
Determination of the Absolute Molar Mass of [Fe-S]-Containing Proteins Using Size Exclusion Chromatography-Multi-Angle Light Scattering (SEC-MALS)

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
3. Theoretical Background
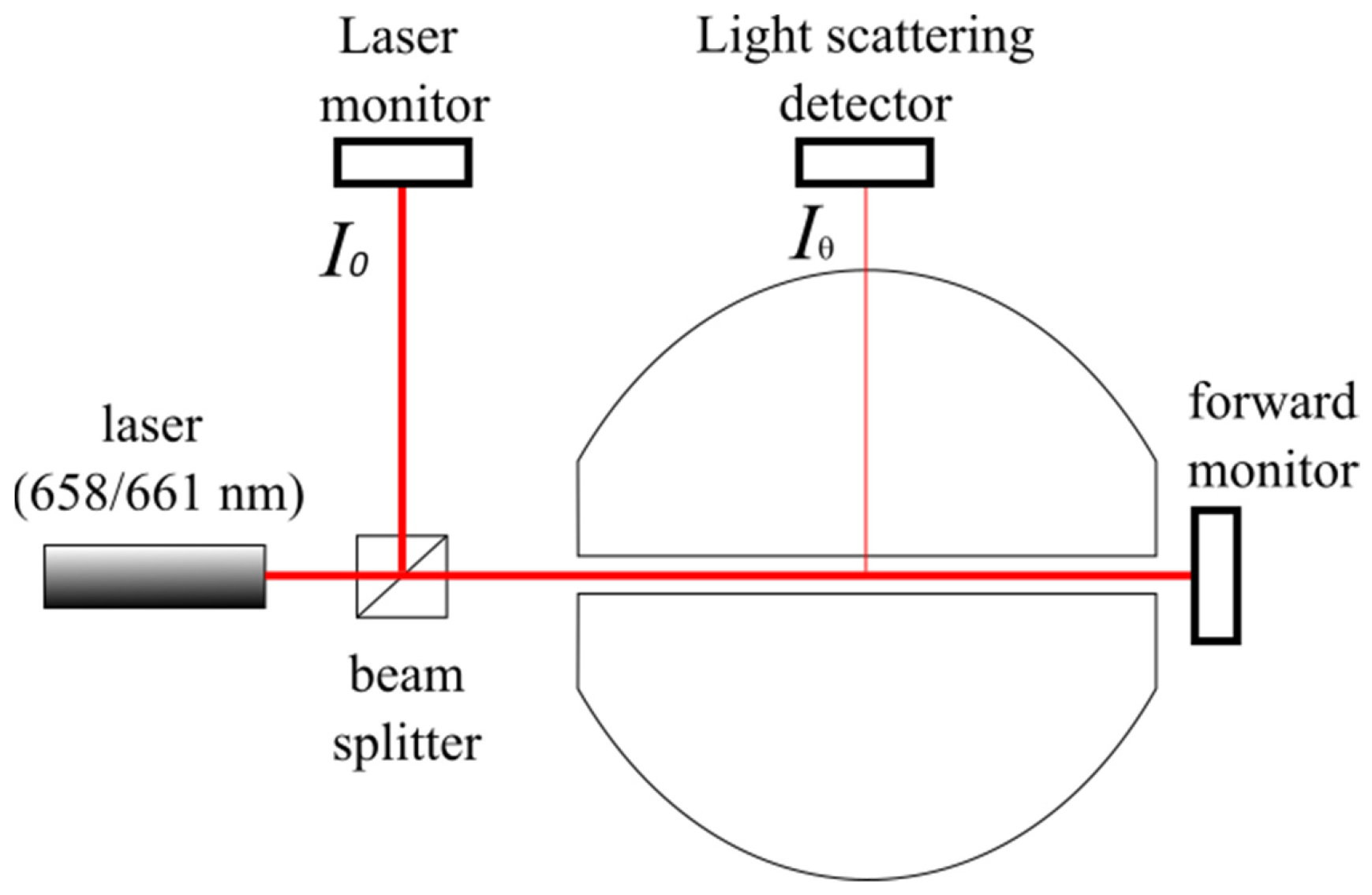
3.1. SEC-MALS Experiment
3.2. Forward Monitor Correction Mode for Samples Absorbing at the Laser Wavelength
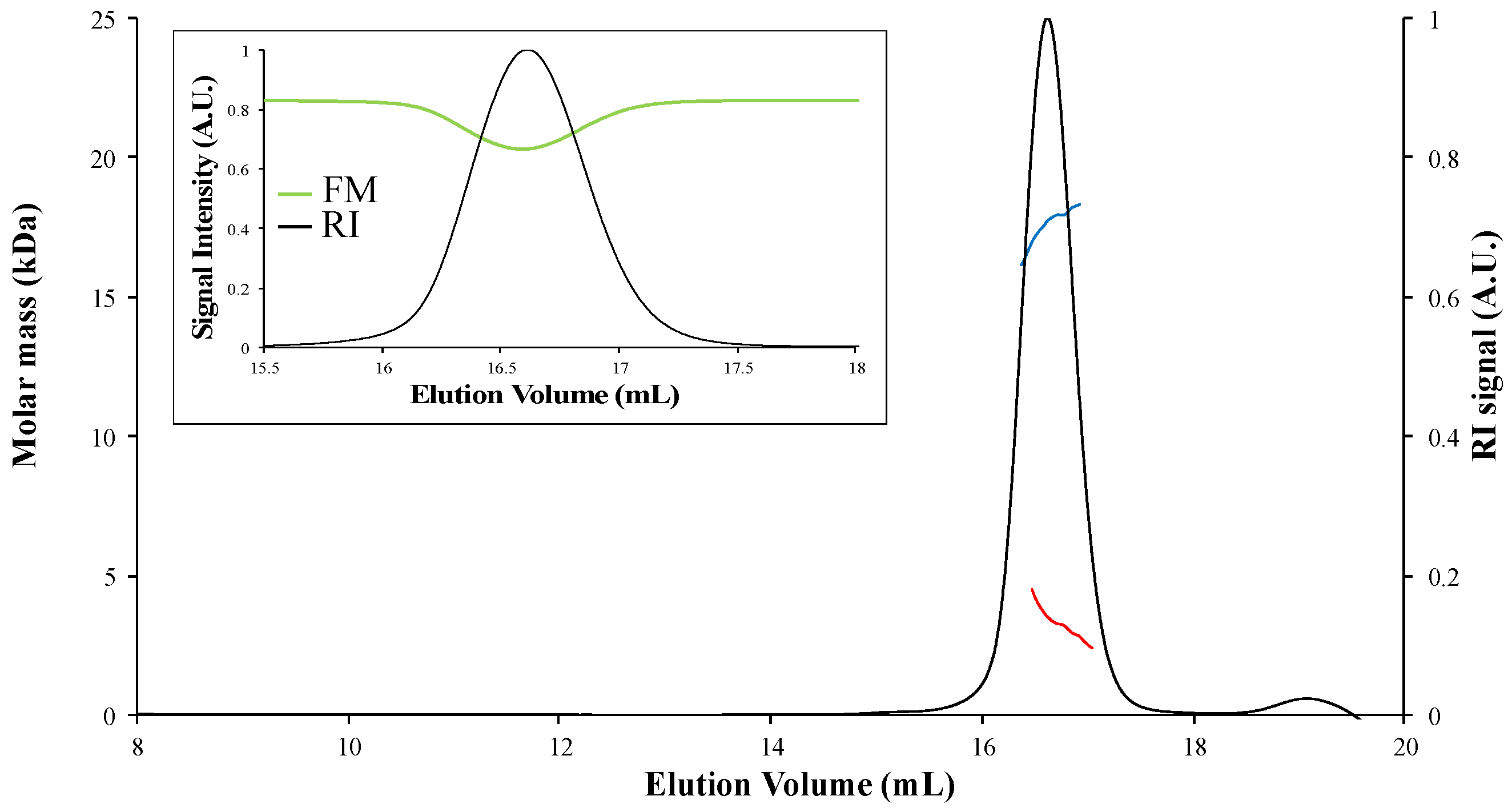
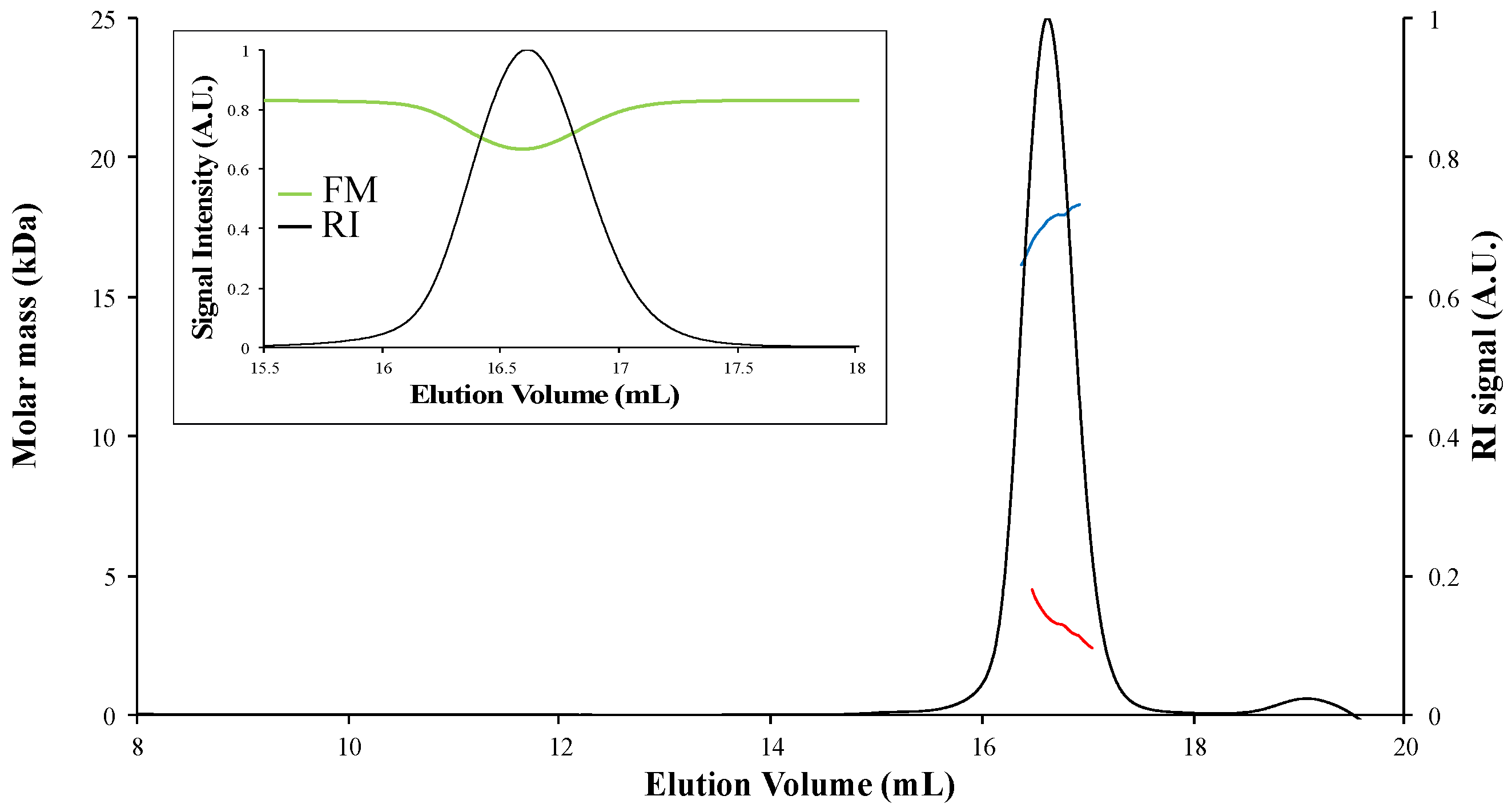
4. Results
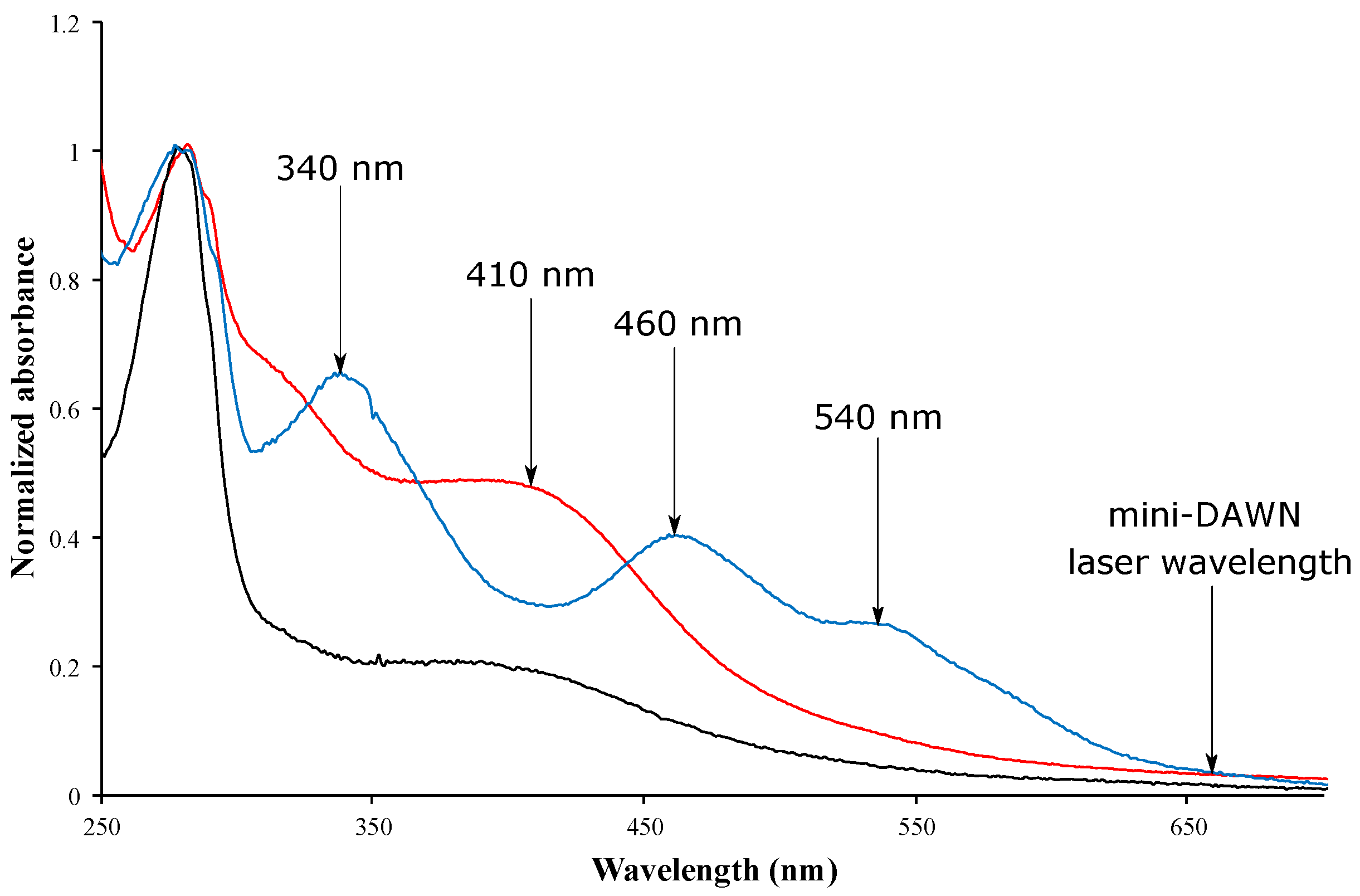
4.1. [Fe-S]-Binding Proteins Absorb at the Laser Wavelength
4.2. Corrections for Samples Absorbing at the Laser Wavelength
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uliyanchenko, E. Size-exclusion chromatography-from high-performance to ultra-performance. Anal. Bioanal. Chem. 2014, 406, 6087–6094. [Google Scholar] [CrossRef]
- Wyatt, P.J. Light scattering and the absolute characterization of macromolecules. Anal. Chim. Acta 1993, 272, 1–40. [Google Scholar] [CrossRef]
- Wen, J.; Arakawa, T.; Philo, J.S. Size-exclusion chromatography with on-line light-scattering, absorbance, and refractive index detectors for studying proteins and their interactions. Anal. Biochem. 1996, 240, 155–166. [Google Scholar] [CrossRef]
- Adawy, A.; Groves, M.R. The Use of Size Exclusion Chromatography to Monitor Protein Self-Assembly. Crystals 2017, 7, 331. [Google Scholar] [CrossRef] [Green Version]
- Some, D.; Amartely, H.; Tsadok, A.; Lebendiker, M. Characterization of Proteins by Size-Exclusion Chromatography Coupled to Multi-Angle Light Scattering (SEC-MALS). J. Vis. Exp. 2019, 148, e59615. [Google Scholar] [CrossRef] [Green Version]
