
DNA-Binding Properties of a Novel Crenarchaeal Chromatin-Organizing Protein in Sulfolobus acidocaldarius


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analyses
2.2. Microbial Strains and Growth Conditions
2.3. Protein Overexpression and Purification
2.4. Western Blotting
2.5. Size Exclusion Chromatography
2.6. Electrophoretic Mobility Shift Assays
2.7. Chromatin Immunoprecipitation and High-Throughput Sequencing
2.8. Atomic Force Microscopy
3. Results and Discussion
3.1. Phylogenetic Occurrence of Sul12a
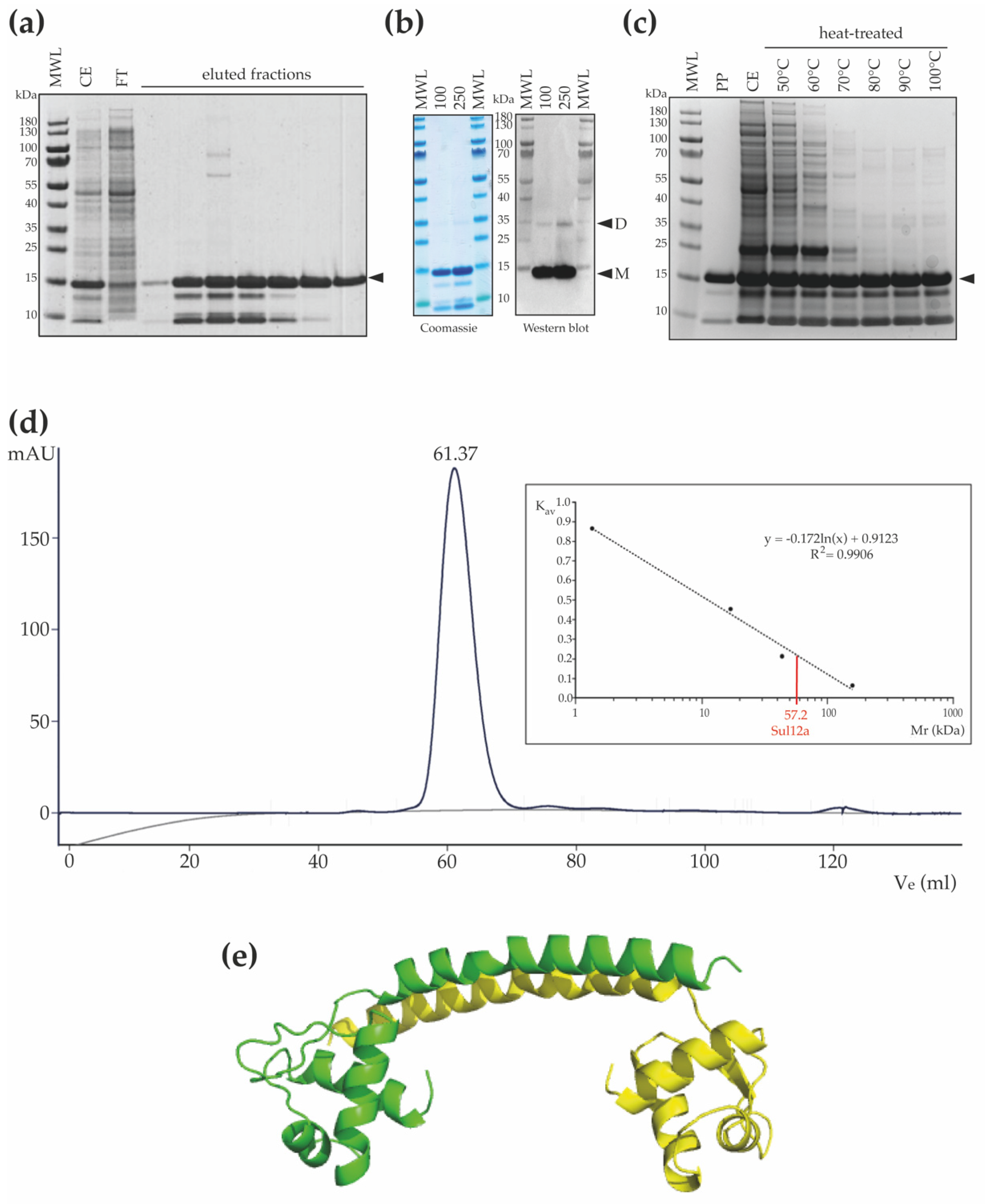
3.2. Structural Homology Model and Oligomeric State of Sul12a
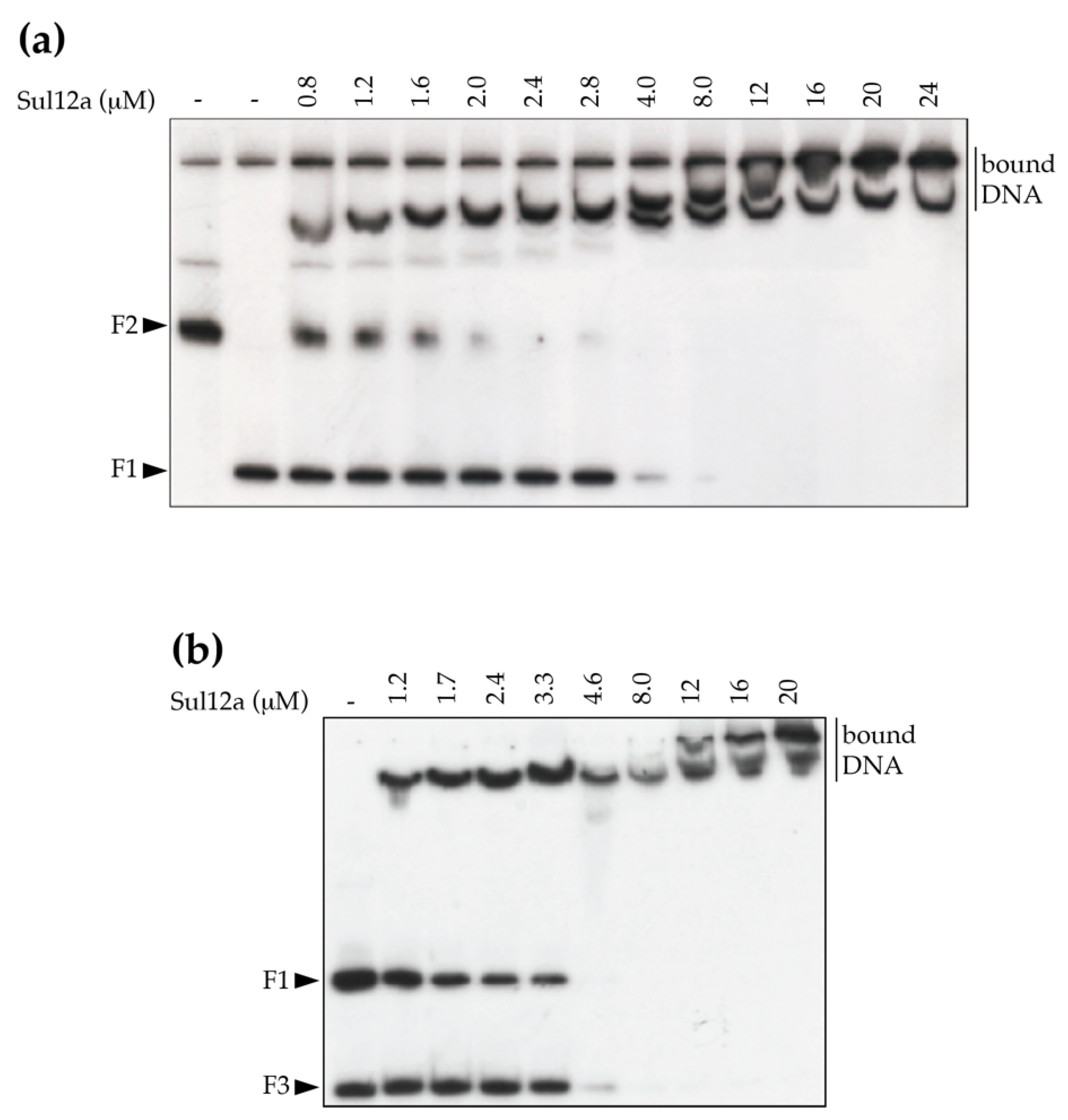
3.3. In Vitro DNA-Binding Characteristics of Sul12a
3.4. Genome-Wide Interactions between Sul12a and DNA
3.5. Sul12a Affects Architectural Characteristics of the DNA upon Binding
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luger, K.; Mäder, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Dame, R.T. The role of nucleoid-associated proteins in the organization and compaction of bacterial chromatin. Mol. Microbiol. 2005, 56, 858–870. [Google Scholar] [CrossRef] [PubMed]
- Henneman, B.; van Emmerik, C.; van Ingen, H.; Dame, R.T. Structure and function of archaeal histones. PLoS Genet. 2018, 14, e1007582. [Google Scholar] [CrossRef]
- Zhao, H.; Winogradoff, D.; Dalal, Y.; Papoian, G.A. The Oligomerization Landscape of Histones. Biophys. J. 2019, 116, 1845–1855. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.L.; Grayling, R.A.; Lurz, R.; Reeve, J.N. Archaeal nucleosomes. Proc. Natl. Acad. Sci. USA 1997, 94, 12633–12637. [Google Scholar] [CrossRef]
- Mattiroli, F.; Bhattacharyya, S.; Dyer, P.N.; White, A.E.; Sandman, K.; Burkhart, B.W.; Byrne, K.R.; Lee, T.; Ahn, N.G.; Santangelo, T.J.; et al. Structure of histone-based chromatin in Archaea. Science 2017, 357, 609–612. [Google Scholar] [CrossRef]
