
Targeted Degradation of 53BP1 Using Ubiquitin Variant Induced Proximity



Abstract
:1. Introduction
2. Materials and Methods
2.1. Phage Display and Ubiquitin Variant (UbV) Selections
2.2. ELISA Binding Assays
2.3. UbV Expression and Purification
2.4. In Vitro Ubiquitination Assays
2.5. Cell Lines and Culture Conditions
2.6. Plasmids and Transfection
2.7. Protein Detection Using Western Blot and Chemiluminescence
2.8. Detection of Intracellular Protein Interaction Using Co-Immunoprecipitation
2.9. Epi-Fluorescent Microscopy
2.10. MTT Survival Assay
2.11. Statistical Analysis
3. Results
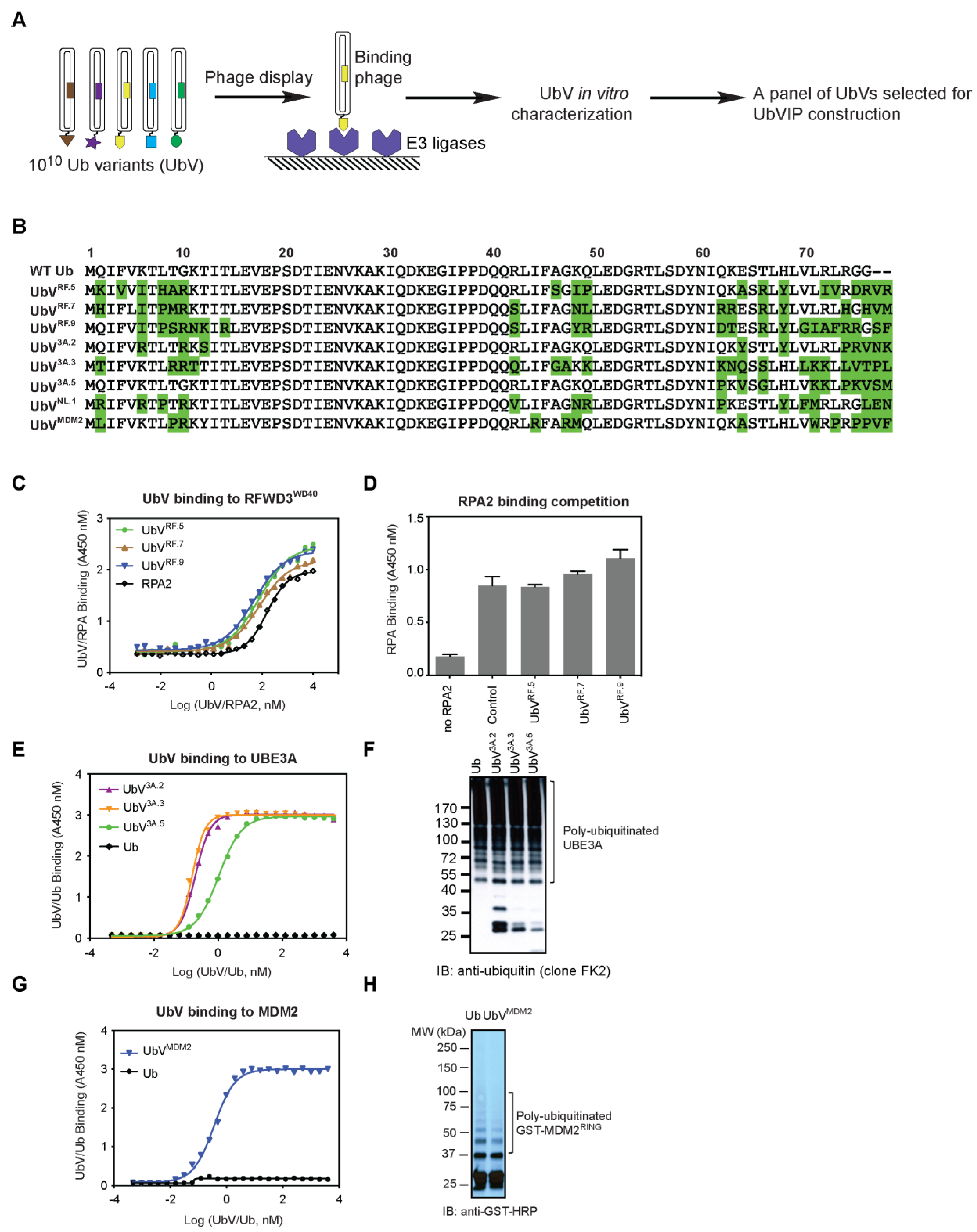
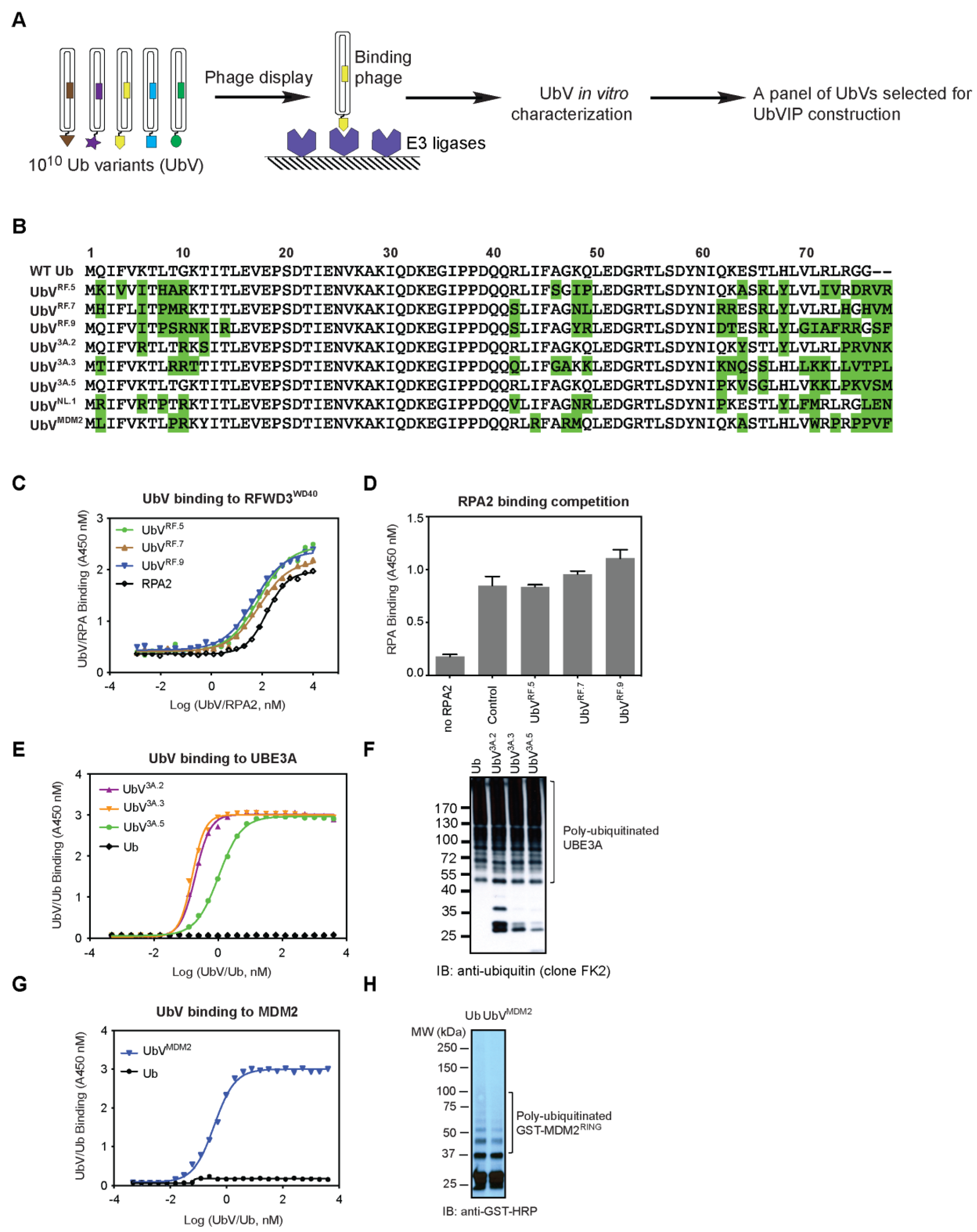
3.1. Identification of Non-Inhibitory UbV Binders for Selected E3 Ligases
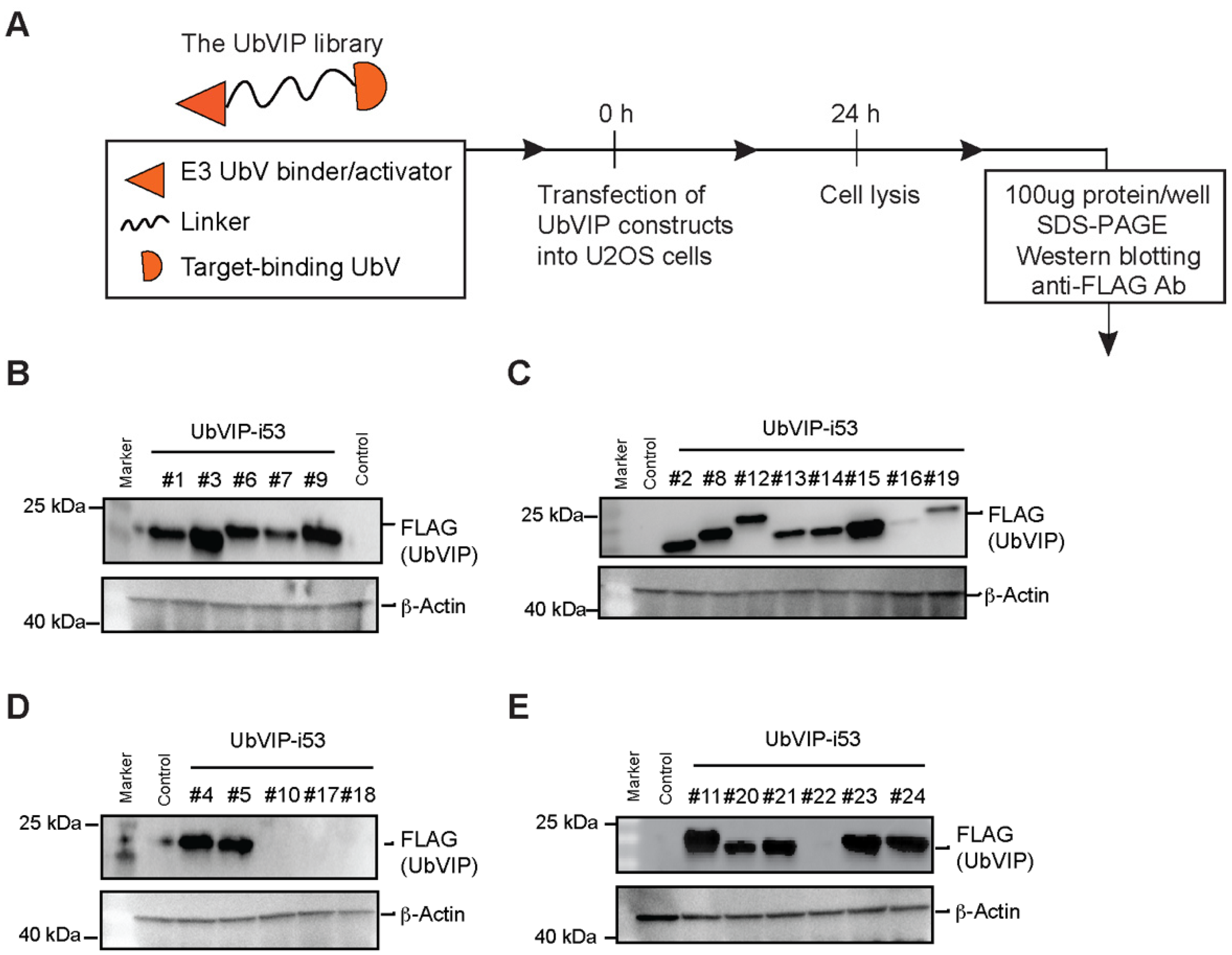
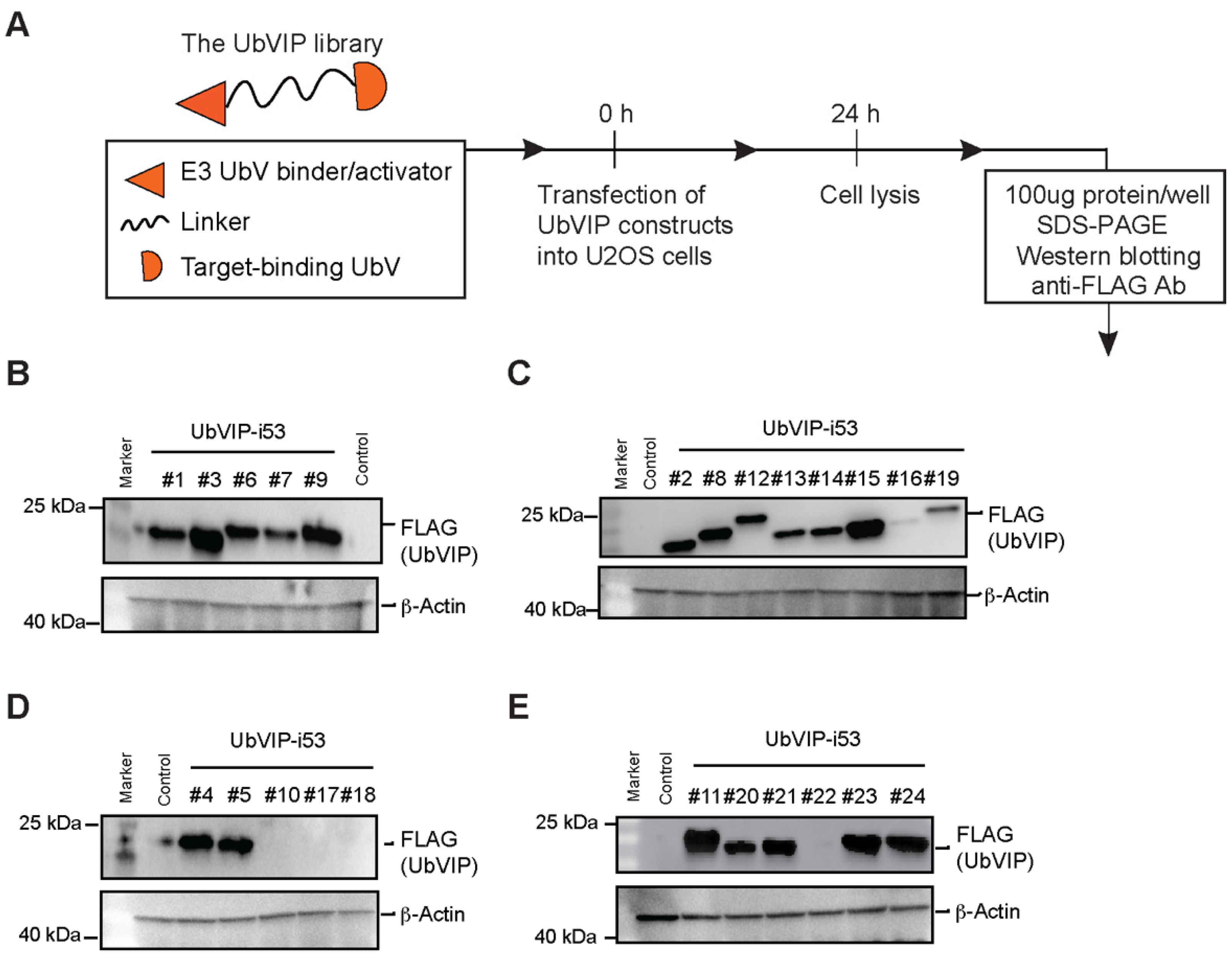
3.2. Design and Cellular Expression Analysis of a UbVIP Library
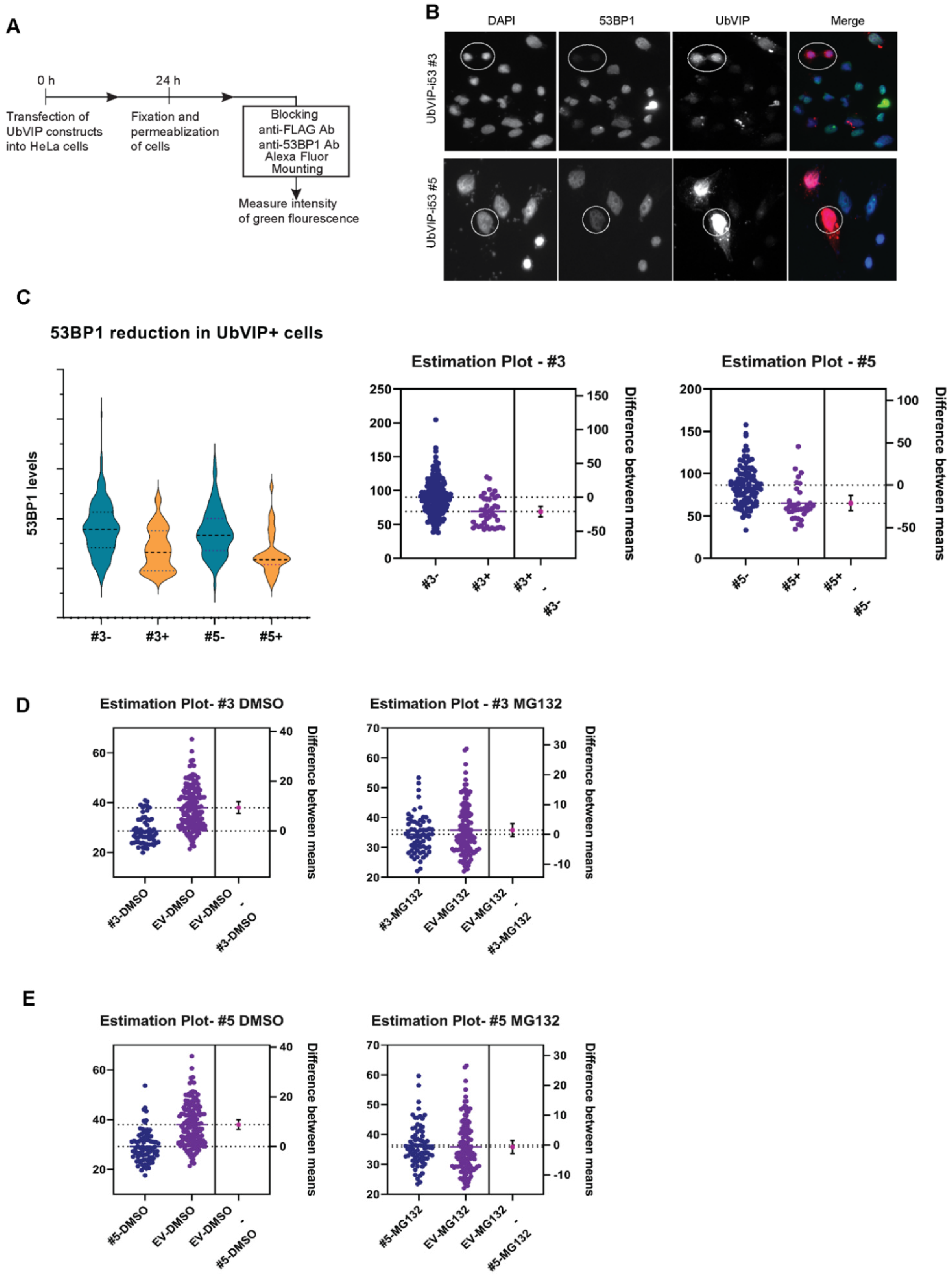
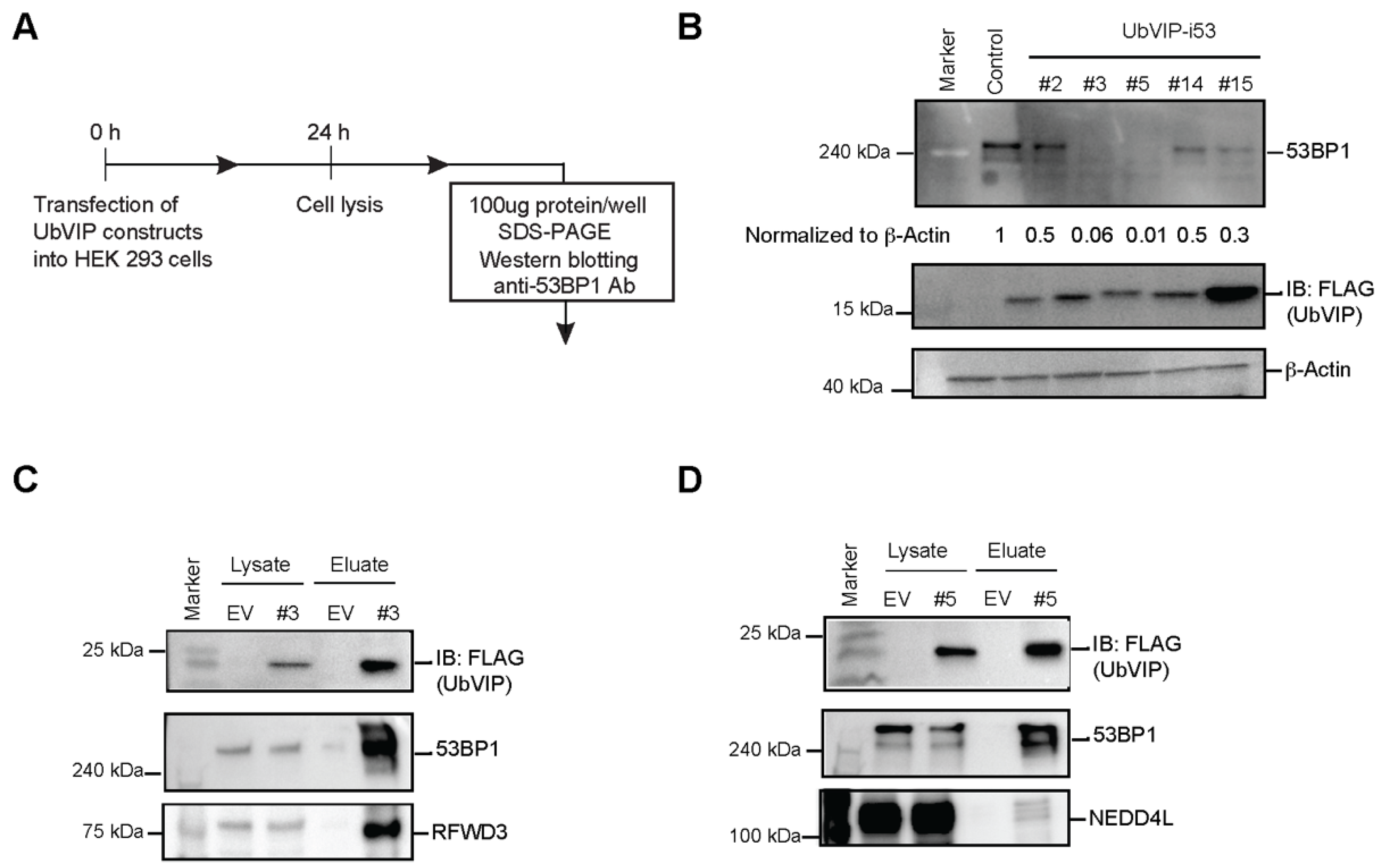
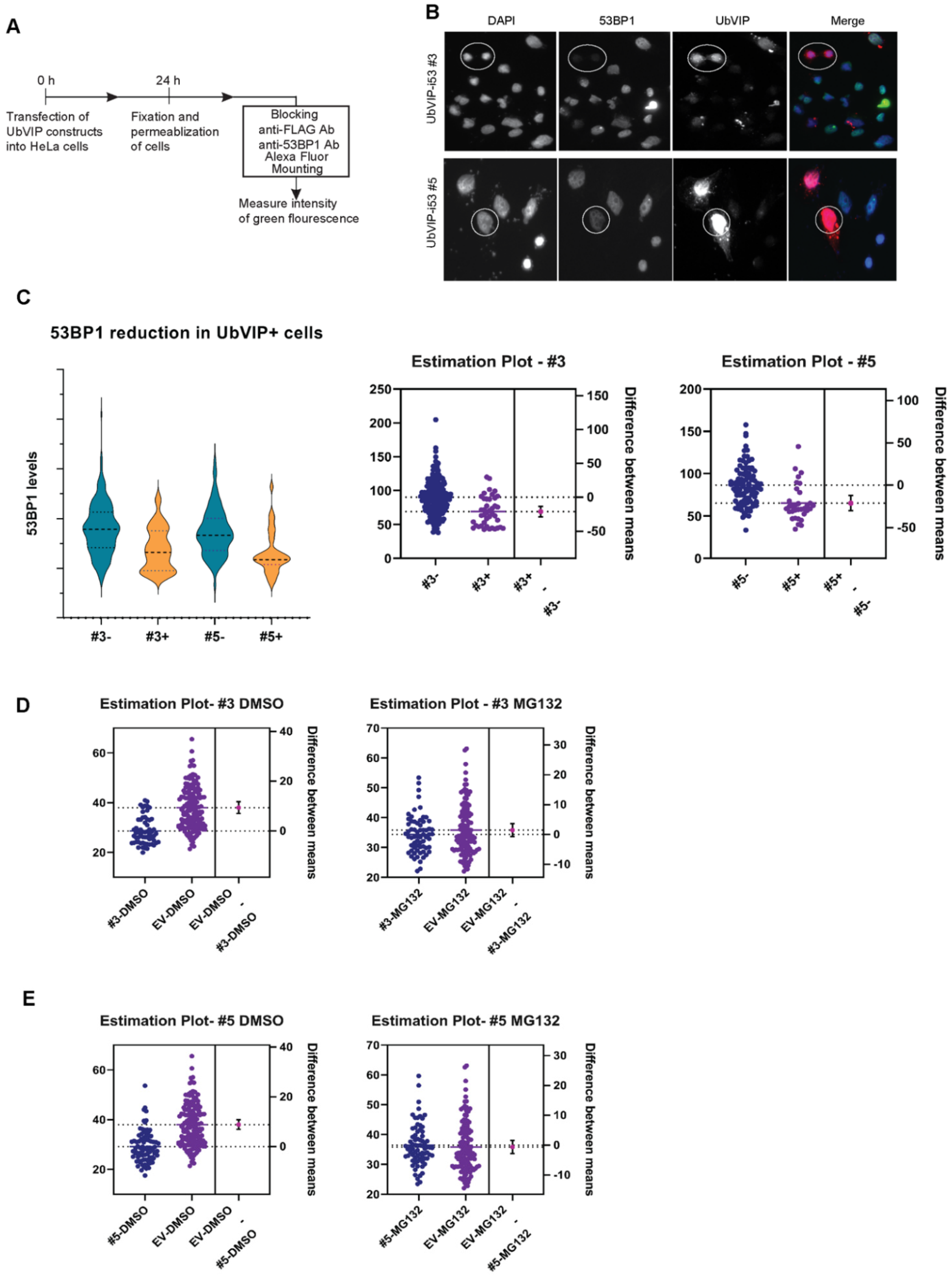
3.3. Reduction of 53BP1 Protein Level in the Presence of Selected UbVIPs
3.4. UbVIP-i53#3 and #5 Target 53BP1 for Proteasome-Mediated Degradation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, P.; Duong, D.; Seyfried, N.T.; Cheng, D.; Xie, Y.; Robert, J.; Rush, J.; Hochstrasser, M.; Finley, D.; Peng, J. Quantitative proteomics reveals the function of unconventional ubiquitin chains in proteasomal degradation. Cell 2009, 137, 