- Hardouin, P.; Velours, C.; Bou-Nader, C.; Assrir, N.; Laalami, S.; Putzer, H.; Durand, D.; Golinelli-Pimpaneau, B. Dissociation of the Dimer of the Intrinsically Disordered Domain of RNase Y upon Antibody Binding. Biophys. J. 2018, 115, 2102–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serebryany, E.; Folta-Stogniew, E.; Liu, J.; Yan, E.C. Homodimerization enhances both sensitivity and dynamic range of the ligand-binding domain of type 1 metabotropic glutamate receptor. FEBS Lett. 2016, 590, 4308–4317. [Google Scholar] [CrossRef] [Green Version]
- Proença, D.; Velours, C.; Leandro, C.; Garcia, M.; Pimentel, M.; Sao-Jose, C. A two-component, multimeric endolysin encoded by a single gene. Mol. Microbiol. 2015, 95, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Loiseau, L.; Fyfe, C.; Aussel, L.; Hajj Chehade, M.; Hernandez, S.B.; Faivre, B.; Hamdane, D.; Mellot-Draznieks, C.; Rascalou, B.; Pelosi, L.; et al. The UbiK protein is an accessory factor necessary for bacterial ubiquinone (UQ) biosynthesis and forms a complex with the UQ biogenesis factor UbiJ. J. Biol. Chem. 2017, 292, 11937–11950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hastie, K.; Rayaprolu, V.; Saphire, E.O. Analysis of Oligomeric and Glycosylated Proteins by Size-Exclusion Chromatography Coupled with Multiangle Light Scattering. Methods Mol. Biol. 2021, 2271, 343–359. [Google Scholar] [CrossRef]
- Zinovyev, G.; Sulaeva, I.; Podzimek, S.; Rössner, D.; Kilpeläinen, I.; Sumerskii, I.; Rosenau, T.; Potthast, A. Getting Closer to Absolute Molar Masses of Technical Lignins. ChemSusChem 2018, 11, 3259–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreini, C.; Rosato, A.; Banci, L. The relationship between environmental dioxygen and iron-sulfur proteins explored at the genome level. PLoS ONE 2017, 12, e0171279. [Google Scholar] [CrossRef] [PubMed]
