
Analysis of Muscle Lipidome in Juvenile Rainbow Trout Fed Rapeseed Oil and Cochayuyo Meal

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Handling and Diets
2.2. Extraction of the Lipid Phase from the Fillet
2.3. LC-MS Data Acquisition
2.4. Data Analysis
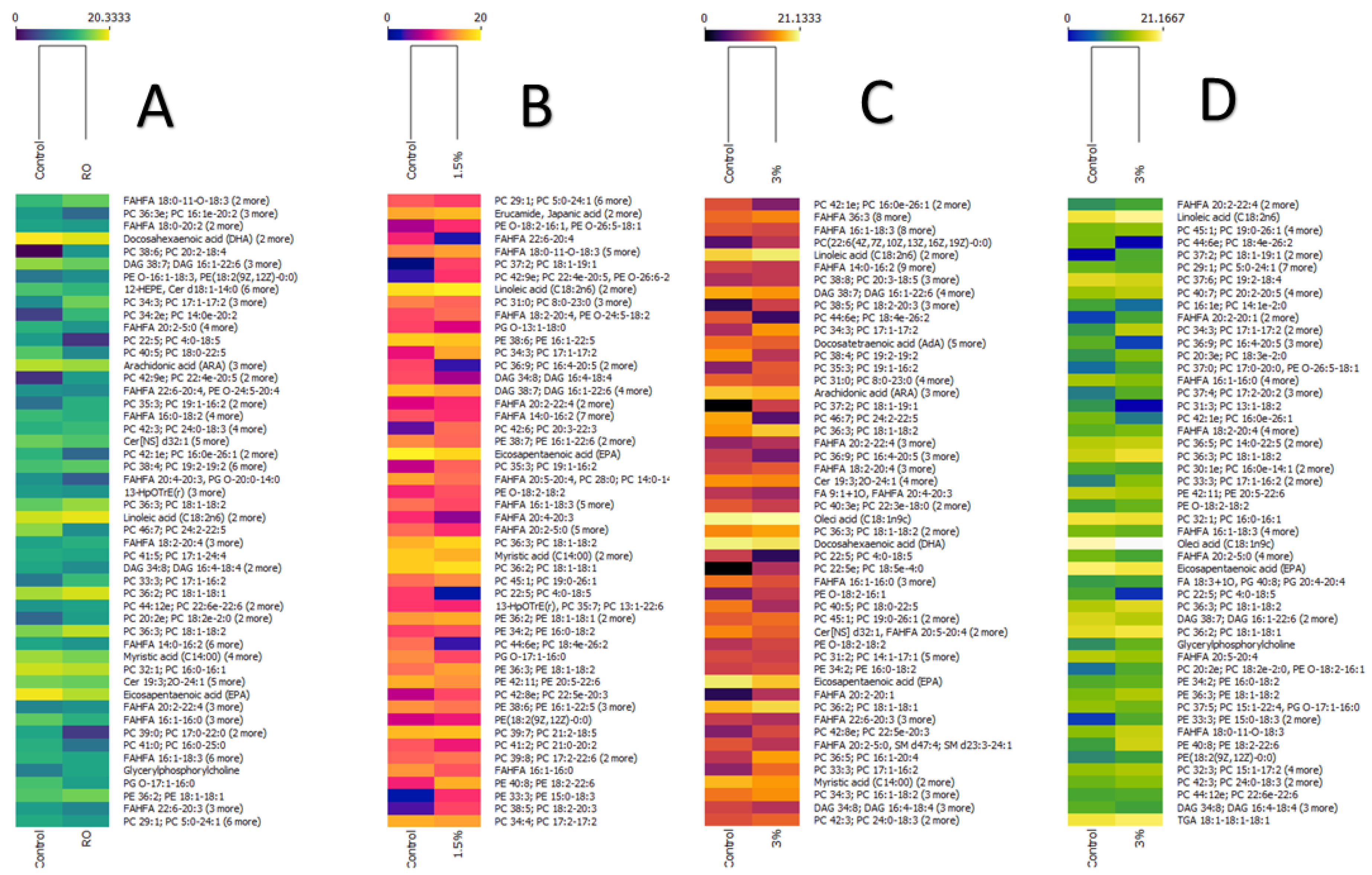
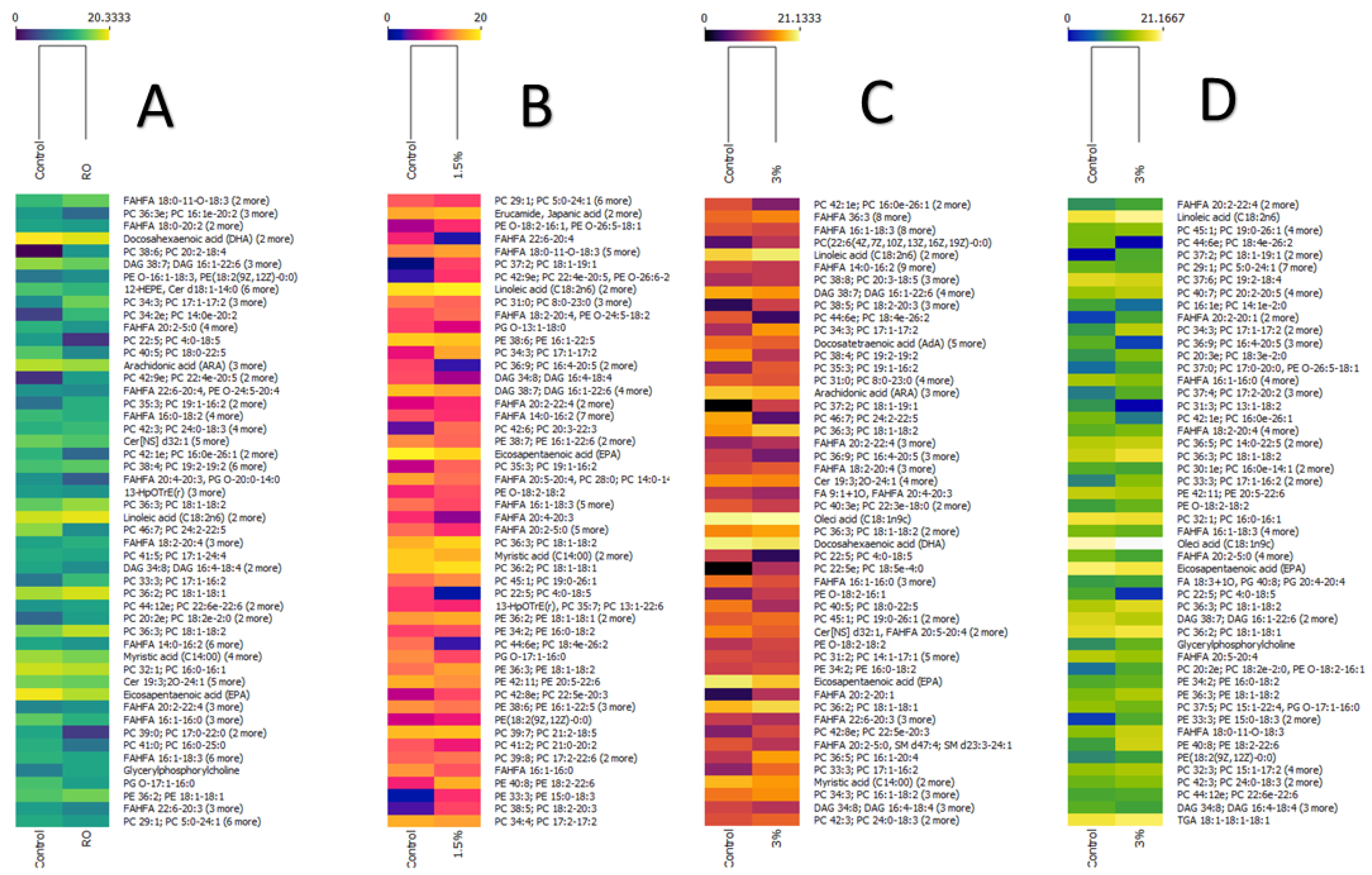
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Kromhout, D.; Bosschieter, E.B.; Coulander, C.D. The inverse relation between fish consumption and 20-year mortality from coronary heart disease. N. Engl. J. Med. 1985, 312, 1205–1209. [Google Scholar] [CrossRef] [PubMed]
- Streppel, M.T.; Ocké, M.C.; Boshuizen, H.C.; Kok, F.J.; Kromhout, D. Long-term fish consumption and n-3 fatty acid intake in relation to (sudden) coronary heart disease death: The Zutphen study. Eur. Heart J. 2008, 29, 2024–2030. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; Fisheries and Aquaculture Department: Rome, Italy, 2018. [Google Scholar]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-L.; Zhao, W.; Hu, W.-S.; Zhu, B.; Xie, J.-J.; Liu, Y.-J.; Tian, L.-X.; Niu, J. Lipid metabolism, growth performance, antioxidant ability and intestinal morphology of rainbow trout (Oncorhynchus mykiss) under cage culture with flowing water were affected by dietary lipid levels. Aquac. Rep. 2021, 19, 100593. [Google Scholar] [CrossRef]
- Turchini, G.; Hermon, K.M.; Francis, D. Fatty acids and beyond: Fillet nutritional characterisation of rainbow trout (Oncorhynchus mykiss) fed different dietary oil sources. Aquaculture 2018, 491, 391–397. [Google Scholar] [CrossRef]
- Caballero, M.; Obach, A.; Rosenlund, G.; Montero, D.; Gisvold, M.; Izquierdo, M. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss. Aquaculture 2002, 214, 253–271. [Google Scholar] [CrossRef]
