
Star Polymers as Non-Viral Carriers for Apoptosis Induction

 ,
,  , , and
, , and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Synthesis of Star Polymer with Poly(Arylene Oxindole) Core and Poly(N,N′-Dimethylaminoethyl Methacrylate) Arms
2.3. Cytotoxicity Assay
2.4. Fluorescence
2.5. Apoptosis Assay
2.6. Flow Cytometry
2.7. RNA Isolation and Quantitative RT-PCR
2.8. Statistical Analysis
3. Results
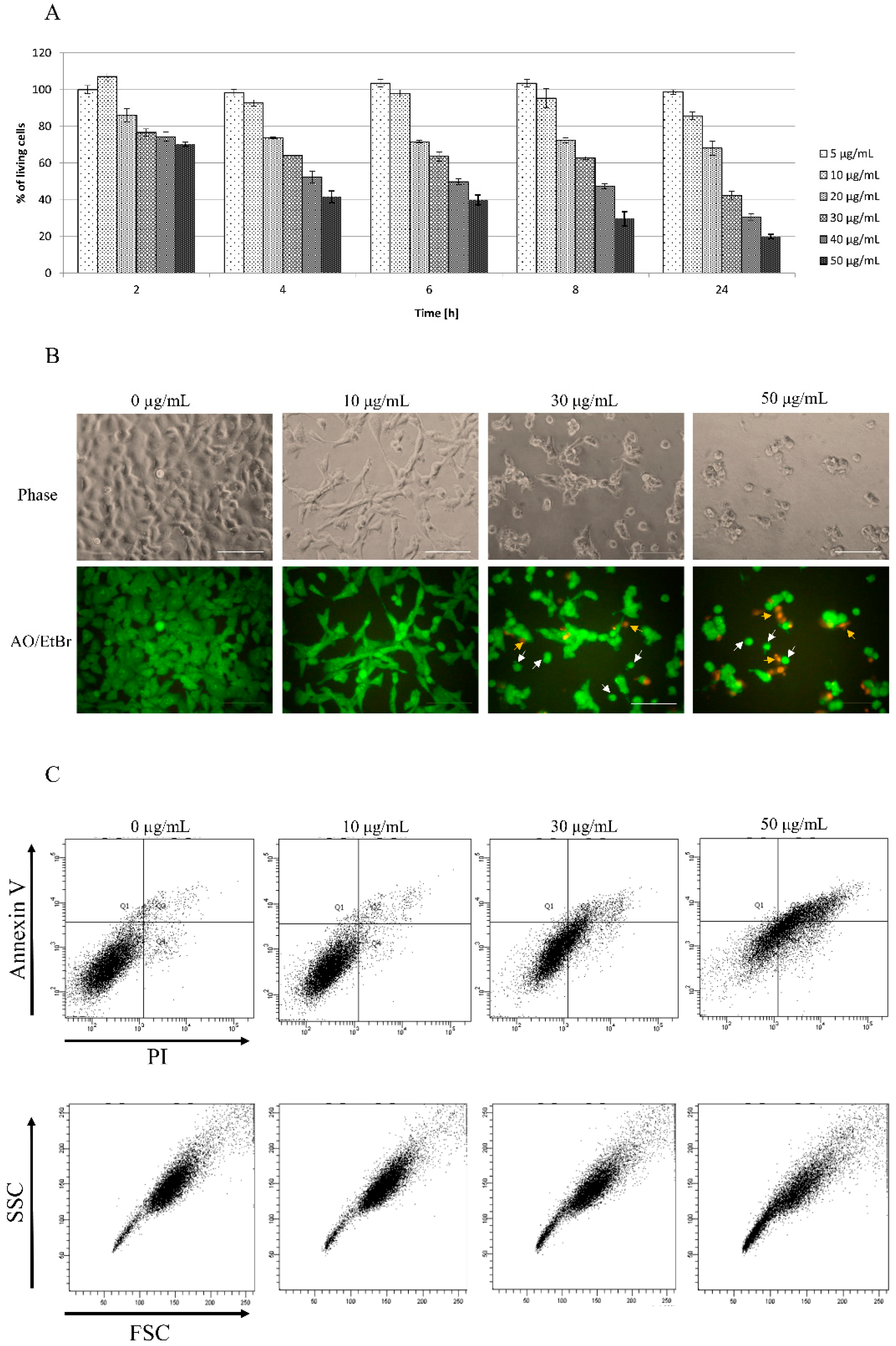
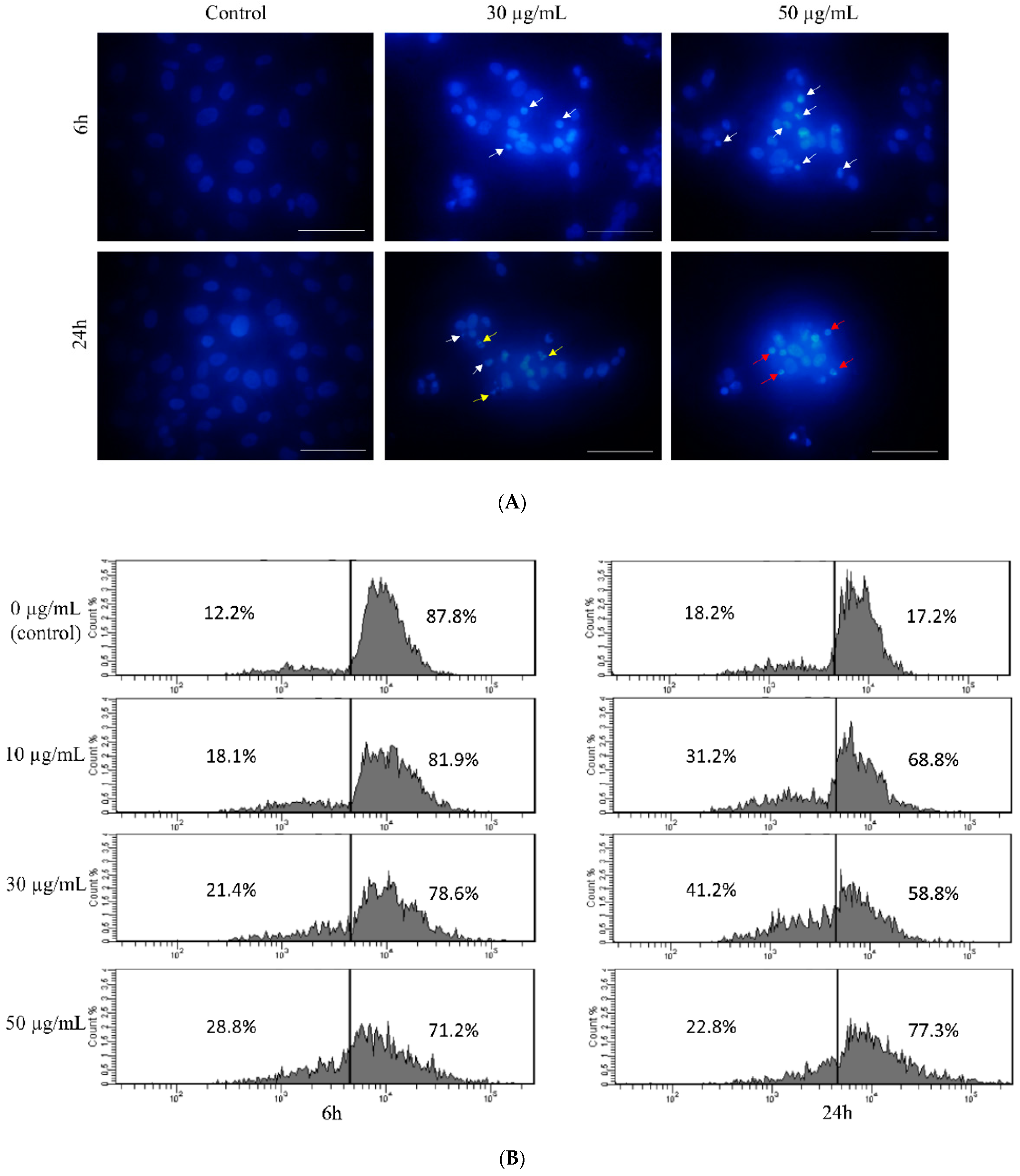
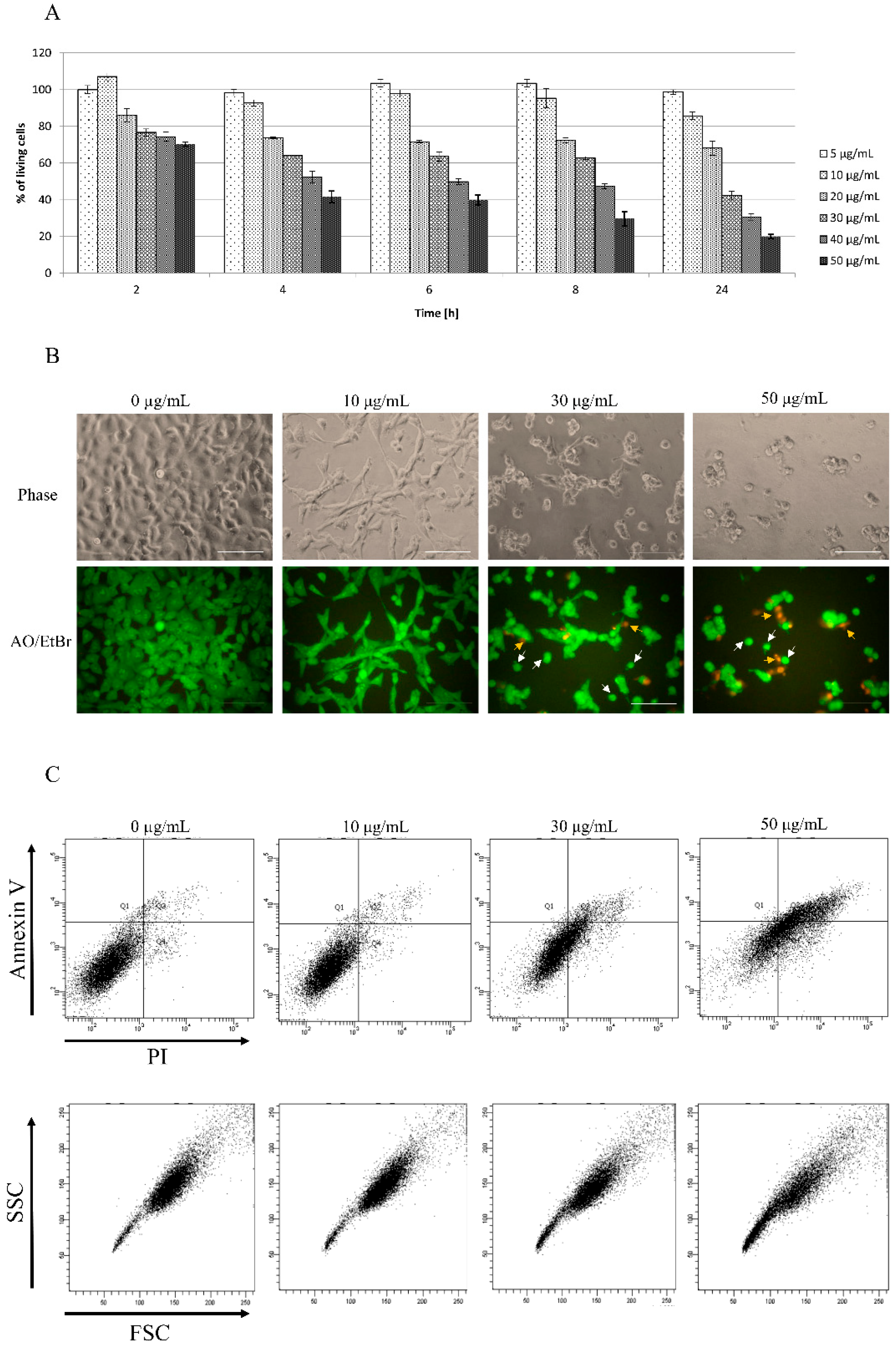
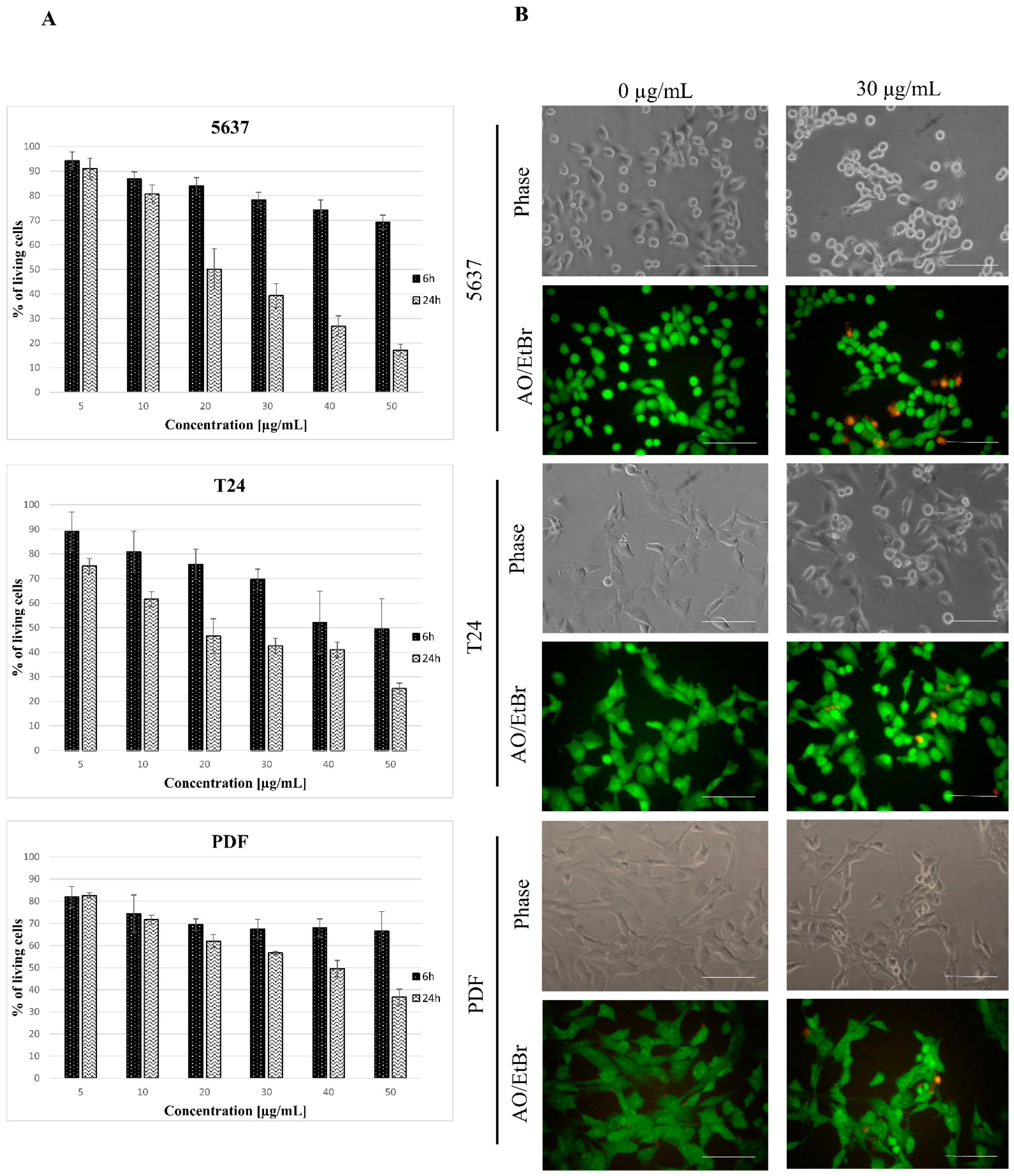
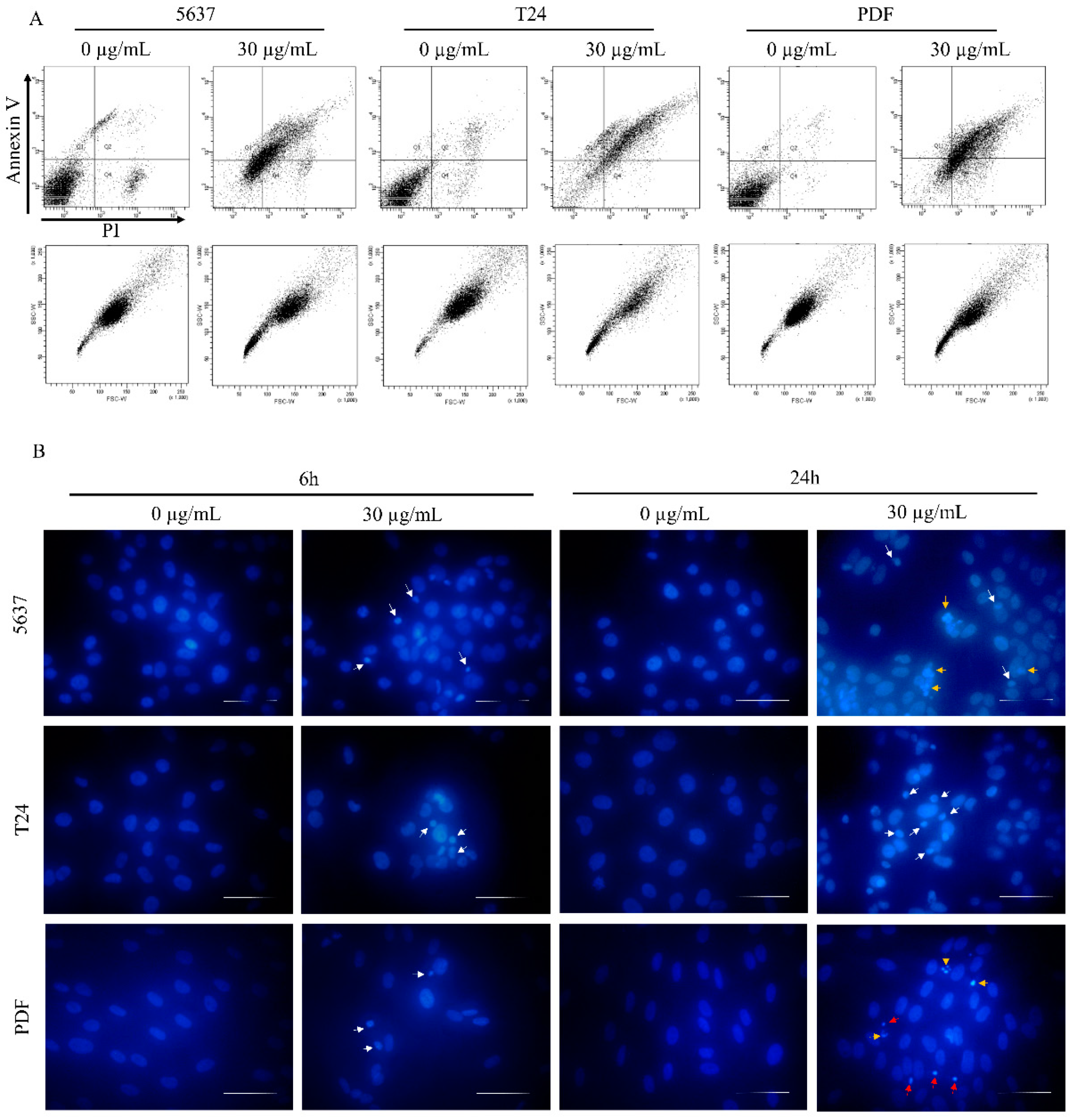
3.1. Toxicity of Star Polymer Is Concentration-Dependent and Induces Mainly Early Apoptosis in HT-1080 Cells
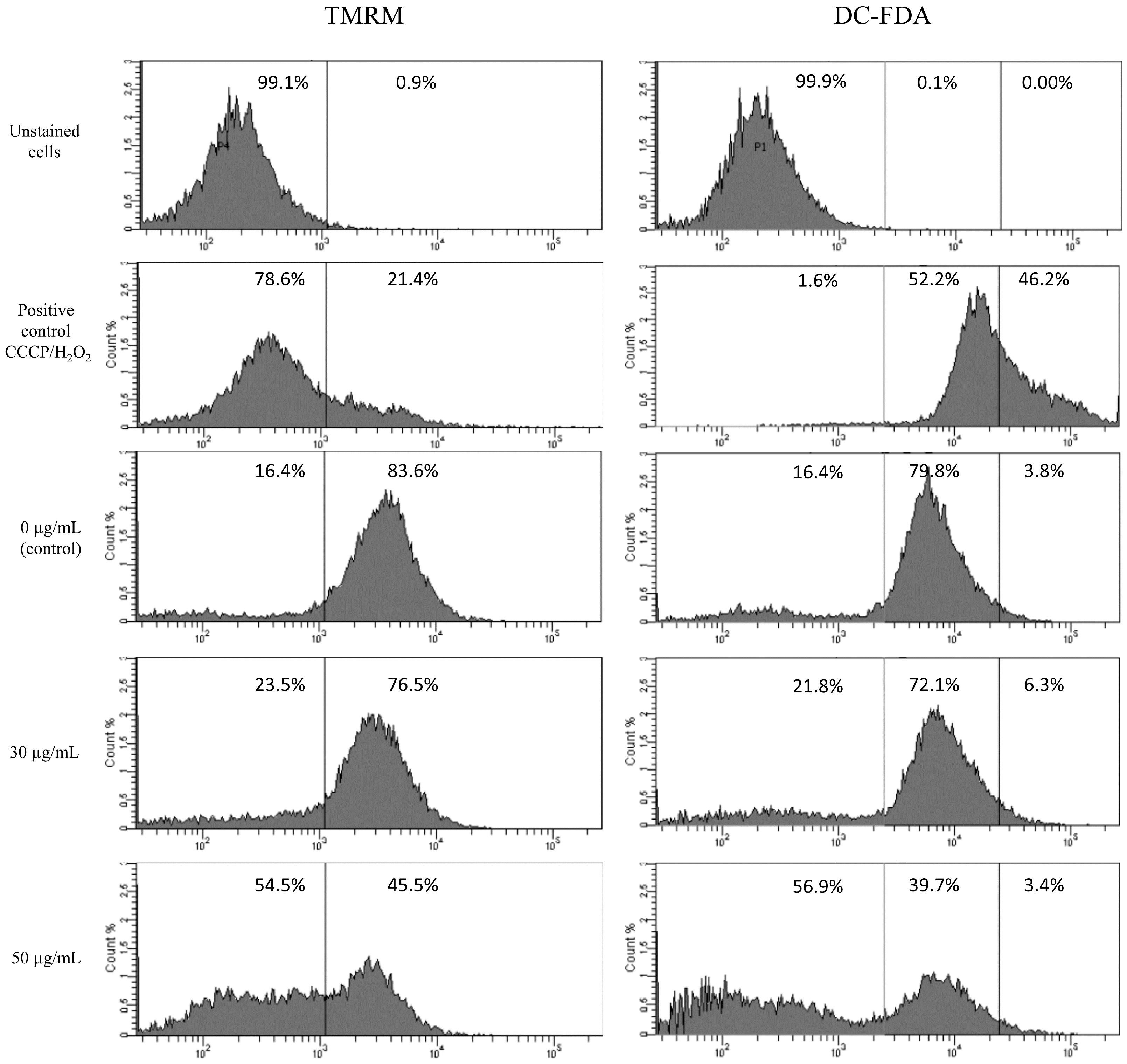
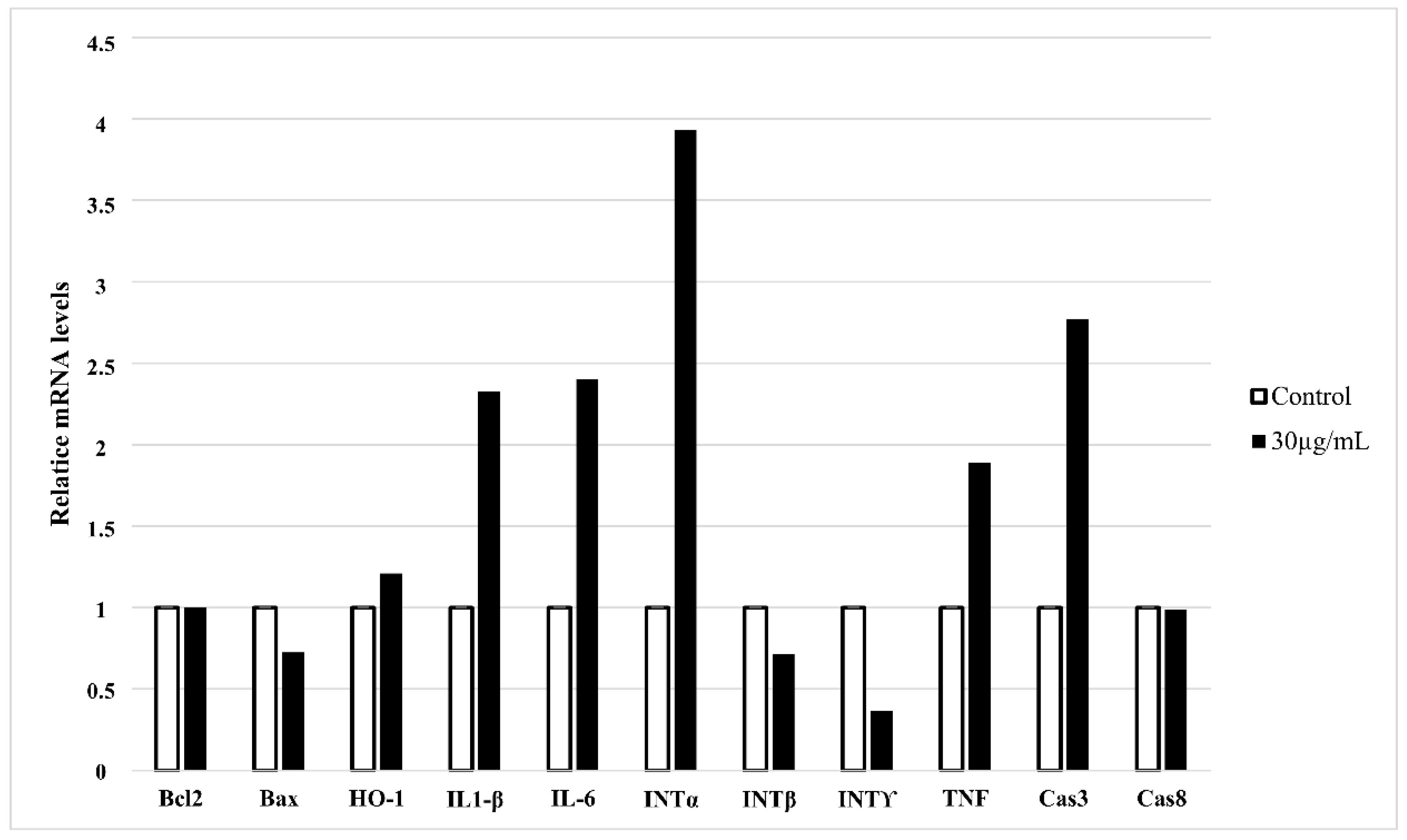
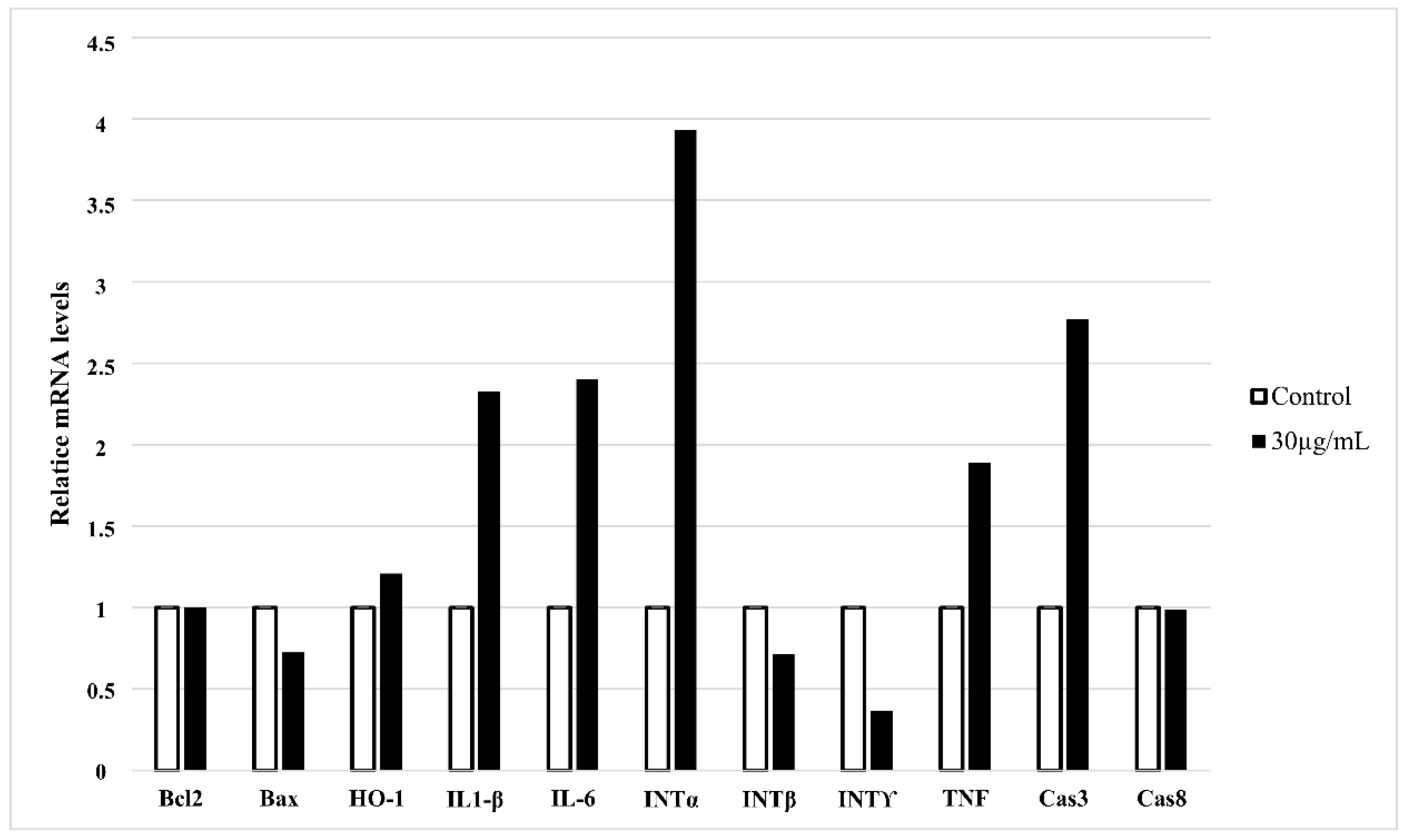
3.2. HT-1080 Cells Treatment with Star Polymer Triggers Changes of Gene Expression Profile and Activates Various Apoptotic Pathways
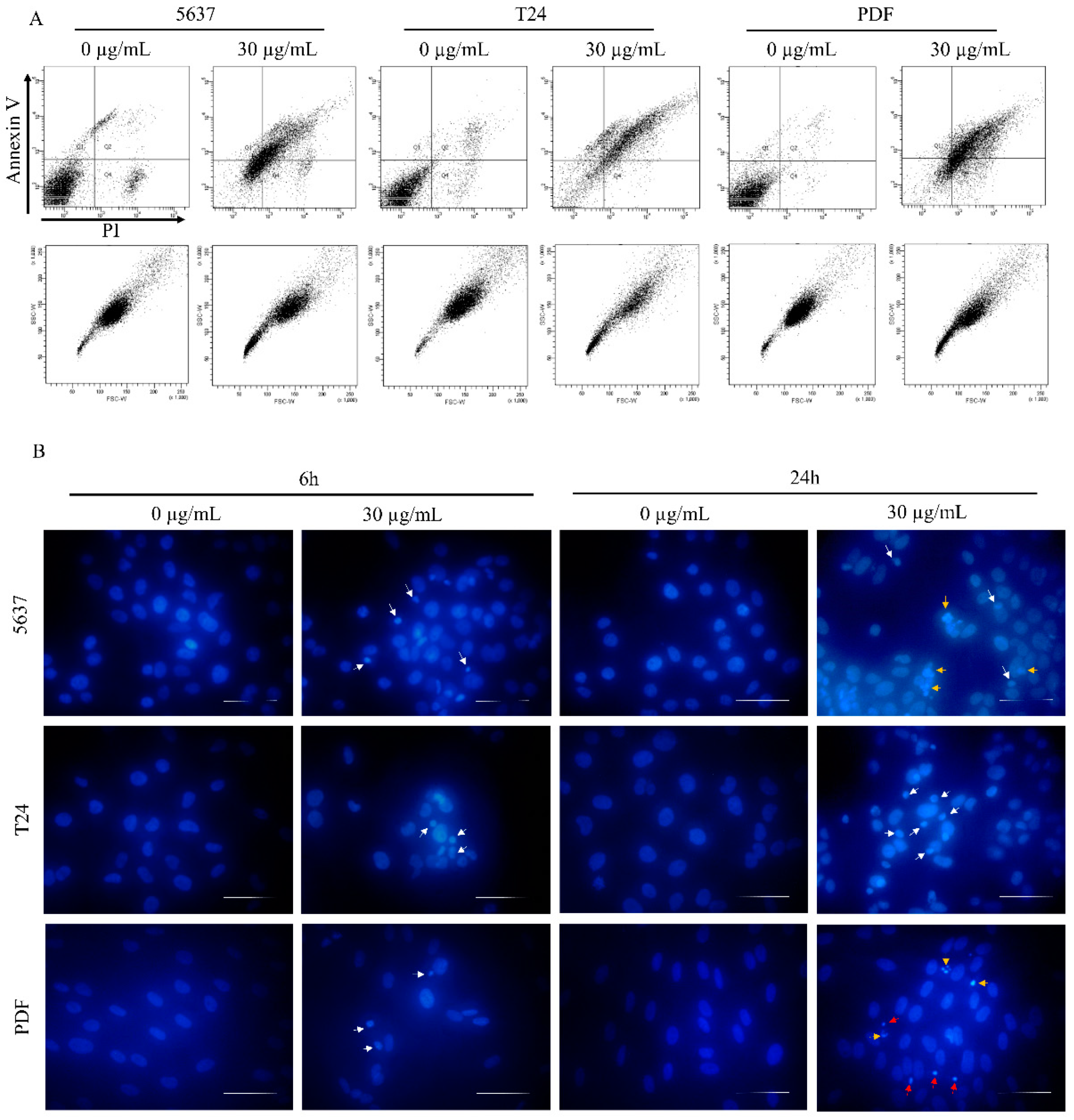
3.3. Star Polymer Causes the Early Apoptosis in Human Bladder Cancer Cell Lines
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, Y.; Steller, H. Programmed cell death in animal development and disease. Cell 2011, 147, 742–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.Y.; Rhee, K. Caspase-mediated cleavage of the centrosomal proteins during apoptosis. Cell Death Dis. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torabi, B.; Flashner, S.; Beishline, K.; Sowash, A.; Donovan, K.; Bassett, G.; Azizkhan-Clifford, J. Caspase cleavage of transcription factor Sp1 enhances apoptosis. Apoptosis 2018, 23, 65–78. [Google Scholar] [CrossRef]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef] [Green Version]