- Beinert, H. Iron-sulfur proteins: Ancient structures, still full of surprises. J. Biol. Inorg. Chem. 2000, 5, 2–15. [Google Scholar] [CrossRef]
- Dinis, P.; Wieckowski, B.M.; Roach, P.L. Metallocofactor assembly for [FeFe]-hydrogenases. Curr. Opin. Struct. Biol. 2016, 41, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Jeoung, J.H.; Martins, B.M.; Dobbek, H. Double-Cubane [8Fe9S] Clusters: A Novel Nitrogenase-Related Cofactor in Biology. Chembiochem 2020, 21, 1710–1716. [Google Scholar] [CrossRef]
- Yakovlev, G.; Reda, T.; Hirst, J. Reevaluating the relationship between EPR spectra and enzyme structure for the iron sulfur clusters in NADH:quinone oxidoreductase. Proc. Natl. Acad. Sci. USA 2007, 104, 12720–12725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crack, J.C.; Green, J.; Thomson, A.J.; Le Brun, N.E. Iron-sulfur clusters as biological sensors: The chemistry of reactions with molecular oxygen and nitric oxide. Acc. Chem. Res. 2014, 47, 3196–3205. [Google Scholar] [CrossRef]
- Bouton, C.; Drapier, J.C. Iron regulatory proteins as NO signal transducers. Sci. STKE 2003, 2003, pe17. [Google Scholar] [CrossRef] [PubMed]
- Crack, J.C.; Green, J.; Thomson, A.J.; Le Brun, N.E. Iron-sulfur cluster sensor-regulators. Curr. Opin. Chem. Biol. 2012, 16, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Mettert, E.L.; Kiley, P.J. Fe-S proteins that regulate gene expression. Biochim. Biophys. Acta 2015, 1853, 1284–1293. [Google Scholar] [CrossRef] [Green Version]
- Crack, J.C.; Jervis, A.J.; Gaskell, A.A.; White, G.F.; Green, J.; Thomson, A.J.; Le Brun, N.E. Signal perception by FNR: The role of the iron-sulfur cluster. Biochem. Soc. Trans. 2008, 36, 1144–1148. [Google Scholar] [CrossRef]