133–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziv, I.; Matiuhin, Y.; Kirkpatrick, D.; Erpapazoglou, Z.; Leon, S.; Pantazopoulou, M.; Kim, W.; Gygi, S.P.; Haguenauer-Tsapis, R.; Reis, N.; et al. A perturbed ubiquitin landscape distinguishes between ubiquitin in trafficking and in proteolysis. Mol. Cell Proteom. 2011, 10, M111.009753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershko, A.; Heller, H.; Elias, S.; Ciechanover, A. Components of ubiquitin-protein ligase system. Resolution, affinity purification, and role in protein breakdown. J. Biol. Chem. 1983, 258, 8206–8214. [Google Scholar] [CrossRef]
- Zhang, W.; Sidhu, S.S. Development of inhibitors in the ubiquitination cascade. FEBS Lett. 2014, 588, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Zheng, N.; Shabek, N. Ubiquitin Ligases: Structure, Function, and Regulation. Annu. Rev. Biochem. 2017, 86, 129–157. [Google Scholar] [CrossRef]
- Ottis, P.; Crews, C.M. Proteolysis-Targeting Chimeras: Induced Protein Degradation as a Therapeutic Strategy. ACS Chem. Biol. 2017, 12, 892–898. [Google Scholar] [CrossRef]
- Sakamoto, K.M.; Kim, K.B.; Kumagai, A.; Mercurio, F.; Crews, C.M.; Deshaies, R.J. Protacs: Chimeric molecules that target proteins to the Skp1–Cullin–F box complex for ubiquitination and degradation. Proc. Natl. Acad. Sci. USA 2001, 98, 8554–8559. [Google Scholar] [CrossRef] [Green Version]