- Fischer, U.; Schulze-Osthoff, K. Apoptosis-based therapies and drug targets. Cell Death Differ. 2005, 12, 942–961. [Google Scholar] [CrossRef]
- Mendrek, B.; Fus, A.; Klarzynska, K.; Sieron, A.L.; Smet, M.; Kowalczuk, A.; Dworak, A. Synthesis, Characterization and Cytotoxicity of Novel Thermoresponsive Star Copolymers of N,N′-Dimethylaminoethyl Methacrylate and Hydroxyl-Bearing Oligo(Ethylene Glycol) Methacrylate. Polymers 2018, 10, 1255. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Li, Z.; Zhu, C.; Xu, L.; Sun, Y.; Han, S. Research progress of nanocarriers for gene therapy targeting abnormal glucose and lipid metabolism in tumors. Drug Deliv. 2021, 28, 2329–2347. [Google Scholar] [CrossRef]
- Siafaka, P.I.; Okur, N.U.; Karavas, E.; Bikiaris, D.N. Surface modified multifunctional and stimuli responsive nanoparticles for drug targeting: Current status and uses. Int. J. Mol. Sci. 2016, 17, 1440. [Google Scholar] [CrossRef]
- Siafaka, P.I.; Okur, N.Ü.; Karantas, I.D.; Okur, M.E.; Gündoğdu, E.A. Current update on nanoplatforms as therapeutic and diagnostic tools: A review for the materials used as nanotheranostics and imaging modalities. Asian J. Pharm. Sci. 2020, 16, 24–46. [Google Scholar] [CrossRef]