- Mettert, E.L.; Kiley, P.J. Reassessing the Structure and Function Relationship of the O2 Sensing Transcription Factor FNR. Antioxid. Redox Signal. 2018, 29, 1830–1840. [Google Scholar] [CrossRef]
- Flint, D.H.; Allen, R.M. Iron-Sulfur Proteins with Nonredox Functions. Chem. Rev. 1996, 96, 2315–2334. [Google Scholar] [CrossRef]
- Beinert, H.; Kennedy, M.C.; Stout, C.D. Aconitase as Iron-Sulfur Protein, Enzyme, and Iron-Regulatory Protein. Chem. Rev. 1996, 96, 2335–2374. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.; Tórtora, V.; Mansilla, S.; Radi, R. Aconitases: Non-redox Iron-Sulfur Proteins Sensitive to Reactive Species. Acc. Chem. Res. 2019, 52, 2609–2619. [Google Scholar] [CrossRef]
- Salameh, M.; Riquier, S.; Guittet, O.; Huang, M.E.; Vernis, L.; Lepoivre, M.; Golinelli-Cohen, M.P. New Insights of the NEET Protein CISD2 Reveals Distinct Features Compared to Its Close Mitochondrial Homolog mitoNEET. Biomedicines 2021, 9, 384. [Google Scholar] [CrossRef]
- Karmi, O.; Marjault, H.B.; Pesce, L.; Carloni, P.; Onuchic, J.N.; Jennings, P.A.; Mittler, R.; Nechushtai, R. The unique fold and lability of the [2Fe-2S] clusters of NEET proteins mediate their key functions in health and disease. J. Biol. Inorg. Chem. 2018, 23, 599–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golinelli-Cohen, M.-P.; Lescop, E.; Mons, C.; Goncalves, S.; Clemancey, M.; Santolini, J.; Guittet, E.; Blondin, G.; Latour, J.M.; Bouton, C. Redox Control of the Human Iron-Sulfur Repair Protein MitoNEET Activity via Its Iron-Sulfur Cluster. J. Biol. Chem. 2016, 291, 7583–7593. [Google Scholar] [CrossRef] [Green Version]