- Hossain, S.; Peng, M.; Small, B.C. Optimizing the fatty acid profile of novel terrestrial oil blends in low fishmeal diets of rainbow trout (Oncorhynchus mykiss) yields comparable fish growth, total fillet n-3 LC-PUFA content, and health performance relative to fish oil. Aquaculture 2021, 545, 737230. [Google Scholar] [CrossRef]
- Pettersson, A.; Johnsson, L.; Brännäs, E.; Pickova, J. Effects of rapeseed oil replacement in fish feed on lipid composition and self-selection by rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2009, 15, 577–586. [Google Scholar] [CrossRef]
- Dantagnan, P.; Hernã¡ndez, A.; Borquez, A.; Mansilla, A. Inclusion of macroalgae meal (Macrocystis pyrifera) as feed ingredient for rainbow trout (Oncorhynchus mykiss): Effect on flesh fatty acid composition. Aquac. Res. 2009, 41, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Quiñones, J.; Díaz, R.; Dantagnan, P.; Hernández, A.; Valdes, M.; Lorenzo, J.M.; Cancino, D.; Sepúlveda, N.; Farías, J.G. Dietary inclusion of Durvillaea antarctica meal and rapeseed (Brassica napus) oil on growth, feed utilization and fillet quality of rainbow trout (Oncorhynchus mykiss). Aquaculture 2020, 530, 735882. [Google Scholar] [CrossRef]
- Kyle, J.E.; Aimo, L.; Bridge, A.J.; Clair, G.; Fedorova, M.; Helms, J.B.; Molenaar, M.R.; Ni, Z.; Orešič, M.; Slenter, D.; et al. Interpreting the lipidome: Bioinformatic approaches to embrace the complexity. Metabolomics 2021, 17, 55. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Köfeler, H.; et al. Update on LIPID MAPS classification, nomenclature, and shorthand notation for MS-derived lipid structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Mitina, A.; Mazin, P.; Vanyushkina, A.; Anikanov, N.; Mair, W.; Guo, S.; Khaitovich, P. Lipidome analysis of milk composition in humans, monkeys, bovids, and pigs. BMC Evol. Biol. 2020, 20, 70. [Google Scholar] [CrossRef] [PubMed]
- Venkata, A.R.; Ramesh, M. A Concise Review on Lipidomics Analysis in Biological Samples. ADMET DMPK 2021, 9, 1–22. [Google Scholar] [CrossRef]