- Fus-Kujawa, A.; Prus, P.; Bajdak-Rusinek, K.; Teper, P.; Gawron, K.; Kowalczuk, A.; Sieron, A.L. An Overview of Methods and Tools for Transfection of Eukaryotic Cells in vitro. Front. Bioeng. Biotechnol. 2021, 20, 701031. [Google Scholar] [CrossRef]
- Weissig, V.; Lozoya, M.; Yu, N.; D’Souza, G.G.M. DQAsomes as the Prototype of Mitochondria-Targeted Pharmaceutical Nanocarriers: An Update. Methods Mol. Biol. 2021, 2275, 13–25. [Google Scholar] [CrossRef]
- Georgiou, T.K.; Vamvakaki, M.; Patrickios, C.S.; Yamasaki, E.N.; Phylactou, L.A. Nanoscopic cationic methacrylate star homopolymers: Synthesis by group transfer polymerization, characterization and evaluation as transfection reagents. Biomacromolecules 2004, 5, 2221–2229. [Google Scholar] [CrossRef]
- Wang, C.; Zhai, Z.; Yang, Y.; Wu, Q.; Cheng, Z.; Liang, L.; Dai, H.; Huang, K.; Lu, W.; Zhang, Z. Efficacy and safety of low dose recombinant tissue-type plasminogen activator for the treatment of acute pulmonary thromboembolism: A randomized, multicenter, controlled trial. Chest 2010, 137, 254–262. [Google Scholar] [CrossRef]
- Mendrek, B.; Sieron, L.; Libera, M.; Smet, M.; Trzebicka, B.; Sieron, A.L.; Dworak, A.; Kowalczuk, A. Polycationic star polymers with hyperbranched cores for gene delivery. Polymer 2014, 55, 4551–4562. [Google Scholar] [CrossRef]
- Mendrek, B.; Sieron, L.; Zymełka-Miara, I.; Binkiewicz, P.; Libera, M.; Smet, M.; Trzebicka, B.; Sieron, A.L.; Kowalczuk, A. Nonviral Plasmid DNA Carriers Based on N,N′-Dimethylaminoethyl Methacrylate and Di(ethylene glycol) Methyl Ether Methacrylate Star Copolymers. Biomacromolecules 2015, 16, 3275–3285. [Google Scholar] [CrossRef]