- Mons, C.; Botzanowski, T.; Nikolaev, A.; Hellwig, P.; Cianférani, S.; Lescop, E.; Bouton, C.; Golinelli-Cohen, M.-P. The H2O2-resistant Fe-S redox switch mitoNEET acts as a pH sensor to repair stress-damaged Fe-S protein. Biochemistry 2018, 57, 5616–5628. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Asai, S.I.; Narai, S.; Nambu, S.; Omura, N.; Sakaguchi, Y.; Suzuki, T.; Ikeda-Saito, M.; Watanabe, K.; Yao, M.; et al. Biochemical and structural characterization of oxygen-sensitive 2-thiouridine synthesis catalyzed by an iron-sulfur protein TtuA. Proc. Natl. Acad. Sci. USA 2017, 114, 4954–4959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arragain, S.; Bimai, O.; Legrand, P.; Caillat, S.; Ravanat, J.L.; Touati, N.; Binet, L.; Atta, M.; Fontecave, M.; Golinelli-Pimpaneau, B. Nonredox thiolation in tRNA occurring via sulfur activation by a [4Fe-4S] cluster. Proc. Natl. Acad. Sci. USA 2017, 114, 7355–7360. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Lénon, M.; Touati, N.; Ravanat, J.-L.; Velours, C.; Fontecave, M.; Barras, F.; Golinelli-Pimpaneau, B. Iron sulfur biology invades tRNA modification: The case of U34 sulfuration. Nucleic Acids Res. 2021, 49, 3997–4007. [Google Scholar] [CrossRef]
- Shigi, N. Biosynthesis and Degradation of Sulfur Modifications in tRNAs. Int. J. Mol. Sci. 2021, 22, 11937. [Google Scholar] [CrossRef]
- Fellner, M.; Desguin, B.; Hausinger, R.P.; Hu, J. Structural insights into the catalytic mechanism of a sacrificial sulfur insertase of the N-type ATP pyrophosphatase family, LarE. Proc. Natl. Acad. Sci USA 2017, 114, 9074–9079. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Pecqueur, L.; Aučynaitė, A.; Fuchs, J.; Rutkienė, R.; Vaitekūnas, J.; Meškys, R.; Boll, M.; Fontecave, M.; Urbonavičius, J.; et al. Structural evidence for a [4Fe-5S] intermediate in the non-redox desulfuration of thiouracil. Angew. Chem. Int. Ed. Engl. 2021, 60, 424–431. [Google Scholar] [CrossRef]