- Song, G.; Chen, K.; Wang, H.; Zhang, M.; Yu, X.; Wang, J.; Shen, Q. In situ and real-time authentication of Thunnus species by iKnife rapid evaporative ionization mass spectrometry based lipidomics without sample pretreatment. Food Chem. 2020, 318, 126504. [Google Scholar] [CrossRef]
- Bell, J.; McGhee, F.; Campbell, P.J.; Sargent, J.R. Rapeseed oil as an alternative to marine fish oil in diets of post-smolt Atlantic salmon (Salmo salar): Changes in flesh fatty acid composition and effectiveness of subsequent fish oil “wash out”. Aquaculture 2003, 218, 515–528. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Wang, X.; Cong, P.; Xu, J.; Xue, C. Mass spectrometry-based lipidomics in food science and nutritional health: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2530–2558. [Google Scholar] [CrossRef]
- Liu, Y.; Jiao, J.-G.; Gao, S.; Ning, L.-J.; Limbu, S.M.; Qiao, F.; Chen, L.-Q.; Zhang, M.-L.; Du, Z.-Y. Dietary oils modify lipid molecules and nutritional value of fillet in Nile tilapia: A deep lipidomics analysis. Food Chem. 2019, 277, 515–523. [Google Scholar] [CrossRef]
- Jin, Y.; Harvey, T.N.; Bartosova, Z.; Hassani, S.; Bruheim, P.; Sandve, S.R.; Vik, J.O. Diet and Life Stage-Associated Lipidome Remodeling in Atlantic Salmon. J. Agric. Food Chem. 2021, 69, 3787–3796. [Google Scholar] [CrossRef]
- Yu, X.; Li, L.; Wang, H.; Song, G.; Wang, J.; Li, S.; Wang, Y.; Shen, Q. Lipidomics study of rainbow trout (Oncorhynchus mykiss) and salmons (Oncorhynchus tshawytscha and Salmo salar) using hydrophilic interaction chromatography and mass spectrometry. LWT 2019, 121, 108988. [Google Scholar] [CrossRef]
- Weil, C.; Lefèvre, F.; Bugeon, J. Characteristics and metabolism of different adipose tissues in fish. Rev. Fish Biol. Fish. 2013, 23, 157–173. [Google Scholar] [CrossRef]