- Fus-Kujawa, A.; Teper, P.; Botor, M.; Klarzynska, K.; Sieron, L.; Verbelen, B.; Smet, M.; Sieron, A.L.; Mendrek, B.; Kowalczuk, A. Functional star polymers as reagents for efficient nucleic acids delivery into HT-1080 cells. Int. J. Polym. Mater. Polym. Biomater. 2021, 70, 356–370. [Google Scholar] [CrossRef]
- Riccardi, C.; Nicoletti, I. Analysis of apoptosis by propidium iodide staining and flow cytometry. Nat. Protoc. 2006, 1, 1458–1461. [Google Scholar] [CrossRef]
- Lächelt, U.; Wagner, E. Nucleic Acid Therapeutics Using Polyplexes: A Journey of 50 Years (and Beyond). Chem. Rev. 2015, 115, 11043–11078. [Google Scholar] [CrossRef]
- Jiang, C.; Chen, J.; Li, Z.; Wang, Z.; Zhang, W.; Liu, J. Recent advances in the development of polyethylenimine-based gene vectors for safe and efficient gene delivery. Exp. Opin. Drug Deliv. 2019, 16, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Makowska, A.; Wahab, L.; Kapetanakis, N.-I.; Vokuhl, C.; Denecke, B.; Shen, L.; Busson, P.; Kontny, U. Interferon Beta Induces Apoptosis in Nasopharyngeal Carcinoma Cells via the TRAIL-Signaling Pathway. Oncotarget 2018, 9, 14228–14250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.; Chen, H.D.; Yu, Y.W.; Li, N.; Chen, W.Q. Changing profiles of cancer burden worldwide and in China: A secondary analysis of the global cancer statistics. Chin. Med. J. 2020, 134, 783–791. [Google Scholar] [CrossRef]
