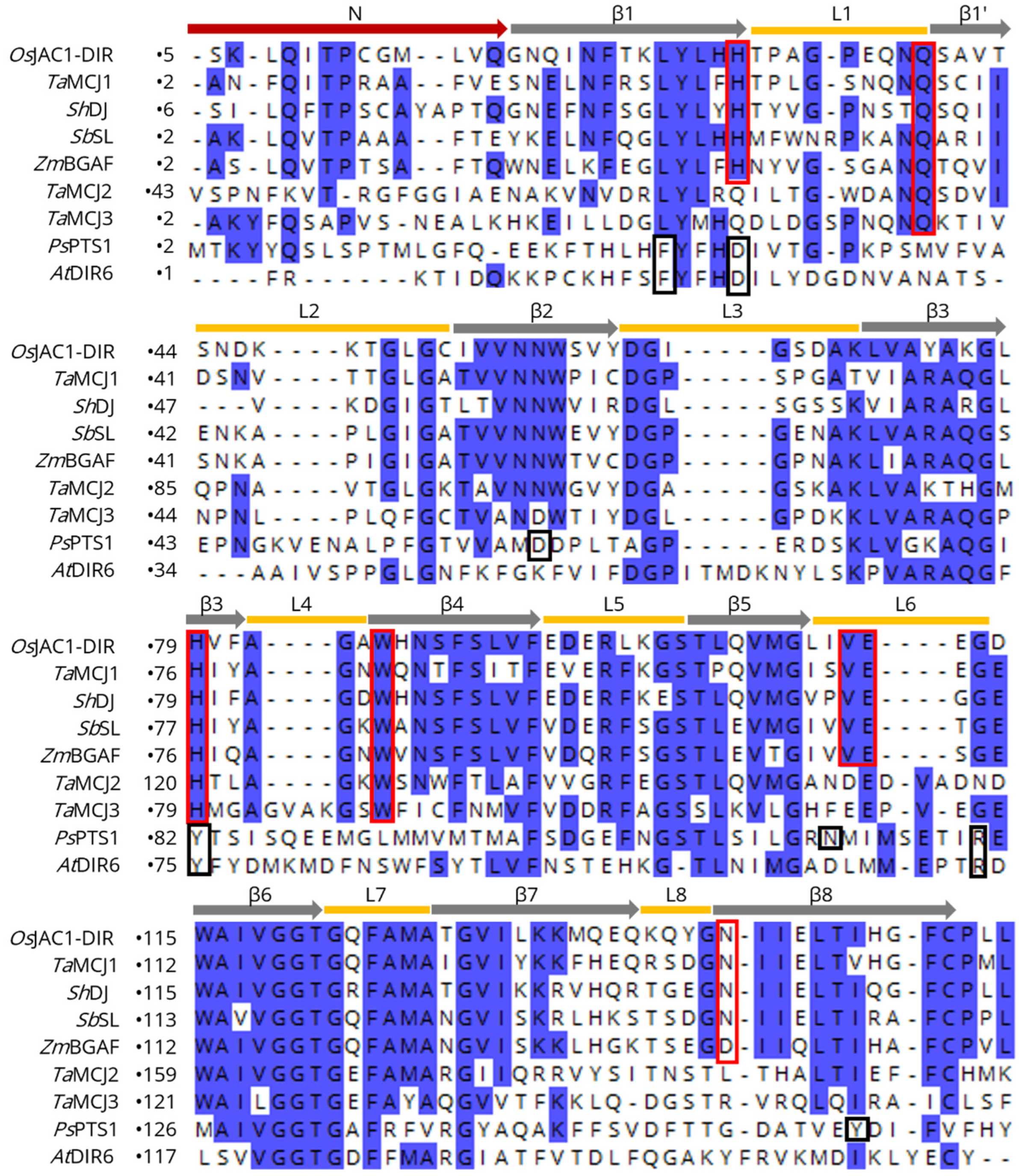
3.1. Overall Structure of the OsJAC1 Domains
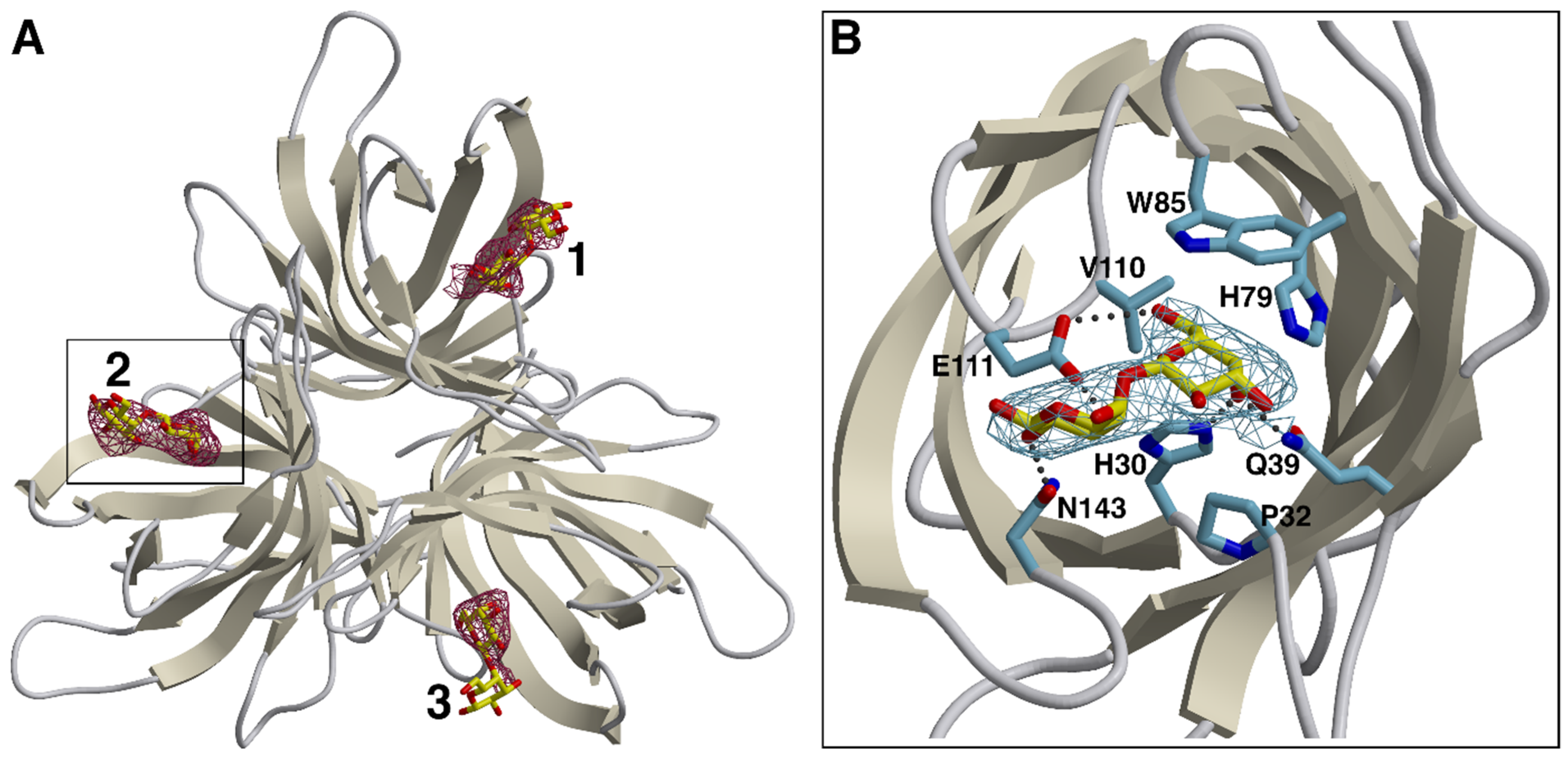
3.2. Carbohydrate Binding Site of the OsJAC1-DIR Domain
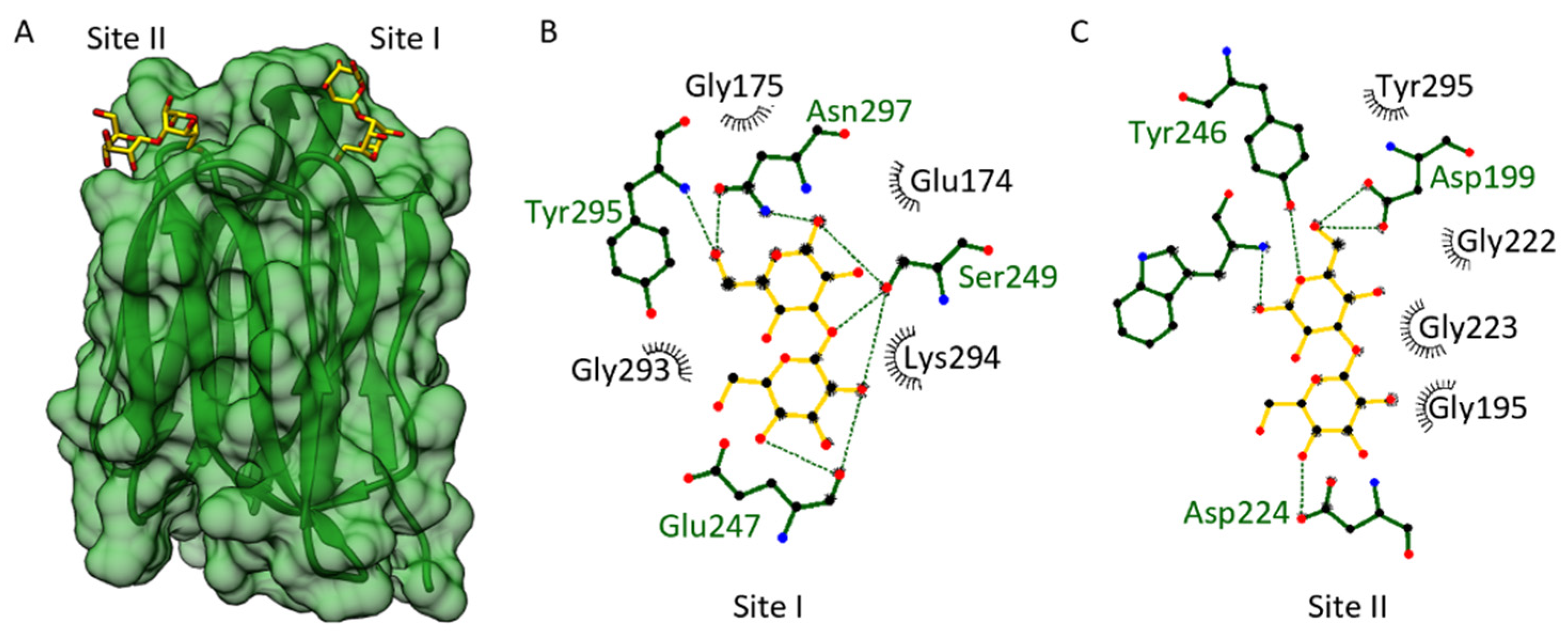
3.3. Carbohydrate Binding Site of the OsJAC1-JRL Domain
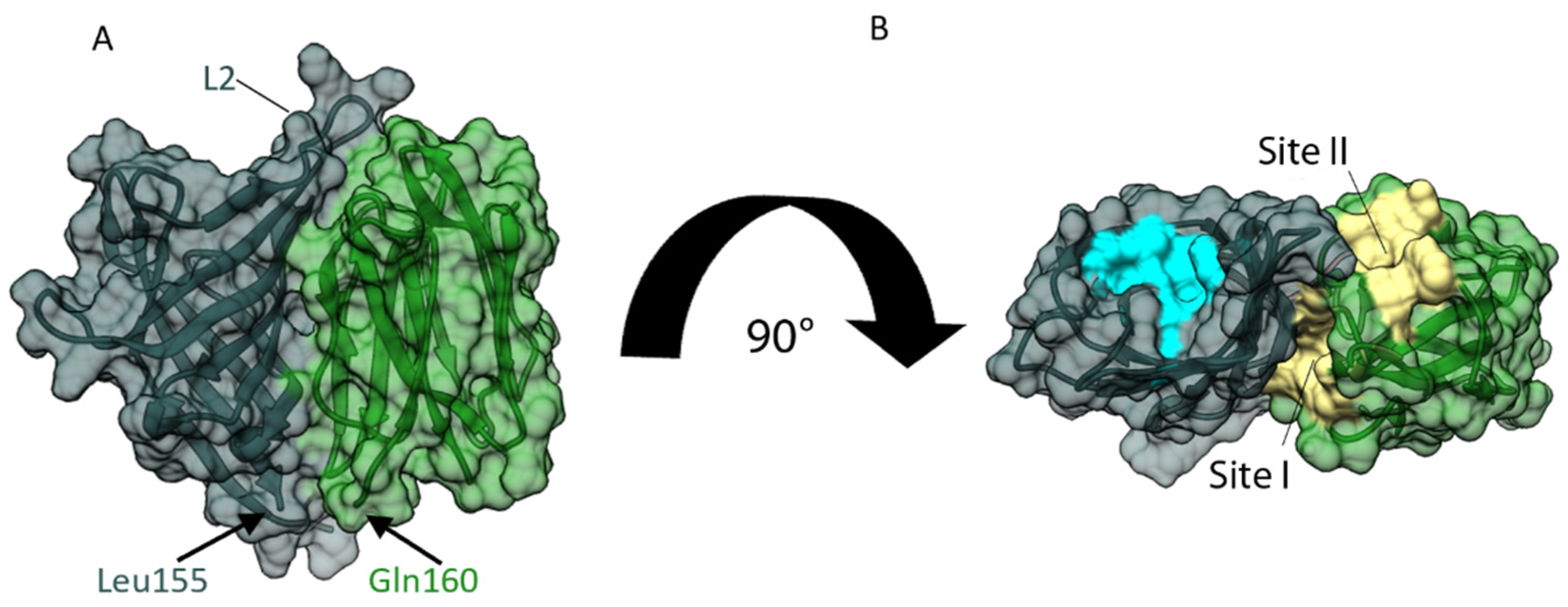
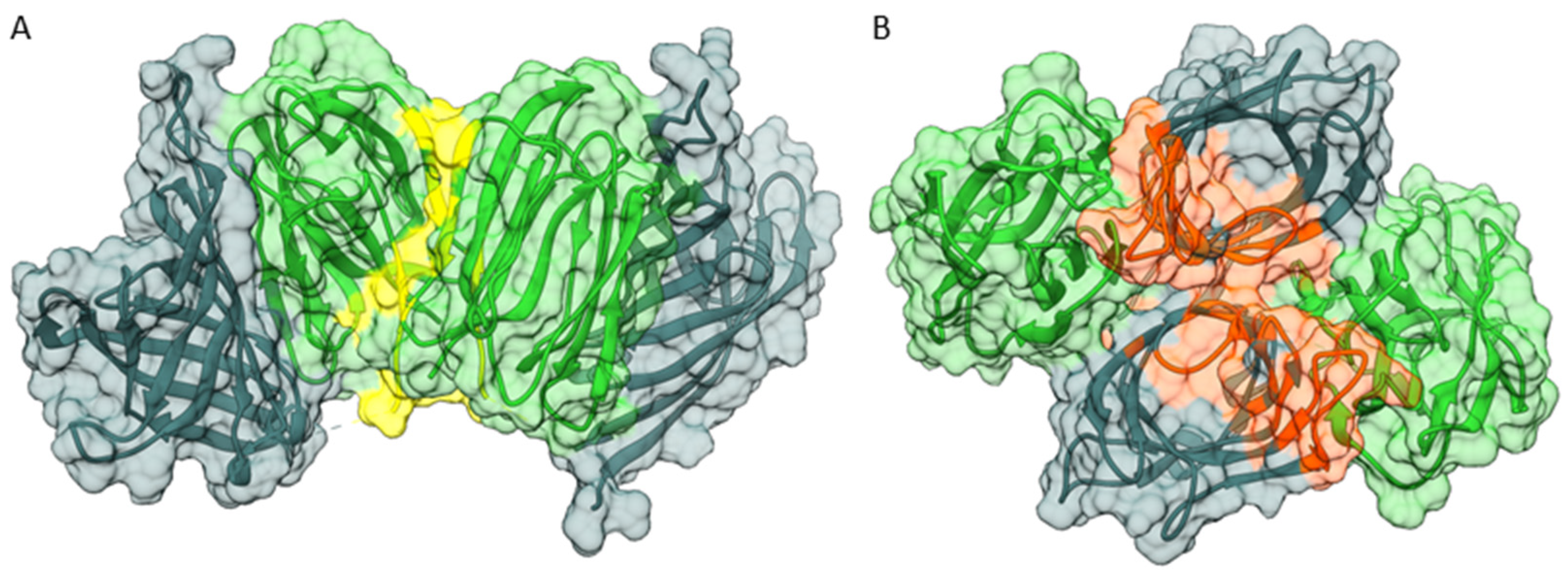
3.4. Domain-Domain Interaction Analysis
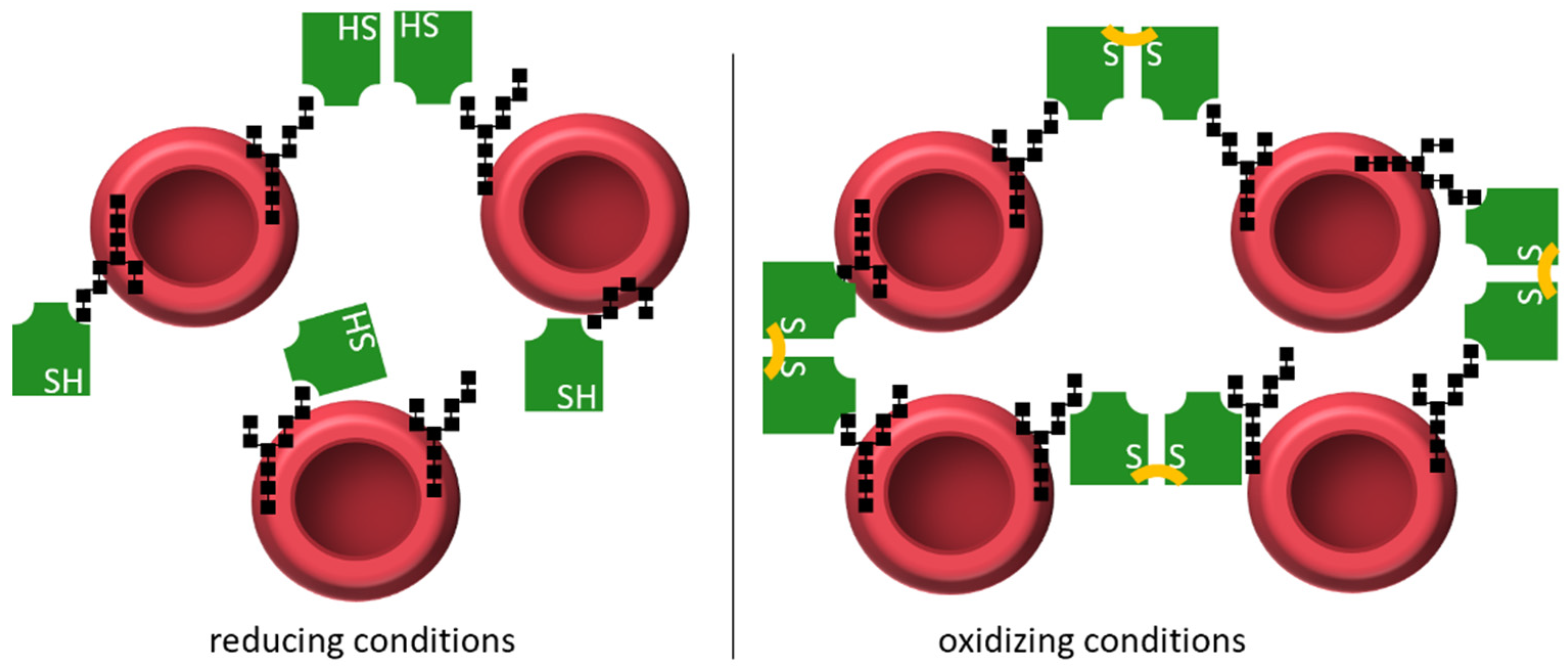
3.5. Hemagglutination Assay
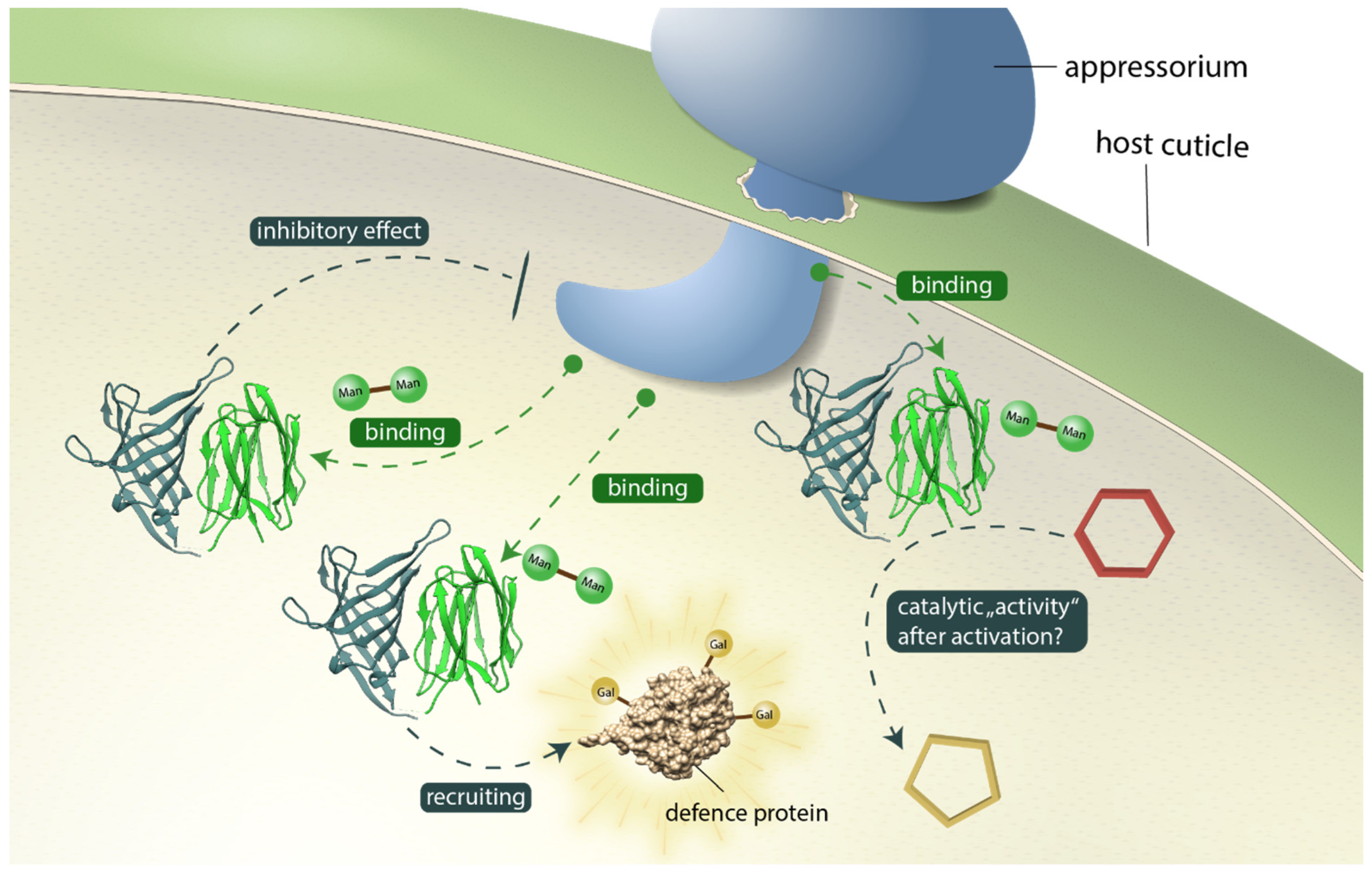
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oerke, E.-C. Crop losses to pests. J. Agricult. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Ma, Q.-H. Monocot chimeric jacalins: A novel subfamily of plant lectins. Crit. Rev. Biotechnol. 2014, 34, 300–306. [Google Scholar] [CrossRef]
- Esch, L.; Schaffrath, U. An update on jacalin-like lectins and their role in plant defense. Int. J. Mol. Sci. 2017, 18, 1592. [Google Scholar] [CrossRef] [PubMed]
- Li, H.M.; Rotter, D.; Bonos, S.A.; Meyer, W.A.; Belanger, F.C. Identification of a gene in the process of being lost from the genus Agrostis. Plant. Physiol. 2005, 138, 2386–2395. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Huang, B.; Chen, J.; Huang, Z.; Yu, P.; Ruan, S.; Zhang, Z. Genome-wide identification and expression analysis of dirigent-jacalin genes from plant chimeric lectins in Moso bamboo (Phyllostachys edulis). PLoS ONE 2021, 16, e0248318. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Li, J.; Xu, Y.; Xu, Z.; Chong, K. Phosphorylation modification of wheat lectin VER2 is associated with vernalization-induced O-GlcNAc signaling and intracellular motility. PLoS ONE 2009, 4, e4854. [Google Scholar] [CrossRef]
- Subramanyam, S.; Smith, D.F.; Clemens, J.C.; Webb, M.A.; Sardesai, N.; Williams, C.E. Functional characterization of HFR1, a high-mannose N-glycan-specific wheat lectin induced by Hessian fly larvae. Plant. Physiol. 2008, 147, 1412–1426. [Google Scholar] [CrossRef]
- Kittur, F.S.; Yu, H.Y.; Bevan, D.R.; Esen, A. Homolog of the maize β-glucosidase aggregating factor from sorghum is a jacalin-related GalNAc-specific lectin but lacks protein aggregating activity. Glycobiology 2009, 19, 277–287. [Google Scholar] [CrossRef][Green Version]
