
Inuloxin A Inhibits Seedling Growth and Affects Redox System of Lycopersicon esculentum Mill. and Lepidium sativum L.

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Chemicals
2.2. Effect of InA on Rootlets and Biomass
2.3. Determination of Ascorbate and Glutathione Pools
2.4. Determination of Total Proteins and Protein Thiol Labelling
2.5. Statistical Analyses
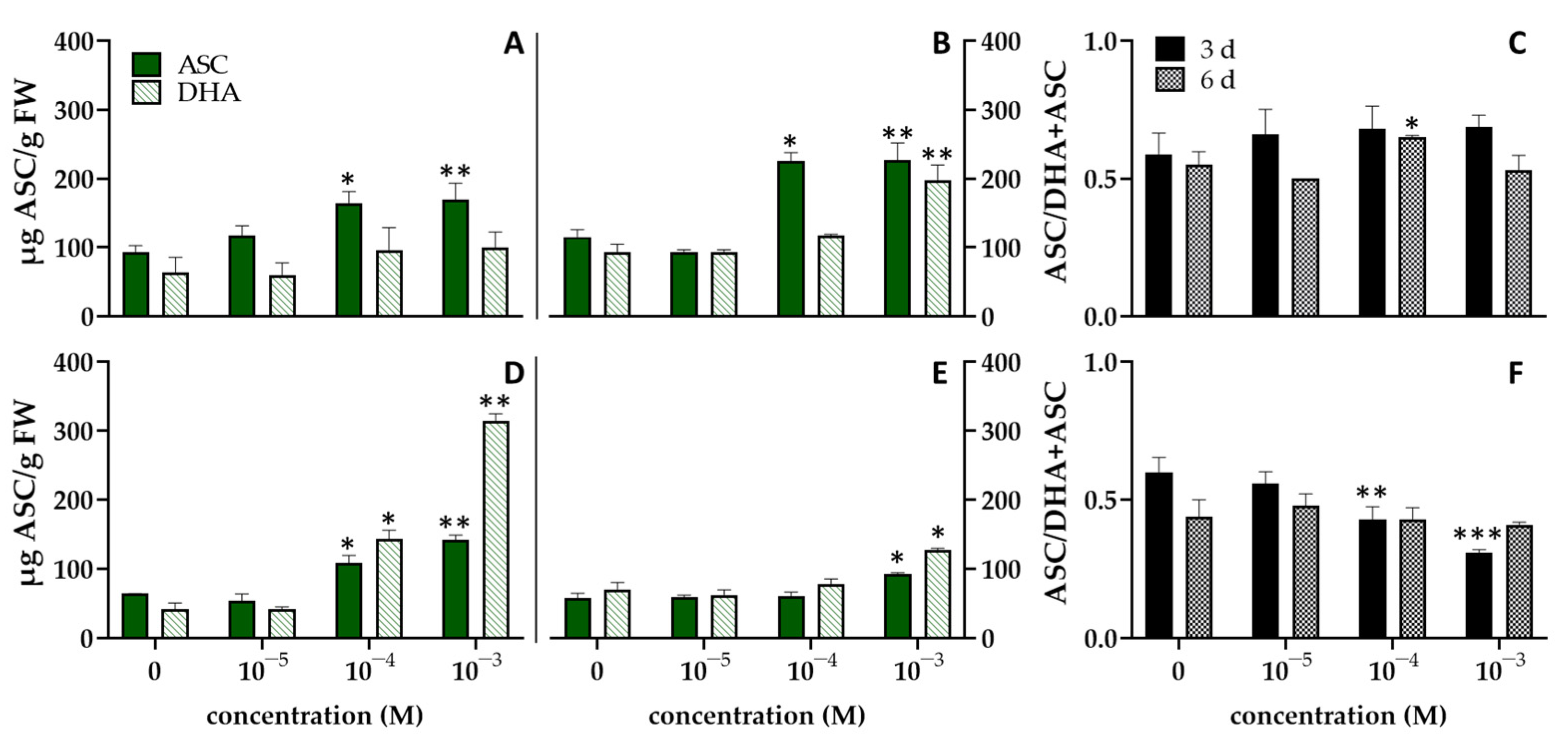
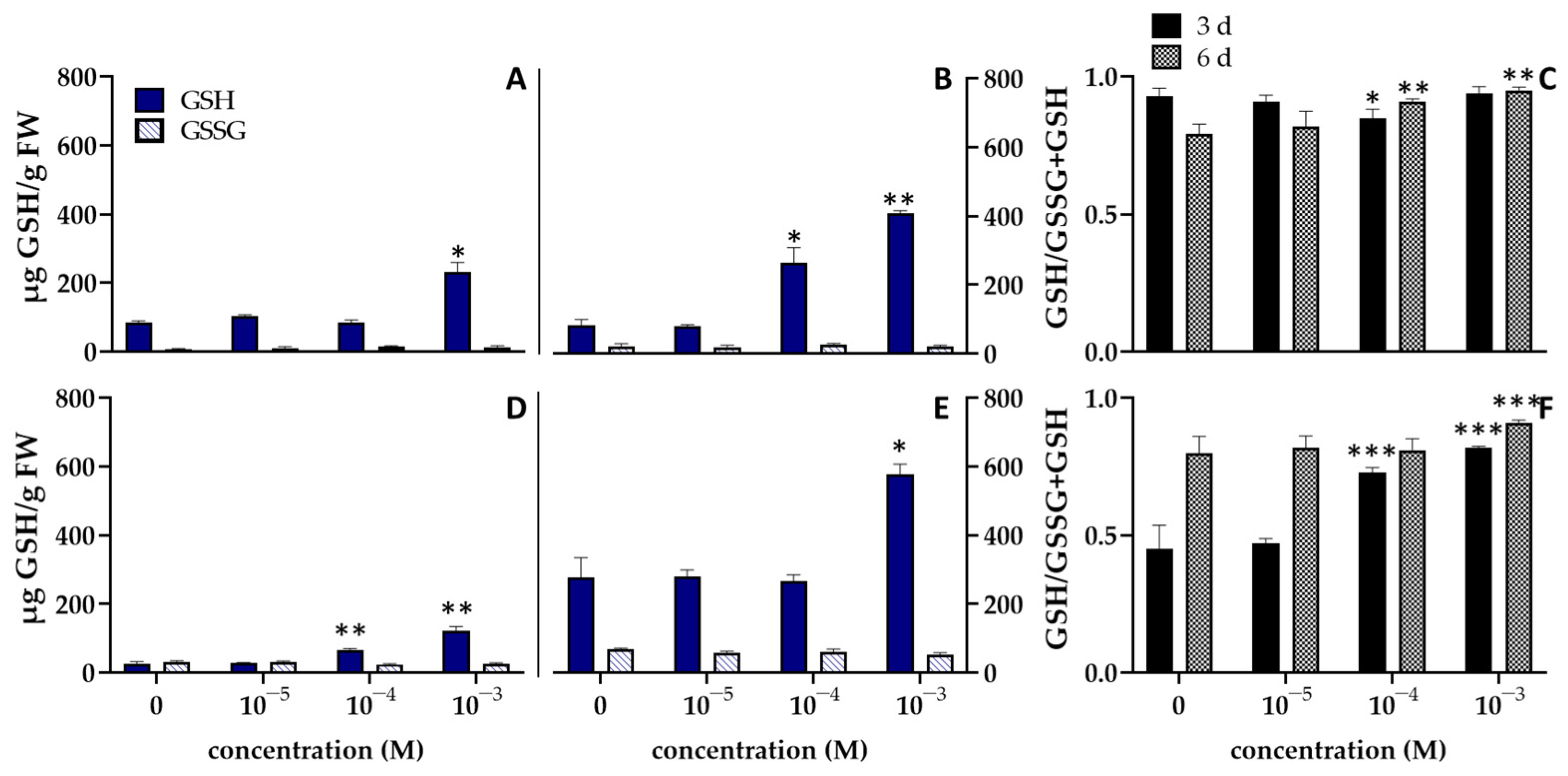
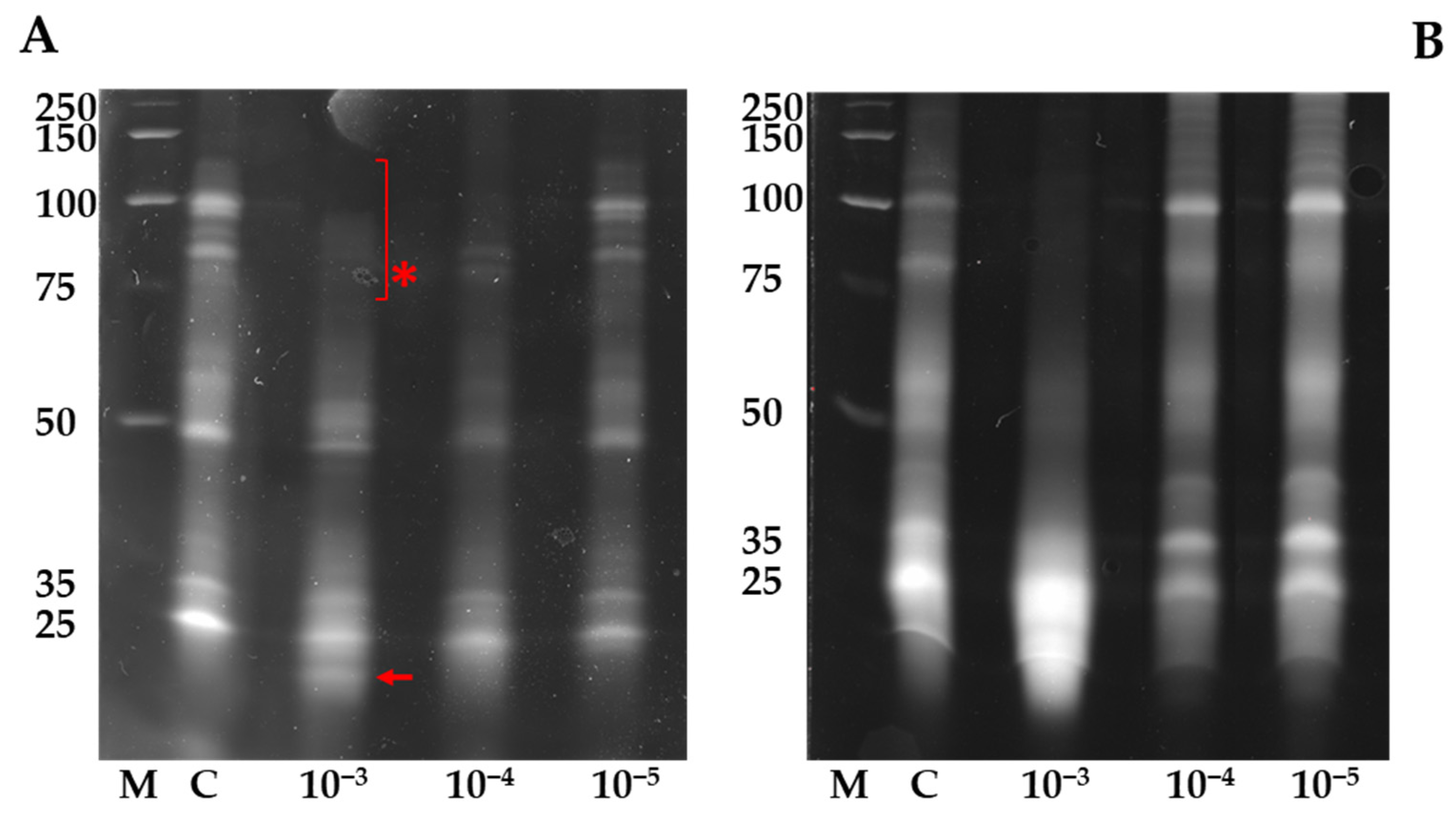
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sterck, L.; Rombauts, S.; Vandepoele, K.; Rouze, P.; Van de Peer, Y. How many genes are there in plants (… and why are they there)? Curr. Plant Biol. 2007, 10, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Ninkovic, V.; Rensing, M.; Dahlin, I.; Markovic, D. Who is my neighbor? Volatile cues in plant interactions. Plant Signal. Behav. 2019, 14, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Inderjit, M.; Duke, S.O. Ecophysiological aspects of allelopathy. Planta 2003, 217, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.S.; Mallik, A.U.; Luo, S.M. Allelopathy