- Scialò, F.; Fernández-Ayala, D.J.; Sanz, A. Role of mitochondrial reverse electron transport in ROS signaling: Potential roles in health and disease. Front. Physiol. 2017, 8, 428. [Google Scholar] [CrossRef]
- Gai, C.; Liu, C.; Wu, X.; Yu, M.; Zheng, J.; Zhang, W.; Lv, S.; Li, W. MT1DP loaded by folate-modified liposomes sensitizes erastin-induced ferroptosis via regulating miR-365a-3p/NRF2 axis in non-small cell lung cancer cells. Cell Death Dis. 2020, 14, 751. [Google Scholar] [CrossRef]
- Khdair, A.; Hamad, I.; Alkhatib, H.; Bustanji, Y.; Mohammad, M.; Tayem, R.; Aiedeh, K. Modified-chitosan nanoparticles: Novel drug delivery systems improve oral bioavailability of doxorubicin. Eur. J. Pharm. Sci. 2016, 10, 38–44. [Google Scholar] [CrossRef]
- Moshikur, R.M.; Ali, M.K.; Wakabayashi, R.; Moniruzzaman, M.; Goto, M. Methotrexate-based ionic liquid as a potent anticancer drug for oral delivery: In vivo pharmacokinetics, biodistribution, and antitumor efficacy. Int. J. Pharm. 2021, 25, 121129. [Google Scholar] [CrossRef]
- Cai, J.; Yue, Y.; Rui, D.; Zhang, Y.; Liu, S.; Wu, C. Effect of chain length on cytotoxicity and endocytosis of cationic polymers. Macromolecules 2011, 44, 2050–2057. [Google Scholar] [CrossRef]
- Xuan, Y.; Yang, Y.; Xiang, L.; Zhang, C. The Role of Oxidative Stress in the Pathogenesis of Vitiligo: A Culprit for Melanocyte Death. Oxid. Med. Cell. Longev. 2022, 2022, 8498472. [Google Scholar] [CrossRef]