- Kittur, F.S.; Lalgondar, M.; Yu, H.Y.; Bevan, D.R.; Esen, A. Maize β-glucosidase-aggregating factor is a polyspecific jacalin-related chimeric lectin, and its lectin domain is responsible for β-glucosidase aggregation. J. Biol. Chem. 2007, 282, 7299–7311. [Google Scholar] [CrossRef]
- Andrade, L.M.; Peixoto-Junior, R.F.; Ribeiro, R.V.; Nóbile, P.M.; Brito, M.S.; Marchiori, P.E.R.; Carlin, S.D.; Martins, A.P.B.; Goldman, M.H.S.; Llerena, J.P.P. Biomass accumulation and cell wall structure of rice plants overexpressing a dirigent-jacalin of sugarcane (ShDJ) under varying conditions of water availability. Front. Plant. Sci. 2019, 10, 65. [Google Scholar] [CrossRef]
- Jiang, J.-F.; Han, Y.; Xing, L.-J.; Xu, Y.-Y.; Xu, Z.-H.; Chong, K. Cloning and expression of a novel cDNA encoding a mannose-specific jacalin-related lectin from Oryza sativa. Toxicon 2006, 47, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.-H.; Han, J.-Q. Identification of monocot chimeric jacalin family reveals functional diversity in wheat. Planta 2021, 253, 30. [Google Scholar] [CrossRef] [PubMed]
- Weidenbach, D.; Esch, L.; Möller, C.; Hensel, G.; Kumlehn, J.; Höfle, C.; Hückelhoven, R.; Schaffrath, U. Polarized defense against fungal pathogens is mediated by the Jacalin-related lectin domain of modular Poaceae-specific proteins. Mol. Plant 2016, 9, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.-H.; Tian, B.; Li, Y.-L. Overexpression of a wheat jasmonate-regulated lectin increases pathogen resistance. Biochimie 2010, 92, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.-H.; Zhen, W.-B.; Liu, Y.-C. Jacalin domain in wheat jasmonate-regulated protein Ta-JA1 confers agglutinating activity and pathogen resistance. Biochimie 2013, 95, 359–365. [Google Scholar] [CrossRef]
- Kittur, F.S.; Yu, H.Y.; Bevan, D.R.; Esen, A. Deletion of the N-terminal dirigent domain in maize β-glucosidase aggregating factor and its homolog sorghum lectin dramatically alters the sugar-specificities of their lectin domains. Plant Physiol. Biochem. 2010, 48, 731–734. [Google Scholar] [CrossRef]