- Peeters, E.; Driessen, R.P.; Werner, F.; Dame, R.T. The interplay between nucleoid organization and transcription in archaeal genomes. Nat. Rev. Microbiol. 2015, 13, 333–341. [Google Scholar] [CrossRef]
- Driessen, R.P.; Dame, R.T. Structure and dynamics of the crenarchaeal nucleoid. Biochem. Soc. Trans. 2013, 41, 321–325. [Google Scholar] [CrossRef]
- Goyal, M.; Banerjee, C.; Nag, S.; Bandyopadhyay, U. The Alba protein family: Structure and function. Biochim. Biophys. Acta 2016, 1864, 570–583. [Google Scholar] [CrossRef]
- Driessen, R.P.; Meng, H.; Suresh, G.; Shahapure, R.; Lanzani, G.; Priyakumar, U.D.; White, M.F.; Schiessel, H.; van John, N.; Dame, R.H. Crenarchaeal chromatin proteins Cren7 and Sul7 compact DNA by inducing rigid bends. Nucleic Acids Res. 2012, 41, 196–205. [Google Scholar] [CrossRef]
- Driessen, R.P.; Lin, S.N.; Waterreus, W.-J.; van der Meulen, A.L.H.; van der Valk, R.A.; Laurens, N.; Moolenaar, G.F.; Pannu, N.S.; Wuite, G.J.L.; Goosen, N.; et al. Diverse architectural properties of Sso10a proteins: Evidence for a role in chromatin compaction and organization. Sci. Rep. 2016, 6, 29422. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Reeve, J.N. Transcription by an archaeal RNA polymerase is slowed but not blocked by an archaeal nucleosome. J. Bacteriol. 2004, 186, 3492–3498. [Google Scholar] [CrossRef] [PubMed]
- Choli, T.; Whitmann-Liebold, B.; Reinhardt, R. Microsequence analysis of DNA-binding proteins 7a, 7b, and 7e from the archaebacterium Sulfolobus acidocaldarius. J. Biol. Chem. 1988, 263, 7087–7093. [Google Scholar] [CrossRef]
- Guo, L.; Feng, Y.; Zhang, Z.; Yao, H.; Luo, Y.; Wang, J.; Huang, L. Biochemical and structural characterization of Cren7, a novel chromatin protein conserved among Crenarchaea. Nucleic Acids Res. 2008, 36, 1129–1137. [Google Scholar] [CrossRef]
- Chen, L.; Brügger, K.; Skovgaard, M.; Redder, P.; She, Q.; Torarinsson, E.; Greve, B.; Awayez, M.; Zibat, A.; Klenk, H.-P.; et al. The genome of Sulfolobus acidocaldarius, a model organism of the Crenarchaeota. J. Bacteriol. 2005, 187, 4992–4999. [Google Scholar] [CrossRef]