- Zorov, D.B.; Bannikova, S.Y.; Belousov, V.V.; Vyssokikh, M.Y.; Zorova, L.D.; Isaev, N.K.; Krasnikov, B.F.; Plotnikov, E.Y. Reactive Oxygen and Nitrogen Species: Friends or Foes? Biochemistry 2005, 70, 215–221. [Google Scholar] [CrossRef]
- Jäättelä, M. Multiple cell death pathways as regulators of tumour initiation and progression. Oncogene 2004, 23, 2746–2756. [Google Scholar] [CrossRef] [Green Version]
- Kotredes, K.P.; Gamero, A.M. Interferons as inducers of apoptosis in malignant cells. J. Interf Cytokine Res. 2013, 33, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Jarry, A.; Malard, F.; Bou-Hanna, C.; Meurette, G.; Mohty, M.; Mosnier, J.-F.; Laboisse, C.L.; Bossard, C. Interferon-Alpha Promotes Th1 Response and Epithelial Apoptosis via Inflammasome Activation in Human Intestinal Mucosa. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Majtnerová, P.; Roušar, T. An overview of apoptosis assays detecting DNA fragmentation. Mol. Biol. Rep. 2018, 45, 1469–1478. [Google Scholar] [CrossRef] [Green Version]
- Do, B.H.; Nguyen, T.P.T.; Ho, N.Q.C.; Le, T.L.; Hoang, N.S.; Doan, C.C. Mitochondria-mediated Caspase-dependent and Caspase-independent apoptosis induced by aqueous extract from Moringa oleifera leaves in human melanoma cells. Mol. Biol. Rep. 2020, 47, 3675–3689. [Google Scholar] [CrossRef]
- Kang, M.H.; Reynolds, C.P. BcI-2 Inhibitors: Targeting mitochondrial apoptotic pathways in cancer therapy. Clin. Cancer Res. 2009, 15, 1126–1132. [Google Scholar] [CrossRef] [Green Version]