- Bondeson, D.P.; Mares, A.; Smith, I.E.D.; Ko, E.; Campos, S.; Miah, A.H.; Mulholland, K.E.; Routly, N.; Buckley, D.L.; Gustafson, J.L.; et al. Catalytic in vivo protein knockdown by small-molecule PROTACs. Nat. Chem. Biol. 2015, 11, 611–617. [Google Scholar] [CrossRef] [Green Version]
- Neklesa, T.K.; Jin, M.; Crew, A.P.; Rossi, A.K.; Willard, R.R.; Dong, H.; Siu, K.; Wang, J.; Gordon, A.D.; Chen, X.; et al. ARV-330: Androgen receptor PROTAC degrader for prostate cancer. J. Clin. Oncol. 2016, 34, 267. [Google Scholar] [CrossRef]
- Salami, J.; Crews, C.M. Waste disposal—An attractive strategy for cancer therapy. Science 2017, 355, 1163–1167. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Gao, H.; Yang, Y.; He, M.; Wu, Y.; Song, Y.; Tong, Y.; Rao, Y. PROTACs: Great opportunities for academia and industry. Signal Transduct. Target. Ther. 2019, 4, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheepstra, M.; Hekking, K.F.W.; van Hijfte, L.; Folmer, R.H.A. Bivalent Ligands for Protein Degradation in Drug Discovery. Comput. Struct. Biotechnol. J. 2019, 17, 160–176. [Google Scholar] [CrossRef]
- Konstantinidou, M.; Li, J.; Zhang, B.; Wang, Z.; Shaabani, S.; Ter Brake, F.; Essa, K.; Dömling, A. PROTACs—A game-changing technology. Expert Opin. Drug Discov. 2019, 14, 1255–1268. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, N.; Mallette, E.; Zhang, W. Targeted modulation of E3 ligases using engineered ubiquitin variants. FEBS J. 2021, 288, 2143–2165. [Google Scholar] [CrossRef]