in Sustainable Agriculture and Forestry; Springer Science+Business Media, LLC: New York, NY, USA, 2008. [Google Scholar]
- Macías, F.A.; Mejías, F.J.; Molinillo, J.M. Recent advances in allelopathy for weed control: From knowledge to applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef]
- Kong, C.H.; Xuan, T.D.; Khanh, T.D.; Tran, H.D.; Trung, N.T. Allelochemicals and signaling chemicals in plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef] [Green Version]
- Vyvyan, J.R. Allelochemicals as leads for new herbicides and agrochemicals. Tetrahedron 2002, 58, 1631–1646. [Google Scholar] [CrossRef]
- Palanivel, H.; Tilaye, G.; Belliathan, S.K.; Benor, S.; Abera, S.; Kamaraj, M. Allelochemicals as Natural Herbicides for Sustainable Agriculture to Promote a Cleaner Environment. In Strategies and Tools for Pollutant Mitigation: Avenues to a Cleaner Environment; Aravind, J., Kamaraj, M., Prashanthi Devi, M., Rajakumar, S., Eds.; Springer Nature: Cham, Switzerland, 2021; pp. 93–116. [Google Scholar]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef]
- Marrone, P.G. Pesticidal natural products–status and future potential. Pest Manag. Sci. 2019, 75, 2325–2340. [Google Scholar] [CrossRef]
- Freda, F.; Masi, M.; Kashefi, J.; Cristofaro, M.; Musmeci, S.; Evidente, A. Acaricidal activity of the plant sesquiterpenoids α-costic acid and inuloxin A against the cattle ectoparasitic tick, Rhipicephalus (Boophilus) annulatus. Int. J. Acarol. 2020, 46, 409–413. [Google Scholar] [CrossRef]
- Bertin, C.; Yang, X.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Dayan, F.; Duke, S.O. Biological activity of allelochemicals. In Plant-Derived Natural Products; Osbourn, A., Lanzotti, V., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 361–384. [Google Scholar]
- Latif, S.; Chiapusio, G.; Weston, L.A. Allelopathy and the role of allelochemicals in plant defence. Adv. Bot. Res. 2017, 82, 19–54. [Google Scholar]
- Hickman, D.T.; Rasmussen, A.; Ritz, K.; Birkett, M.A.; Neve, P. Allelochemicals as multi-kingdom plant defence compounds: Towards an integrated approach. Pest Manag. Sci. 2021, 77, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; 368p. [Google Scholar]
- Macías, F.A.; Molinillo, J.M.M.G.; Varela, R.M.; Galindo, J.C.G. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Zermane, N.; Cimmino, A.; Avolio, F.; Boari, A.; Vurro, M.; Evidente, A. Inuloxins A–D, phytotoxic bi-and tri-cyclic sesquiterpene lactones produced by Inula viscosa: Potential for broomrapes and field dodder management. Phytochemistry 2013, 86, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Celik, T.; Aslanturk, O. Evaluation of cytotoxicity and genotoxicity of Inula viscosa leaf extracts with allium test. J. Biomed. Biotechnol. 2010, 2010, 189252. [Google Scholar] [CrossRef] [Green Version]
- Dor, E.; Hershenhorn, J. Allelopathic effects of Inula viscosa leaf extracts on weeds. Allelopath. J. 2012, 30, 281–289. [Google Scholar]
- Parolin, P.; Scotta, M.I.; Bresch, C. Biology of Dittrichia viscosa, a mediterranean ruderal plant: A review. Phyton Int. J. Exp. Bot. 2014, 83, 251–262. Available online: https://hal.inrae.fr/hal-02634395 (accessed on 6 December 2021).