- Kostyniuk, D.J.; Culbert, B.M.; Mennigen, J.A.; Gilmour, K.M. Social status affects lipid metabolism in rainbow trout, oncorhynchus mykiss. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2018, 315, R241–R255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Liao, K.; Mai, K.; Ai, Q. Dietary lipid levels affect lipoprotein clearance, fatty acid transport, lipogenesis and lipolysis at the transcriptional level in muscle and adipose tissue of large yellow croaker (Larimichthys crocea). Aquac. Res. 2017, 48, 3925–3934. [Google Scholar] [CrossRef]
- Panserat, S.; Ducasse-Cabanot, S.; Plagnes-Juan, E.; Srivastava, P.; Kolditz, C.; Piumi, F.; Esquerré, D.; Kaushik, S. Dietary fat level modifies the expression of hepatic genes in juvenile rainbow trout (Oncorhynchus mykiss) as revealed by microarray analysis. Aquaculture 2008, 275, 235–241. [Google Scholar] [CrossRef]
- Ayisi, C.L.; Yamei, C.; Zhao, J.L. Genes, transcription factors and enzymes involved in lipid metabolism in fin fish. Agric. Gene 2018, 7, 7–14. [Google Scholar] [CrossRef]
- Olsen, R.E.; Dragnes, B.T.; Myklebust, R.; Ringø, E. Effect of soybean oil and soybean lecithin on intestinal lipid composition and lipid droplet accumulation of rainbow trout, Oncorhynchus mykiss Walbaum. Fish Physiol. Biochem. 2003, 29, 181–192. [Google Scholar] [CrossRef]
- Wood, P.L. Fatty Acyl Esters of Hydroxy Fatty Acid (FAHFA) Lipid Families. Metabolites 2020, 10, 512. [Google Scholar] [CrossRef]
- Coudray, C.; Durand, E.; Balas, L.; Sultan, A.; Casas, F.; Feillet-Coudray, C. Potential favourable health effects of some dietary uncommon fatty acids. OCL Oilseeds Fats Crops Lipids 2021, 28, 9p. [Google Scholar] [CrossRef]
- Riecan, M.; Paluchova, V.; Lopes, M.; Brejchova, K.; Kuda, O. Branched and linear fatty acid esters of hydroxy fatty acids (FAHFA) relevant to human health. Pharmacol. Ther. 2021, 231, 107972. [Google Scholar] [CrossRef]
- Jackowski, S. Cell Cycle Regulation of Membrane Phospholipid Metabolism. J. Biol. Chem. 1996, 271, 20219–20222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurray, W.C.; Magee, W.L. Phospholipid metabolism. Annu. Rev. Biochem. 1972, 41, 129–160. [Google Scholar] [CrossRef] [PubMed]