- Ernst, A.; Avvakumov, G.; Tong, J.; Fan, Y.; Zhao, Y.; Alberts, P.; Persaud, A.; Walker, J.R.; Neculai, A.-M.; Neculai, D.; et al. A strategy for modulation of enzymes in the ubiquitin system. Science 2013, 339, 590–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Wu, K.-P.; Sartori, M.A.; Kamadurai, H.B.; Ordureau, A.; Jiang, C.; Mercredi, P.Y.; Murchie, R.; Hu, J.; Persaud, A.; et al. System-Wide Modulation of HECT E3 Ligases with Selective Ubiquitin Variant Probes. Mol. Cell 2016, 62, 121–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrielsen, M.; Buetow, L.; Nakasone, M.A.; Ahmed, S.F.; Sibbet, G.J.; Smith, B.O.; Zhang, W.; Sidhu, S.S.; Huang, D.T. A General Strategy for Discovery of Inhibitors and Activators of RING and U-box E3 Ligases with Ubiquitin Variants. Mol. Cell 2017, 68, 456–470.e10. [Google Scholar] [CrossRef] [Green Version]
- Watson, E.R.; Grace, C.R.R.; Zhang, W.; Miller, D.J.; Davidson, I.F.; Prabu, J.R.; Yu, S.; Bolhuis, D.L.; Kulko, E.T.; Vollrath, R.; et al. Protein engineering of a ubiquitin-variant inhibitor of APC/C identifies a cryptic K48 ubiquitin chain binding site. Proc. Natl. Acad. Sci. USA 2019, 116, 17280–17289. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Chu, J.; Yucer, N.; Leng, M.; Wang, S.-Y.; Chen, B.P.C.; Hittelman, W.N.; Wang, Y. RING finger and WD repeat domain 3 (RFWD3) associates with replication protein A (RPA) and facilitates RPA-mediated DNA damage response. J. Biol. Chem. 2011, 286, 22314–22322. [Google Scholar] [CrossRef] [Green Version]