- Alkofahi, A.; Atta, A.H. Pharmacological screening of the anti-ulcerogenic effects of some Jordanian medicinal plants in rats. J. Ethnopharmacol. 1999, 67, 341–345. [Google Scholar] [CrossRef]
- Boari, A.; Vurro, M.; Calabrese, G.J.; Mahmoud, M.N.Z.; Cazzato, E.; Fracchiolla, M. Evaluation of Dittrichia viscosa (L.) Greuter Dried Biomass for Weed Management. Plants 2021, 10, 147. [Google Scholar] [CrossRef]
- Moeini, A.; Masi, M.; Zonno, M.C.; Boari, A.; Cimmino, A.; Tarallo, O.; Evidente, A. Encapsulation of inuloxin A, a plant germacrane sesquiterpene with potential herbicidal activity, in β-cyclodextrins. Org. Biomol. Chem. 2019, 17, 2508–2515. [Google Scholar] [CrossRef]
- Cimmino, A.; Fernandez-Aparicio, M.; Andolfi, A.; Basso, S.; Rubiales, D.; Evidente, A. Effect of fungal and plant metabolites on broomrapes (Orobanche and Phelipanche spp.) seed germination and radicle growth. J. Agric. Food Chem. 2014, 62, 10485–10492. [Google Scholar] [CrossRef]
- Krishnamurthy, A.; Rathinasabapathi, B. Auxin and its transport play a role in plant tolerance to arsenite-induced oxidative stress in Arabidopsis thaliana. Plant Cell Environ. 2013, 36, 1838–1849. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, R.; Zhang, P.; Chen, Q.; Luo, Q.; Zhu, Y. The nitrification inhibitor methyl 3-(4-hydroxyphenyl) propionate modulates root development by interfering with auxin signaling via the NO/ROS pathway in Arabidopsis. Plant Physiol. 2016, 171, 1686–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Lu, L.Y.; Hu, L.Y.; Cao, W.; Sun, K.; Sun, Q.B.; Siddikee, A.; Shi, R.H.; Dai, C.C. Evidence for the involvement of auxin, ethylene and ROS signaling during primary root inhibition of Arabidopsis by the allelochemical benzoic acid. Plant Cell Physiol. 2018, 59, 1889–1904. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Chuan, Y.; Guo, C.; Liao, J.; Xu, Y.; Mei, X.; Zhu, S. Panax notoginseng root cell death caused by the autotoxic ginsenoside Rg1 is due to over-accumulation of ROS, as revealed by transcriptomic and cellular approaches. Front. Plant Sci. 2018, 9, 264. [Google Scholar] [CrossRef] [Green Version]
- Lara-Nunez, A.; Romero-Romero, T.; Ventura, J.L.; Blancas, V.; Anaya, A.L.; Cruz-Ortega, R. Allelochemical stress causes inhibition of growth and oxidative damage in Lycopersicon esculentum Mill. Plant Cell Environ. 2006, 29, 2009–2016. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in Plants: From Functions to Biofortification. Antioxidants 2019, 8, 519. [Google Scholar] [CrossRef] [Green Version]
- Vurro, M.; Boari, A.; Pilgeram, A.L.; Sands, D.C. Exogenous amino acids inhibit seed germination and tubercle formation by Orobanche ramose (Broomrape): Potential application for management of parasitic weeds. Biol. Control 2006, 36, 258–265. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol. 1996, 132, 361–373. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Adamidi, C.; Lozano, R.M.; Yee, B.C.; Momma, M.; Kobrehel, K.; Ermel, R.; Frick, O.L. Thioredoxin-linked mitigation of allergic responses to wheat. Proc. Natl. Acad. Sci. USA 1997, 94, 5372–5377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paciolla, C.; De Tullio, M.C.; Chiappetta, A.; Innocenti, A.M.; Bitonti, M.B.; Liso, R.; Arrigoni, O. Short- and Long-Term Effects of Dehydroascorbate in Lupinus albus and Allium cepa Roots. Plant Cell Physiol. 2001, 42, 857–863. [Google Scholar] [CrossRef] [Green Version]