- Tchong, S.I.; Xu, H.; White, R.H. L-cysteine desulfidase: An [4Fe-4S] enzyme isolated from Methanocaldococcus jannaschii that catalyzes the breakdown of L-cysteine into pyruvate, ammonia, and sulfide. Biochemistry 2005, 44, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- Mons, C.; Ferecatu, I.; Riquier, S.; Lescop, E.; Bouton, C.; Golinelli-Cohen, M.-P. Combined biochemical, biophysical and cellular methods to study Fe-S cluster transfer and cytosolic aconitase repair by mitoNEET. Methods Enzymol. 2017, 595, 83–108. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Brown, P.H.; Schuck, P. On the distribution of protein refractive index increments. Biophys. J. 2011, 100, 2309–2317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorber, B. Analytical light scattering methods in molecular and structural biology: Experimental aspects and results. arXiv 2018, arXiv:1810.00611. [Google Scholar]
- Technical Note TN3000, Wyatt Technology, Calibration and Normalization of MALS Detectors. Available online: https://chem.uiowa.edu/sites/chem.uiowa.edu/files/groups/TN3000%20-%20Calibration%20and%20Normalization%20of%20MALS%20Detectors%20Rev%20A.pdf (accessed on 2 February 2022).
- Freibert, S.A.; Weiler, B.D.; Bill, E.; Pierik, A.J.; Mühlenhoff, U.; Lill, R. Biochemical Reconstitution and Spectroscopic Analysis of Iron–Sulfur Proteins. Methods Enzymol. 2018, 599, 197–226. [Google Scholar] [CrossRef]
- Ferecatu, I.; Goncalves, S.; Golinelli-Cohen, M.P.; Clemancey, M.; Martelli, A.; Riquier, S.; Guittet, E.; Latour, J.M.; Puccio, H.; Drapier, J.C.; et al. The diabetes drug target MitoNEET governs a novel trafficking pathway to rebuild an Fe-S cluster into cytosolic aconitase/iron regulatory protein 1. J. Biol. Chem. 2014, 289, 28070–28086. [Google Scholar] [CrossRef] [Green Version]