- Fagone, P.; Jackowski, S. Membrane phospholipid synthesis and endoplasmic reticulum function. J. Lipid Res. 2009, 50, S311–S316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tocher, D.R.; Bendiksen, E.Å.; Campbell, P.J.; Bell, J.G. The role of phospholipids in nutrition and metabolism of teleost fish. Aquaculture 2008, 280, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Katavic, V.; Agrawal, G.K.; Hajduch, M.; Harris, S.L.; Thelen, J.J. Protein and lipid composition analysis of oil bodies from twoBrassica napus cultivars. Proteomics 2006, 6, 4586–4598. [Google Scholar] [CrossRef]
- Riera-Heredia, N.; Lutfi, E.; Sánchez-Moya, A.; Gutiérrez, J.; Capilla, E.; Navarro, I. Short-Term Responses to Fatty Acids on Lipid Metabolism and Adipogenesis in Rainbow Trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2020, 21, 1623. [Google Scholar] [CrossRef] [Green Version]
- Maulucci, G.; Cohen, O.; Daniel, B.; Sansone, A.; Petropoulou, P.I.; Filou, S.; Spyridonidis, A.; Pani, G.; De Spirito, M.; Chatgilialoglu, C.; et al. Fatty acid-related modulations of membrane fluidity in cells: Detection and implications. Free Radic. Res. 2016, 50, S40–S50. [Google Scholar] [CrossRef] [Green Version]
- Hazel, J.R. Influence of thermal acclimation on membrane lipid composition of rainbow trout liver. Am. J. Physiol. Integr. Comp. Physiol. 1979, 236, R91–R101. [Google Scholar] [CrossRef] [Green Version]
- Cottin, S.C.; Sanders, T.; Hall, W. The differential effects of EPA and DHA on cardiovascular risk factors. Proc. Nutr. Soc. 2011, 70, 215–231. [Google Scholar] [CrossRef] [Green Version]
- Monroig, Ó.; Navarro, J.C.; Tocher, D.R. Long-Chain Polyunsaturated Fatty Acids in Fish: Recent Advances on Desaturases and Elongases Involved in Their Biosynthesis. Av. En Nutr. Acuicola 2011, 1, 257–283. Available online: https://nutricionacuicola.uanl.mx/index.php/acu/article/view/93 (accessed on 14 November 2021).