- Elia, A.E.; Wang, D.C.; Willis, N.A.; Boardman, A.P.; Hajdu, I.; Adeyemi, R.O.; Lowry, E.; Gygi, S.P.; Scully, R.; Elledge, S.J. RFWD3-Dependent Ubiquitination of RPA Regulates Repair at Stalled Replication Forks. Mol. Cell 2015, 60, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Pashkova, N.; Gakhar, L.; Winistorfer, S.C.; Yu, L.; Ramaswamy, S.; Piper, R.C. WD40 repeat propellers define a ubiquitin-binding domain that regulates turnover of F box proteins. Mol. Cell 2010, 40, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, J.; Paranjape, S.R.; Walker, M.P.; Choudhury, R.; Wolter, J.; Fragola, G.; Emanuele, M.J.; Major, M.; Zylka, M.J. The autism-linked UBE3A T485A mutant E3 ubiquitin ligase activates the Wnt / Beta-catenin pathway by inhibiting the proteasome. J. Biol. Chem. 2017, 292, 12503–12515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneekloth, A.R.; Pucheault, M.; Tae, H.S.; Crews, C.M. Targeted intracellular protein degradation induced by a small molecule: En route to chemical proteomics. Bioorg. Med. Chem. Lett. 2008, 18, 5904–5908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzian, T.; Wang, Y.; Van Pelt, C.S.; Box, N.F.; Travis, E.L.; Lozano, G. Guillermina. Haploinsufficiency of Mdm2 and Mdm4 in Tumorigenesis and Development. Mol. Cell. Biol. 2007, 27, 5479–5485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, K.; Klejnot, M.; Kowalczyk, D.; Hock, A.K.; Sibbet, G.J.; Vousden, K.H.; Huang, D.T. Structural analysis of MDM2 RING separates degradation from regulation of p53 transcription activity. Nat. Struct. Mol. Biol. 2017, 24, 578–587. [Google Scholar] [CrossRef]