- Davin, L.B.; Wang, H.-B.; Crowell, A.L.; Bedgar, D.L.; Martin, D.M.; Sarkanen, S.; Lewis, N.G. Stereoselective bimolecular phenoxy radical coupling by an auxiliary (dirigent) protein without an active center. Science 1997, 275, 362–367. [Google Scholar] [CrossRef]
- Halls, S.C.; Davin, L.B.; Kramer, D.M.; Lewis, N.G. Kinetic study of coniferyl alcohol radical binding to the (+)-pinoresinol forming dirigent protein. Biochemistry 2004, 43, 2587–2595. [Google Scholar] [CrossRef]
- Effenberger, I.; Zhang, B.; Li, L.; Wang, Q.; Liu, Y.; Klaiber, I.; Pfannstiel, J.; Wang, Q.; Schaller, A. Dirigent proteins from cotton (Gossypium sp.) for the atropselective synthesis of gossypol. Angew. Chem. Int. Ed. 2015, 54, 14660–14663. [Google Scholar] [CrossRef]
- Meng, Q.; Moinuddin, S.G.; Kim, S.-J.; Bedgar, D.L.; Costa, M.A.; Thomas, D.G.; Young, R.P.; Smith, C.A.; Cort, J.R.; Davin, L.B. Pterocarpan synthase (PTS) structures suggest a common quinone methide–stabilizing function in dirigent proteins and proteins with dirigent-like domains. J. Biol. Chem. 2020, 295, 11584–11601. [Google Scholar] [CrossRef]
- Uchida, K.; Akashi, T.; Aoki, T. The Missing Link in Leguminous Pterocarpan Biosynthesis is a Dirigent Domain-Containing Protein with Isoflavanol Dehydratase Activity. Plant Cell Physiol. 2017, 58, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Li, R.-J.; Sun, J.-T.; Ma, F.; Zhang, H.-X.; Jin, J.-H.; Ali, M.; ul Haq, S.; Wang, J.-E.; Gong, Z.-H. Genome-wide analysis of dirigent gene family in pepper (Capsicum annuum L.) and characterization of CaDIR7 in biotic and abiotic stresses. Sci. Rep. 2018, 8, 5500. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-L.; Xu, L.-P.; Fang, J.-P.; Su, Y.-C.; Fu, H.-Y.; Que, Y.-X.; Xu, J.-S. A novel dirigent protein gene with highly stem-specific expression from sugarcane, response to drought, salt and oxidative stresses. Plant Cell Rep. 2012, 31, 1801–1812. [Google Scholar]
- Hosmani, P.S.; Kamiya, T.; Danku, J.; Naseer, S.; Geldner, N.; Guerinot, M.L.; Salt, D.E. Dirigent domain-containing protein is part of the machinery required for formation of the lignin-based Casparian strip in the root. Proc. Nat. Acad. Sci. USA 2013, 110, 14498–14503. [Google Scholar] [CrossRef]