- Mastropasqua, L.; Dipierro, N.; Paciolla, C. Effects of Darkness and Light Spectra on Nutrients and Pigments in Radish, Soybean, Mung Bean and Pumpkin Sprouts. Antioxidants 2020, 9, 558. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bhowmik, P.C. Challenges and opportunities in implementing allelopathy for natural weed management. Crop Prot. 2003, 22, 661–671. [Google Scholar] [CrossRef]
- Scavo, A.; Mauromicale, G. Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future. Agronomy 2021, 11, 2104. [Google Scholar] [CrossRef]
- Duke, O.S. Proving Allelopathy in crop-weed interactions. Weed Sci. 2015, 63, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Fumagalli, P.; Andolfi, A.; Avolio, F.; Boari, A.; Cimmino, A.; Finizio, A. Ecotoxicological characterisation of a mycoherbicide mixture isolated from the fungus Ascochyta caulina. Pest Manag. Sci. 2013, 69, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Cordeau, S.; Triolet, M.; Wayman, S.; Steinberg, C.; Guillemin, J.P. Bioherbicides: Dead in the water? A review of the existing products for integrated weed management. Crop Prot. 2016, 87, 44–49. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Sabetta, W.; Paradiso, A.; Paciolla, C.; de Pinto, M.C. Chemistry, Biosynthesis, and Antioxidative Function of Glutathione in Plants. In Glutathione in Plant Growth, Development, and Stress Tolerance; Hossain, M.A., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–27. ISBN 978-3-319-66681-5. [Google Scholar]
- Matsui, T.; Kitagawa, Y.; Okumura, M.; Shigeta, Y. Accurate standard hydrogen electrode potential and applications to the redox potentials of vitamin C and NAD/NADH. J. Phys. Chem. A 2015, 119, 369–376. [Google Scholar] [CrossRef]
- Bilska, A.; Kobus-Cisowska, J.; Kmiecik, D.; Danyluk, B.; Kowalski, R.; Szymanowska, D.; Gramza-Michałowska, A.; Szczepaniak, O. Cholinesterase inhibitory activity, antioxidative potential and microbial stability of innovative liver pâté fortified with rosemary extract (Rosmarinus officinalis). Electron. J. Biotechnol. 2019, 40, 22–29. [Google Scholar] [CrossRef]
- Fiorani, M.; De Sanctis, R.; Scarlatti, F.; Vallorani, L.; De Bellis, R.; Serafini, G.; Bianchi, M.; Stocchi, V. Dehydroascorbic acid irreversibly inhibits hexokinase activity. Mol. Cell. Biochem. 2000, 209, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Cordoba-Pedregosa, M.D.C.; Gonzalez-Reyes, J.A.; Canadillas, M.D.S.; Navas, P.; Cordoba, F. Role of apoplastic and cell-wall peroxidases on the stimulation of root elongation by ascorbate. Plant Physiol. 1996, 112, 1119–1125. [Google Scholar] [CrossRef] [Green Version]
- Sano, S. Molecular and functional characterization of monodehydro-ascorbate and dehydroascorbate reductases. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M.A., Munné-Bosch, S., Burritt, D.J., Diaz-Vivancos, P., Fujita, M., Lorence, A., Eds.; Springer: Cham, Switzerland, 2017; Volume 1, pp. 129–156. [Google Scholar]