- Ghobrial, I.M.; Witzig, T.E.; Adjei, A.A. Targeting Apoptosis Pathways in Cancer Therapy. CA Cancer J. Clin. 2005, 55, 178–194. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta—Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Dai, D.F.; Rabinovitch, P.S.; Ungvari, Z. Mitochondria and cardiovascular aging. Circ. Res. 2012, 110, 1109–1124. [Google Scholar] [CrossRef]
- Yee, C.; Yang, W.; Hekimi, S. The intrinsic apoptosis pathway mediates the pro-longevity response to mitochondrial ROS in C elegans. Cell 2014, 157, 897–909. [Google Scholar] [CrossRef] [Green Version]
- Villalpando-Rodriguez, G.E.; Gibson, S.B. Reactive Oxygen Species (ROS) Regulates Different Types of Cell Death by Acting as a Rheostat. Oxid. Med. Cell. Longev. 2021, 2021, 9912436. [Google Scholar] [CrossRef] [PubMed]
- Madreiter-Sokolowski, C.T.; Thomas, C.; Ristow, M. Interrelation between ROS and Ca2+ in aging and age-related diseases. Redox Biol. 2020, 36, 101678. [Google Scholar] [CrossRef] [PubMed]
- Kajstura, M.; Halicka, D.; Pryjma, J.; Darzynkiewicz, Z. Discontinuous Fragmentation of Nuclear DNADuring Apoptosis Revealed by Discrete “Sub-G1” Peaks on DNA Content Histograms. Cytom. A 2007, 71, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.J.; Han, L.H.; Cong, R.S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Edlich, F. BCL-2 proteins and apoptosis: Recent insights and unknowns. Biochem. Biophys. Res. Commun. 2018, 500, 26–34. [Google Scholar] [CrossRef]