- Huwa, N.; Weiergräber, O.H.; Kirsch, C.; Schaffrath, U.; Classen, T. Biochemical and Initial Structural Characterization of the Monocot Chimeric Jacalin OsJAC1. Int. J. Mol. Sci. 2021, 22, 5639. [Google Scholar] [CrossRef] [PubMed]
- Paniagua, C.; Bilkova, A.; Jackson, P.; Dabravolski, S.; Riber, W.; Didi, V.; Houser, J.; Gigli-Bisceglia, N.; Wimmerova, M.; Budínská, E. Dirigent proteins in plants: Modulating cell wall metabolism during abiotic and biotic stress exposure. J. Exp. Bot. 2017, 68, 3287–3301. [Google Scholar] [CrossRef]
- Kabsch, W. Xds. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef]
- Vagin, A.; Lebedev, A. MoRDa, an automatic molecular replacement pipeline. Acta Crystallogr. Sect. A Found. Adv. 2015, 71, S19. [Google Scholar] [CrossRef]
- Vagin, A.; Teplyakov, A. MOLREP: An automated program for molecular replacement. J. Appl. Cryst. 1997, 30, 1022–1025. [Google Scholar] [CrossRef]
- Moriarty, N.W.; Grosse-Kunstleve, R.W.; Adams, P.D. electronic Ligand Builder and Optimization Workbench (eLBOW): A tool for ligand coordinate and restraint generation. Acta Crystallogr. Sect. D Biol. Crystallogr. 2009, 65, 1074–1080. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Van Zundert, G.; Rodrigues, J.; Trellet, M.; Schmitz, C.; Kastritis, P.; Karaca, E.; Melquiond, A.; van Dijk, M.; De Vries, S.; Bonvin, A. The HADDOCK2. 2 web server: User-friendly integrative modeling of biomolecular complexes. J. Mol. Biol. 2016, 428, 720–725. [Google Scholar] [CrossRef] [PubMed]
- De Vries, S.J.; Bonvin, A.M. CPORT: A consensus interface predictor and its performance in prediction-driven docking with HADDOCK. PLoS ONE 2011, 6, e17695. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand–Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Sano, K.; Ogawa, H. Hemagglutination (inhibition) assay. In Lectins; Springer: Berlin/Heidelberg, Germany, 2014; pp. 47–52. [Google Scholar]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Pratap, J.; Jeyaprakash, A.A.; Rani, P.G.; Sekar, K.; Surolia, A.; Vijayan, M. Crystal structures of artocarpin, a Moraceae lectin with mannose specificity, and its complex with methyl-α-D-mannose: Implications to the generation of carbohydrate specificity. J. Mol. Biol. 2002, 317, 237–247. [Google Scholar] [CrossRef]