- Hong, Y.; Hu, H.Y.; Xie, X.; Li, F.M. Responses of enzymatic antioxidants and non-enzymatic antioxidants in the cyanobacterium Microcystis aeruginosa to the allelochemical ethyl 2-methyl acetoacetate (EMA) isolated from reed (Phragmites communis). J. Plant Physiol. 2008, 165, 1264–1273. [Google Scholar] [CrossRef]
- Wang, J.; Zhong, X.; Li, F.; Shi, Z. Effects of nicosulfuron on growth, oxidative damage, and the ascorbate-glutathione pathway in paired nearly isogenic lines of waxy maize (Zea mays L.). Pestic. Biochem. Phys. 2018, 145, 108–117. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; De Leonardis, S.; Mulè, G.; Logrieco, A.F.; Paciolla, C. A Novel and Potentially Multifaceted Dehydroascorbate Reductase Increasing the Antioxidant Systems is Induced by Beauvericin in Tomato. Antioxidants 2020, 9, 435. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days | Treatment | Lycopersicon esculentum | Lepidium sativum | ||
|---|---|---|---|---|---|
| Root Length (mm) | Root Length Inhibition (%) a | Root Length (mm) | Root Length Inhibition (%) a | ||
| 3 | 10−5 M | 34.8 ± 2.92 | 4.13 | 21.2 ± 0.19 | 9.97 |
| 10−4 M | 26.8 ± 1.82 | 26.2 | 19.9 ± 2.21 | 15.5 | |
| 10−3 M | 14.1 ± 0.45 ** | 61.2 | 13.2 ± 1.07 *** | 44.1 | |
| control | 36.3 ± 0.22 | - | 23.6 ± 1.56 | - | |
| 6 | 10−5 M | 65.1 ± 3.82 | 0.04 | 34.2 ± 1.63 | 0.91 |
| 10−4 M | 52.8 ± 5.94 | 18.9 | 20.3 ± 2.97 * | 41.2 | |
| 10−3 M | 13.9 ± 1.20 *** | 78.6 | 3.73 ± 1.81 ** | 89.2 | |
| control | 65.2 ± 4.67 | - | 34.6 ± 2.76 | - | |
| Days | Treatment | Lycopersicon esculentum | Lepidium sativum | ||
|---|---|---|---|---|---|
| Weight (mg) | Weight Inhibition (%) a | Weight (mg) | Weight Inhibition (%) a | ||
| 3 | 10−5 M | 18.8 ± 4.66 | 12.1 | 13.8 ± 0.44 | 14.4 |
| 10−4 M | 16.7 ± 0.55 | 22.3 | 11.6 ± 1.20 * | 28.1 | |
| 10−3 M | 13.3 ± 0.71 ** | 37.7 | 10.6 ± 0.28 ** | 34.4 | |
| control | 21.4 ± 0.84 | - | 16.1 ± 1.35 | - | |
| 6 | 10−5 M | 25.0 ± 3.30 | 0.62 | 17.6 ± 0.16 | 4.19 |
| 10−4 M | 22.7 ± 1.16 | 9.62 | 16.1 ± 0.41 | 12.3 | |
| 10−3 M | 17.0 ± 0.35 ** | 32.4 | 9.16 ± 0.99 *** | 50.2 | |
| control | 25.2 ± 0.74 | - | 18.4 ± 0.98 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villani, A.; Zonno, M.C.; de Leonardis, S.; Vurro, M.; Paciolla, C. Inuloxin A Inhibits Seedling Growth and Affects Redox System of Lycopersicon esculentum Mill. and Lepidium sativum L. Biomolecules 2022, 12, 302. https://doi.org/10.3390/biom12020302
Villani A, Zonno MC, de Leonardis S, Vurro M, Paciolla C. Inuloxin A Inhibits Seedling Growth and Affects Redox System of Lycopersicon esculentum Mill. and Lepidium sativum L. Biomolecules. 2022; 12(2):302. https://doi.org/10.3390/biom12020302
Chicago/Turabian StyleVillani, Alessandra, Maria Chiara Zonno, Silvana de Leonardis, Maurizio Vurro, and Costantino Paciolla. 2022. "Inuloxin A Inhibits Seedling Growth and Affects Redox System of Lycopersicon esculentum Mill. and Lepidium sativum L." Biomolecules 12, no. 2: 302. https://doi.org/10.3390/biom12020302
APA StyleVillani, A., Zonno, M. C., de Leonardis, S., Vurro, M., & Paciolla, C. (2022). Inuloxin A Inhibits Seedling Growth and Affects Redox System of Lycopersicon esculentum Mill. and Lepidium sativum L. Biomolecules, 12(2), 302. https://doi.org/10.3390/biom12020302