- Oberto, J. SyntTax: A web server linking synteny to prokaryotic taxonomy. BMC Bioinform. 2013, 14, 4. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Chan, P.P.; Holmes, A.D.; Smith, A.M.; Tran, D.; Lowe, T.M. The UCSC Archaeal Genome Browser: 2012 update. Nucleic Acids Res. 2011, 40, D646–D652. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-Model: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; van Wolferen, M.; Wagner, A.; Lassak, K.; Meyer, B.H.; Reimann, J.; Albers, S.-V. Versatile genetic toolbox for the crenarchaeote Sulfolobus acidocaldarius. Front. Microbiol. 2012, 3, 214. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.D.; Brock, K.M.; Belly, R.T.; Weiss, R.L. Sulfolobus: A new genus of sulfur-oxidizing bacteria living at low pH and high temperature. Arch. Microbiol. 1972, 84, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, M.D.; Magelhaes, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Enoru-Eta, J.; Gigot, D.; Thia-Toong, T.L.; Glansdorff, N.; Charlier, D. Purification and characterization of Sa-Lrp, a DNA-binding protein from the extreme thermoacidophilic archaeon Sulfolobus acidocaldarius homologous to the bacterial regulator Lrp. J. Bacteriol. 2000, 182, 3661–3672. [Google Scholar] [CrossRef]
- Peeters, E.; Wartel, C.; Maes, D.; Charlier, D. Analysis of the DNA-binding sequence specificity of the archaeal transcriptional regulator Ss-LrpB from Sulfolobus solfataricus by systematic mutagenesis and high resolution contact probing. Nucleic Acids Res. 2007, 35, 623–633. [Google Scholar] [CrossRef]
- Wang, K.; Lindas, A.-C. Chromatin Immunoprecipitation Assay in the Hyperthermoacidophilic Crenarchaeon, Sulfolobus acidocaldarius. Methods Mol. Biol. 2018, 1689, 139–146. [Google Scholar]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, T.; Meyer, C.; Eeckhoute, J.; Johnson, D.; Bernstein, D.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-seq (MACS). Genome Biol. 2015, 9, R137. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdottir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]