- Nakae, S.; Shionyu, M.; Ogawa, T.; Shirai, T. Structures of jacalin-related lectin PPL3 regulating pearl shell biomineralization. Proteins Struct. Funct. Bioinf. 2018, 86, 644–653. [Google Scholar] [CrossRef]
- Gasper, R.; Effenberger, I.; Kolesinski, P.; Terlecka, B.; Hofmann, E.; Schaller, A. Dirigent protein mode of action revealed by the crystal structure of AtDIR6. Plant Physiol. 2016, 172, 2165–2175. [Google Scholar] [CrossRef]
- Fenn, T.D.; Ringe, D.; Petsko, G.A. POVScript+: A program for model and data visualization using persistence of vision ray-tracing. J. Appl. Cryst. 2003, 36, 944–947. [Google Scholar] [CrossRef]
- Merritt, E.A.; Bacon, D.J. [26] Raster3D: Photorealistic molecular graphics. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1997; Volume 277, pp. 505–524. [Google Scholar]
- Kabsch, W.; Sander, C. Dictionary of protein secondary structure: Pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 1983, 22, 2577–2637. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Boratyn, G.M.; Schäffer, A.A.; Agarwala, R.; Altschul, S.F.; Lipman, D.J.; Madden, T.L. Domain enhanced lookup time accelerated BLAST. Biol. Direct 2012, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A completely reimplemented MPI bioinformatics toolkit with a new HHpred server at its core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Peumans, W.J.; Hause, B.; Van Damme, E.J. The galactose-binding and mannose-binding jacalin-related lectins are located in different sub-cellular compartments. FEBS Lett. 2000, 477, 186–192. [Google Scholar] [CrossRef]
- Rougé, P.; Peumans, W.J.; Barre, A.; Van Damme, E.J. A structural basis for the difference in specificity between the two jacalin-related lectins from mulberry (Morus nigra) bark. Biochem. Biophys. Res. Commun. 2003, 304, 91–97. [Google Scholar] [CrossRef]
- Meagher, J.L.; Winter, H.C.; Ezell, P.; Goldstein, I.J.; Stuckey, J.A. Crystal structure of banana lectin reveals a novel second sugar binding site. Glycobiology 2005, 15, 1033–1042. [Google Scholar] [CrossRef]
- Azarkan, M.; Feller, G.; Vandenameele, J.; Herman, R.; El Mahyaoui, R.; Sauvage, E.; Broeck, A.V.; Matagne, A.; Charlier, P.; Kerff, F. Biochemical and structural characterization of a mannose binding jacalin-related lectin with two-sugar binding sites from pineapple (Ananas comosus) stem. Sci. Rep. 2018, 8, 11508. [Google Scholar] [CrossRef]