- Rex, J.; Lutz, A.; Faletti, L.E.; Albrecht, U.; Thomas, M.; Bode, J.G.; Borner, C.; Sawodny, O.; Merfort, I. IL-1β and TNFα differentially influence NF-κB activity and FasL-induced apoptosis in primary murine hepatocytes during LPS-induced inflammation. Front. Physiol. 2019, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Vandenabeele, P.; Galluzi, L.; Berghe, T.V.; Kroemer, G. Molecular mechanisms of necroptosis: An ordered cellular explosion. Nat. Rev. 2010, 11, 700–714. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fus-Kujawa, A.; Sieroń, Ł.; Dobrzyńska, E.; Chajec, Ł.; Mendrek, B.; Jarosz, N.; Głowacki, Ł.; Dubaj, K.; Dubaj, W.; Kowalczuk, A.; et al. Star Polymers as Non-Viral Carriers for Apoptosis Induction. Biomolecules 2022, 12, 608. https://doi.org/10.3390/biom12050608
Fus-Kujawa A, Sieroń Ł, Dobrzyńska E, Chajec Ł, Mendrek B, Jarosz N, Głowacki Ł, Dubaj K, Dubaj W, Kowalczuk A, et al. Star Polymers as Non-Viral Carriers for Apoptosis Induction. Biomolecules. 2022; 12(5):608. https://doi.org/10.3390/biom12050608
Chicago/Turabian StyleFus-Kujawa, Agnieszka, Łukasz Sieroń, Estera Dobrzyńska, Łukasz Chajec, Barbara Mendrek, Natalia Jarosz, Łukasz Głowacki, Kamila Dubaj, Wojciech Dubaj, Agnieszka Kowalczuk, and et al. 2022. "Star Polymers as Non-Viral Carriers for Apoptosis Induction" Biomolecules 12, no. 5: 608. https://doi.org/10.3390/biom12050608
APA StyleFus-Kujawa, A., Sieroń, Ł., Dobrzyńska, E., Chajec, Ł., Mendrek, B., Jarosz, N., Głowacki, Ł., Dubaj, K., Dubaj, W., Kowalczuk, A., & Bajdak-Rusinek, K. (2022). Star Polymers as Non-Viral Carriers for Apoptosis Induction. Biomolecules, 12(5), 608. https://doi.org/10.3390/biom12050608