- Chen, L.; Chen, L.-R.; Zhou, X.E.; Wang, Y.; Kahsai, M.A.; Clark, A.T.; Edmondson, S.P.; Liu, Z.-J.; Rose, J.P.; Wang, B.-C.; et al. The hyperthermophilic protein Sso10a is a dimer o.of winged helic DNA-binding domains linked by an antiparallel coiled coil rod. J. Mol. Biol. 2004, 341, 73–91. [Google Scholar] [CrossRef]
- Ahmad, M.U.D.; Waege, I.; Hausner, W.; Thomm, M.; Boos, W.; Diederichs, K.; Welte, W. Structural insights into nonspecific binding of DNA by TrmBL2, an archaeal chromatin protein. J. Mol. Biol. 2015, 427, 3216–3229. [Google Scholar] [CrossRef]
- Vogt, M.S.; Völpel, S.L.; Albers, S.-V.; Essen, L.O.; Banerjee, A. Crystal structure of an Lrs14-like archaeal biofilm regulator from Sulfolobus acidocaldarius. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Orell, A.; Peeters, E.; Vassen, V.; Jachlewski, S.; Schalles, S.; Siebers, B.; Albers, S.-V. Lrs14 transcriptional regulatorsinfluence biofilm formation and cell motility of Crenarchaea. ISME J. 2013, 7, 1886–1898. [Google Scholar] [CrossRef] [PubMed]
- Driessen, R.P.; Dame, R.T. Nucleoid-associated proteins in Crenarchaeota. Biochem. Soc. Trans. 2011, 39, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Dame, R.T.; Wyman, C.; Goosen, N. H-NS mediated compaction of DNA visualized by atomic force microscopy. Nucleic Acids Res. 2000, 28, 3504–3510. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nr | Genomic Coordinates of ChIP-Seq Enrichment Region | Fold Enrichment 1 | Gene Number of Nearest ORF | Peak Summit Location 2 |
|---|---|---|---|---|
| 1 | 11,506–12,946 | 2.5 | Saci_0017 | G |
| 2 | 395,203–396,359 | 4.1 | Saci_0472 | G |
| 3 | 792,502–793,792 | 3.4 | Saci_0991 | G |
| 4 | 840,354–841,874 | 5.0 | Saci_1041 | G |
| 5 | 844,058–848,204 | 2.7 | Saci_1045 | G |
| 6 | 1,053,261–1,054,510 | 4.6 | Saci_1237 | I |