- Nagae, M.; Mishra, S.K.; Hanashima, S.; Tateno, H.; Yamaguchi, Y. Distinct roles for each N-glycan branch interacting with mannose-binding type Jacalin-related lectins Orysata and Calsepa. Glycobiology 2017, 27, 1120–1133. [Google Scholar] [CrossRef]
- Sankaranarayanan, R.; Sekar, K.; Banerjee, R.; Sharma, V.; Surolia, A.; Vijayan, M. A novel mode of carbohydrate recognition in jacalin, a Moraceae plant lectin with a β-prism fold. Nat. Struct. Biol. 1996, 3, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-W.; Smith, C.A.; Daily, M.D.; Cort, J.R.; Davin, L.B.; Lewis, N.G. Trimeric structure of (+)-pinoresinol-forming dirigent protein at 1.95 Å resolution with three isolated active sites. J. Biol. Chem. 2015, 290, 1308–1318. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Yamamura, M.; Matsuda, F.; Ono, E.; Nakabayashi, R.; Sugawara, S.; Mori, T.; Tobimatsu, Y.; Umezawa, T.; Saito, K. Seed-coat protective neolignans are produced by the dirigent protein AtDP1 and the laccase AtLAC5 in Arabidopsis. Plant Cell 2021, 33, 129–152. [Google Scholar] [CrossRef]
- Moore, A.D.; Björklund, Å.K.; Ekman, D.; Bornberg-Bauer, E.; Elofsson, A. Arrangements in the modular evolution of proteins. Trends Biochem. 2008, 33, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.J.; Ahn, J.-W.; Jung, S.; Hwang, J.E.; Hong, M.J.; Choi, H.-I.; Kim, J.-B. Overexpression of rice jacalin-related mannose-binding lectin (OsJAC1) enhances resistance to ionizing radiation in Arabidopsis. BMC Plant Biol. 2019, 19, 561. [Google Scholar] [CrossRef] [PubMed]
- Gomord, V.; Fitchette, A.C.; Menu-Bouaouiche, L.; Saint-Jore-Dupas, C.; Plasson, C.; Michaud, D.; Faye, L. Plant-specific glycosylation patterns in the context of therapeutic protein production. Plant Biotechnol. J. 2010, 8, 564–587. [Google Scholar] [CrossRef]
- Kim, S.S.; Sattely, E.S. Dirigent Proteins Guide Asymmetric Heterocoupling for the Synthesis of Complex Natural Product Analogues. J. Am. Chem. Soc. 2021, 143, 5011–5021. [Google Scholar] [CrossRef]