- Sprecher, H. Metabolism of highly unsaturated n-3 and n-6 fatty acids. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2000, 1486, 219–231. [Google Scholar] [CrossRef]
- Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; Lopez-Hernandez, J.; Bozzo, C.; Navarrete, E.; Osorio, A.; Rios, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef]
- Wan, A.H.L.; Davies, S.J.; Soler-Vila, A.; Fitzgerald, R.; Johnson, M.P. Macroalgae as a sustainable aquafeed ingredient. Rev. Aquac. 2019, 11, 458–492. [Google Scholar] [CrossRef]
- Bruni, L.; Secci, G.; Mancini, S.; Faccenda, F.; Parisi, G. A commercial macroalgae extract in a plant-protein rich diet diminished saturated fatty acids of Oncorhynchus mykiss walbaum fillets. Ital. J. Anim. Sci. 2020, 19, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Güroy, B.; Ergün, S.; Merrifield, D.L.; Güroy, D. Effect of autoclaved Ulva meal on growth performance, nutrient utilization and fatty acid profile of rainbow trout, Oncorhynchus mykiss. Aquac. Int. 2012, 21, 605–615. [Google Scholar] [CrossRef]
- Caruso, D.; Risé, P.; Galella, G.; Regazzoni, C.; Toia, A.; Galli, G.; Galli, C. Formation of 22 and 24 carbon 6-desaturated fatty acids from exogenous deuterated arachidonic acid is activated in THP-1 cells at high substrate concentrations. FEBS Lett. 1994, 343, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Safavi, S.V.; Kenari, A.A.; Tabarsa, M.; Esmaeili, M. Effect of sulfated polysaccharides extracted from marine macroalgae (Ulva intestinalis and Gracilariopsis persica) on growth performance, fatty acid profile, and immune response of rainbow trout (Oncorhynchus mykiss). J. Appl. Phycol. 2019, 31, 4021–4035. [Google Scholar] [CrossRef]
- Niu, J.; Li, C.-H.; Tian, L.-X.; Liu, Y.-J.; Chen, X.; Wu, K.-C.; Jun, W.; Huang, Z.; Wang, Y.; Lin, H.-Z. Suitable dietary chitosan improves the growth performance, survival and immune function of tiger shrimp, Penaeus monodon. Aquac. Res. 2013, 46, 1668–1678. [Google Scholar] [CrossRef]
- Qin, L.; Xu, H.; He, Y.; Liang, C.; Wang, K.; Cao, J.; Qu, C.; Miao, J. Purification, Chemical Characterization and Immunomodulatory Activity of a Sulfated Polysaccharide from Marine Brown Algae Durvillaea antarctica. Mar. Drugs 2022, 20, 223. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, X.; Li, J.; Jiang, H.; Shan, X.; Wang, Y.; Ma, W.; Hao, J.; Yu, G. A β-glucan from Durvillaea Antarctica has immunomodulatory effects on RAW264.7 macrophages via toll-like receptor 4. Carbohydr. Polym. 2018, 191, 255–265. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Ingredients % DM | Control | RO | 1.5% | 3% | 6% |
|---|---|---|---|---|---|
| Fish meal | 100 | 100 | 100 | 100 | 100 |
| Fish oil | 100 | 10 | 10 | 10 | 10 |
| Brassica napus oil | 0 | 90 | 90 | 90 | 90 |
| D. antarctica meal | 0 | 0 | 1.5 | 3 | 6 |
| Cellulose | 5.4 | 5.4 | 4.1 | 2.7 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiñones, J.; Díaz, R.; Beltrán, J.F.; Velazquez, L.; Cancino, D.; Muñoz, E.; Dantagnan, P.; Hernández, A.; Sepúlveda, N.; Farías, J.G. Analysis of Muscle Lipidome in Juvenile Rainbow Trout Fed Rapeseed Oil and Cochayuyo Meal. Biomolecules 2022, 12, 805. https://doi.org/10.3390/biom12060805
Quiñones J, Díaz R, Beltrán JF, Velazquez L, Cancino D, Muñoz E, Dantagnan P, Hernández A, Sepúlveda N, Farías JG. Analysis of Muscle Lipidome in Juvenile Rainbow Trout Fed Rapeseed Oil and Cochayuyo Meal. Biomolecules. 2022; 12(6):805. https://doi.org/10.3390/biom12060805
Chicago/Turabian StyleQuiñones, John, Rommy Díaz, Jorge F. Beltrán, Lidiana Velazquez, David Cancino, Erwin Muñoz, Patricio Dantagnan, Adrián Hernández, Néstor Sepúlveda, and Jorge G. Farías. 2022. "Analysis of Muscle Lipidome in Juvenile Rainbow Trout Fed Rapeseed Oil and Cochayuyo Meal" Biomolecules 12, no. 6: 805. https://doi.org/10.3390/biom12060805
APA StyleQuiñones, J., Díaz, R., Beltrán, J. F., Velazquez, L., Cancino, D., Muñoz, E., Dantagnan, P., Hernández, A., Sepúlveda, N., & Farías, J. G. (2022). Analysis of Muscle Lipidome in Juvenile Rainbow Trout Fed Rapeseed Oil and Cochayuyo Meal. Biomolecules, 12(6), 805. https://doi.org/10.3390/biom12060805