- Cyrus, K.; Wehenkel, M.; Choi, E.-Y.; Lee, H.; Swanson, H.; Kim, K.-B. Jostling for position: Optimizing linker location in the design of estrogen receptor-targeting PROTACs. ChemMedChem 2010, 5, 979–985. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zaro, J.L.; Shen, W.-C. Fusion protein linkers: Property, design and functionality. Adv. Drug Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef] [Green Version]
- George, R.A.; Heringa, J. An analysis of protein domain linkers: Their classification and role in protein folding. Protein Eng. 2002, 15, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Canny, M.D.; Moatti, N.; Wan, L.C.K.; Fradet-Turcotte, A.; Krasner, D.; Mateos-Gomez, A.P.; Zimmermann, M.; Orthwein, A.; Juang, Y.-C.; Zhang, W.; et al. Inhibition of 53BP1 favors homology-dependent DNA repair and increases CRISPR-Cas9 genome-editing efficiency. Nat. Biotechnol. 2018, 36, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Ward, I.M.; Minn, K.; Van Deursen, J.; Chen, J. p53 Binding Protein 53BP1 Is Required for DNA Damage Responses and Tumor Suppression in Mice. Mol. Cell. Biol. 2003, 23, 2556–2563. [Google Scholar] [CrossRef] [Green Version]