- Kaku, H.; Nishizawa, Y.; Ishii-Minami, N.; Akimoto-Tomiyama, C.; Dohmae, N.; Takio, K.; Minami, E.; Shibuya, N. Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Nat. Acad. Sci. USA 2006, 103, 11086–11091. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen | Protein Buffer | Protein Stock Concentration [mg/mL] | Reservoir Solution | Protein-Tag |
|---|---|---|---|---|
| JRL-cf1 | 25 mM sodium phosphate, pH 7.0 | 10 | 0.05 M MES pH 6.0, 28% (w/v) PEG 3350 | His |
| JRL-cf2 | 25 mM sodium phosphate, pH 7.0 | 11 | 0.2 M NH4I, 20% (w/v) PEG 3350 | Strep |
| DIR-cf1 | 15 mM Tris-HCl, pH 7.4, 1 mM DTT | 6.25 | 0.2 M CaCl2, 20% (w/v) PEG 3350 | none a |
| DIR-cf2 | 15 mM Tris-HCl, pH 7.4, 1 mM DTT | 6.25 | 0.2 M ammonium dihydrogen phosphate; 0.1 M Tris-HCl pH 8.5; 50% (v/v) MPD | none a |
| Specimen | OsJAC1-DIR (cf1) | OsJAC1-DIR (cf2) | OsJAC1-DIR (cf2) w/Galactobiose | OsJAC1-JRL (cf1) | OsJAC1-JRL (cf2) |
|---|---|---|---|---|---|
| PDB code | 7R5Z | 7YWE | 7YWF | 7YWG | 7YWW |
| Beamline | ID30A-3 (ESRF) | ID30A-3 (ESRF) | ID23-1 (ESRF) | ID23-1 (ESRF) | ID29 (ESRF) |
| Detector | EIGER 4M | EIGER 4M | EIGER2 16M | PILATUS 6M | PILATUS 6M |
| Data collection statistics | |||||
| Wavelength (Å) | 0.968 | 0.968 | 0.775 | 0.979 | 0.919 |
| Space group | P 21 21 21 | H 3 (R3:H) | H 3 (R3:H) | P 21 21 21 | C 2 2 21 |
| Unit cell | |||||
| a, b, c (Å) | 49.0, 87.4, 93.7 | 158.9, 158.9, 47.9 | 158.1, 158.1, 48.7 | 34.9, 63.5, 105.9 | 80.3, 99.6, 93.7 |
| α, β, γ (°) | 90.0, 90.0, 90.0 | 90.0, 90.0, 120.0 | 90.0, 90.0, 120.0 | 90.0, 90.0, 90.0 | 90.0, 90.0, 90.0 |
| Resolution (Å) | 43.68–1.75 (1.80–1.75) | 45.87–2.15 (2.21–2.15) | 45.90–2.60 (2.67–2.60) | 54.46–1.10 (1.13–1.10) | 52.02–1.40 (1.44–1.40) |
| Wilson B-factor (Ų) | 31.9 | 60.8 | 86.5 | 13.3 | 22.4 |
| No. reflections | 41,085 | 24,425 | 13,939 | 87,556 | 72,055 |
| Multiplicity | 13.7 (13.2) | 4.2 (4.4) | 5.7 (5.8) | 2.3 (1.4) | 3.74 (3.72) |
| Completeness (%) | 99.4 (97.6) | 99.7 (99.9) | 99.9 (100.0) | 90.9 (56.7) | 92.5 (96.6) |
| I/σI | 10.11 (0.69) | 7.83 (0.68) | 6.02 (0.51) | 5.45 (0.85) | 8.80 (0.65) |
| CC1/2 (%) | 99.8 (34.6) | 99.8 (35.3) | 99.5 (38.1) | 98.8 (61.2) | 99.9 (42.9) |
| Rmeas (%) | 13.9 (439.7) | 12.4 (276.8) | 20.4 (325.6) | 10.4 (73.0) | 4.9 (117.1) |