| 7 | 1,142,415–1,144,126 | 3.2 | Saci_1339 | I |
| 8 | 1,153,758–1,156,200 | 3.8 | Saci_1353 | G |
| 9 | 1,174,705–1,176,888 | 5.1 | Saci_1374 | G |
| 10 | 1,283,681–1,285,346 | 2.6 | Saci_1506 | G |
| 11 | 1,659,329–1,663,786 | 2.9 | Saci_1872 | G |
| 12 | 1,704,818–1,705,948 | 2.7 | Saci_1906 | G |
| 13 | 1,758,951–1,762,070 | 2.8 | Saci_1947 | G |
| 14 | 1,916,073–1,927,216 | 6.9 | Saci_2102 | G |
| 15 | 2,086,681–2,091,395 | 2.7 | Saci_2246 | G |
| 16 | 2,189,724–2,193,028 | 2.5 | Saci_2344 | G |
| Probe | Experiment 1 | Length (bp) | KDapp (μM) |
|---|---|---|---|
| Enrichment probe 9 (F1) | a | 337 | 3.9 |
| carAB promoter region (F2) | a | 578 | 1.2 |
| Enrichment probe 9 (F1) | b | 337 | 2.3 |
| argO promoter region (F3) | b | 145 | 3.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemmens, L.; Wang, K.; Ruykens, E.; Nguyen, V.T.; Lindås, A.-C.; Willaert, R.; Couturier, M.; Peeters, E. DNA-Binding Properties of a Novel Crenarchaeal Chromatin-Organizing Protein in Sulfolobus acidocaldarius. Biomolecules 2022, 12, 524. https://doi.org/10.3390/biom12040524
Lemmens L, Wang K, Ruykens E, Nguyen VT, Lindås A-C, Willaert R, Couturier M, Peeters E. DNA-Binding Properties of a Novel Crenarchaeal Chromatin-Organizing Protein in Sulfolobus acidocaldarius. Biomolecules. 2022; 12(4):524. https://doi.org/10.3390/biom12040524
Chicago/Turabian StyleLemmens, Liesbeth, Kun Wang, Ebert Ruykens, Van Tinh Nguyen, Ann-Christin Lindås, Ronnie Willaert, Mohea Couturier, and Eveline Peeters. 2022. "DNA-Binding Properties of a Novel Crenarchaeal Chromatin-Organizing Protein in Sulfolobus acidocaldarius" Biomolecules 12, no. 4: 524. https://doi.org/10.3390/biom12040524
APA StyleLemmens, L., Wang, K., Ruykens, E., Nguyen, V. T., Lindås, A.-C., Willaert, R., Couturier, M., & Peeters, E. (2022). DNA-Binding Properties of a Novel Crenarchaeal Chromatin-Organizing Protein in Sulfolobus acidocaldarius. Biomolecules, 12(4), 524. https://doi.org/10.3390/biom12040524