- Gorelik, M.; Orlicky, S.; Sartori, M.A.; Tang, X.; Marcon, E.; Kurinov, I.; Greenblatt, J.F.; Tyers, M.; Moffat, J.; Sicheri, F.; et al. Inhibition of SCF ubiquitin ligases by engineered ubiquitin variants that target the Cul1 binding site on the Skp1-F-box interface. Proc. Natl. Acad. Sci. USA. 2016, 113, 3527–3532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorelik, M.; Manczyk, N.; Pavlenco, A.; Kurinov, I.; Sidhu, S.S.; Sicheri, F. A Structure-Based Strategy for Engineering Selective Ubiquitin Variant Inhibitors of Skp1-Cul1-F-Box Ubiquitin Ligases. Structure 2018, 26, 1226–1236.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorba, A.; Nguyen, C.; Xu, Y.; Starr, J.; Borzilleri, K.; Smith, J.; Zhu, H.; Farley, K.A.; Ding, W.; Schiemer, J.; et al. Delineating the role of cooperativity in the design of potent PROTACs for BTK. Proc. Natl. Acad. Sci. USA 2018, 115, E7285–E7292. [Google Scholar] [CrossRef] [Green Version]
- Alabi, S.B.; Crews, C.M. Major advances in targeted protein degradation: PROTACs, LYTACs, and MADTACs. J. Biol. Chem. 2021, 296, 100647. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.E.; Wang, S.L.; Jaime-Figueroa, S.; Harbin, A.; Wang, J.; Hamman, B.D.; Crews, C.M. Differential PROTAC substrate specificity dictated by orientation of recruited E3 ligase. Nat. Commun. 2019, 10, 131. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identifier | Construct |
|---|---|
| UbVIP-i53#1 | UbVRF.5-8aa-i53 |
| UbVIP-i53#2 | UbVRF.7-8aa-i53 |
| UbVIP-i53#3 | UbVRF.9-8aa-i53 |
| UbVIP-i53#4 | UbV3A.2-8aa-i53 |
| UbVIP-i53#5 | UbVNL.1-8aa-i53 |
| UbVIP-i53#6 | UbVMDM2-8aa-i53 |
| UbVIP-i53#7 | UbVRF.5-15aa-i53 |