| Refinement statistics | |||||
| No. reflections | 41,050 | 24,347 | 13,912 | 87,550 | 71,955 |
| Rwork (%) | 19.10 | 21.78 | 24.24 | 15.51 | 16.87 |
| Rfree (%) | 22.90 | 25.44 | 27.77 | 18.80 | 19.46 |
| RMSD bonds (Å) | 0.006 | 0.002 | 0.002 | 0.007 | 0.013 |
| RMSD angles (°) | 0.863 | 0.460 | 0.567 | 1.031 | 1.190 |
| <B> [Å2] (no. atoms) | |||||
| Protein | 55.4 (3352) | 80.3 (3204) | 111.4 (3109) | 17.3 (2424) | 28.1 (2368) |
| Water | 49.6 (120) | 68.7 (26) | 97.1 (5) | 31.8 (441) | 40.1 (299) |
| Other | 47.2 (6) | 96.4 (28) | 118.5 (90) | 13.3 (5) | 44.1 (39) |
| Ramachandran plot | |||||
| Favored (%) | 96.27 | 96.24 | 95.07 | 97.22 | 96.25 |
| Allowed (%) | 3.03 | 3.05 | 4.23 | 2.78 | 3.75 |
| Outliers (%) | 0.70 | 0.70 | 0.70 | 0.00 | 0.00 |
| DTT [mM] | Protein | µg/mL |
|---|---|---|
| 4 | OsJAC1 | 0.98 |
| 4 | DIR | 0.98 |
| 4 | JRL | 15.6–7.8 |
| 0 | OsJAC1 | 0.98 |
| 0 | DIR | 0.98 |
| 0 | JRL | 1.95–3.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huwa, N.; Weiergräber, O.H.; Fejzagić, A.V.; Kirsch, C.; Schaffrath, U.; Classen, T. The Crystal Structure of the Defense Conferring Rice Protein OsJAC1 Reveals a Carbohydrate Binding Site on the Dirigent-like Domain. Biomolecules 2022, 12, 1126. https://doi.org/10.3390/biom12081126
Huwa N, Weiergräber OH, Fejzagić AV, Kirsch C, Schaffrath U, Classen T. The Crystal Structure of the Defense Conferring Rice Protein OsJAC1 Reveals a Carbohydrate Binding Site on the Dirigent-like Domain. Biomolecules. 2022; 12(8):1126. https://doi.org/10.3390/biom12081126
Chicago/Turabian StyleHuwa, Nikolai, Oliver H. Weiergräber, Alexander V. Fejzagić, Christian Kirsch, Ulrich Schaffrath, and Thomas Classen. 2022. "The Crystal Structure of the Defense Conferring Rice Protein OsJAC1 Reveals a Carbohydrate Binding Site on the Dirigent-like Domain" Biomolecules 12, no. 8: 1126. https://doi.org/10.3390/biom12081126
APA StyleHuwa, N., Weiergräber, O. H., Fejzagić, A. V., Kirsch, C., Schaffrath, U., & Classen, T. (2022). The Crystal Structure of the Defense Conferring Rice Protein OsJAC1 Reveals a Carbohydrate Binding Site on the Dirigent-like Domain. Biomolecules, 12(8), 1126. https://doi.org/10.3390/biom12081126