| UbVIP-i53#8 | UbVRF.7-15aa-i53 |
| UbVIP-i53#9 | UbVRF.9-15aa-i53 |
| UbVIP-i53#10 | UbV3A.2-15aa-i53 |
| UbVIP-i53#11 | UbVNL.1-15aa-i53 |
| UbVIP-i53#12 | UbVMDM2-15aa-i53 |
| UbVIP-i53#13 | i53-8aa-UbVRF.5 |
| UbVIP-i53#14 | i53-8aa-UbVRF.7 |
| UbVIP-i53#15 | i53-8aa-UbVRF.9 |
| UbVIP-i53#16 | i53-8aa-UbV3A.2 |
| UbVIP-i53#17 | i53-8aa-UbVNL.1 |
| UbVIP-i53#18 | i53-8aa-UbVMDM2 |
| UbVIP-i53#19 | i53-15aa-UbVRF.5 |
| UbVIP-i53#20 | i53-15aa-UbVRF.7 |
| UbVIP-i53#21 | i53-15aa-UbVRF.9 |
| UbVIP-i53#22 | i53-15aa-UbV3A.2 |
| UbVIP-i53#23 | i53-15aa-UbVNL.1 |
| UbVIP-i53#24 | i53-15aa-UbVMDM2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aminu, B.; Fux, J.; Mallette, E.; Petersen, N.; Zhang, W. Targeted Degradation of 53BP1 Using Ubiquitin Variant Induced Proximity. Biomolecules 2022, 12, 479. https://doi.org/10.3390/biom12040479
Aminu B, Fux J, Mallette E, Petersen N, Zhang W. Targeted Degradation of 53BP1 Using Ubiquitin Variant Induced Proximity. Biomolecules. 2022; 12(4):479. https://doi.org/10.3390/biom12040479
Chicago/Turabian StyleAminu, Bayonle, Julia Fux, Evan Mallette, Nathaniel Petersen, and Wei Zhang. 2022. "Targeted Degradation of 53BP1 Using Ubiquitin Variant Induced Proximity" Biomolecules 12, no. 4: 479. https://doi.org/10.3390/biom12040479
APA StyleAminu, B., Fux, J., Mallette, E., Petersen, N., & Zhang, W. (2022). Targeted Degradation of 53BP1 Using Ubiquitin Variant Induced Proximity. Biomolecules, 12(4), 479. https://doi.org/10.3390/biom12040479