- Paddock, M.L.; Wiley, S.E.; Axelrod, H.L.; Cohen, A.E.; Roy, M.; Abresch, E.C.; Capraro, D.; Murphy, A.N.; Nechushtai, R.; Dixon, J.E.; et al. MitoNEET is a uniquely folded 2Fe 2S outer mitochondrial membraneprotein stabilized by pioglitazone. Proc. Natl. Acad. Sci. USA 2007, 104, 14342–14347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Liu, R.; Ross, S.; Smart, E.J.; Zhu, H.; Gong, W. Crystallographic studies of human MitoNEET. J. Biol. Chem. 2007, 282, 33242–33246. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Zhou, T.; Ye, K.; Wang, J. Crystal structure of human mitoNEET reveals distinct groups of iron sulfur proteins. Proc. Natl. Acad. Sci. USA 2007, 104, 14640–14645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamir, S.; Eisenberg-Domovich, Y.; Conlan, A.R.; Stofleth, J.T.; Lipper, C.H.; Paddock, M.L.; Mittler, R.; Jennings, P.A.; Livnah, O.; Nechushtai, R.A. A point mutation in the [2Fe-2S] cluster binding region of the NAF-1 protein (H114C) dramatically hinders the cluster donor properties. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1572–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freibert, S.A.; Boniecki, M.T.; Stümpfig, C.; Schulz, V.; Krapoth, N.; Winge, D.R.; Mühlenhoff, U.; Stehling, O.; Cygler, M.; Lill, R. N-terminal tyrosine of ISCU2 triggers [2Fe-2S] cluster synthesis by ISCU2 dimerization. Nat. Commun. 2021, 12, 6902. [Google Scholar] [CrossRef] [PubMed]
- Camponeschi, F.; Prusty, N.R.; Heider, S.A.E.; Ciofi-Baffoni, S.; Banci, L. GLRX3 Acts as a [2Fe-2S] Cluster Chaperone in the Cytosolic Iron-Sulfur Assembly Machinery Transferring [2Fe-2S] Clusters to NUBP1. J. Am. Chem. Soc. 2020, 142. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
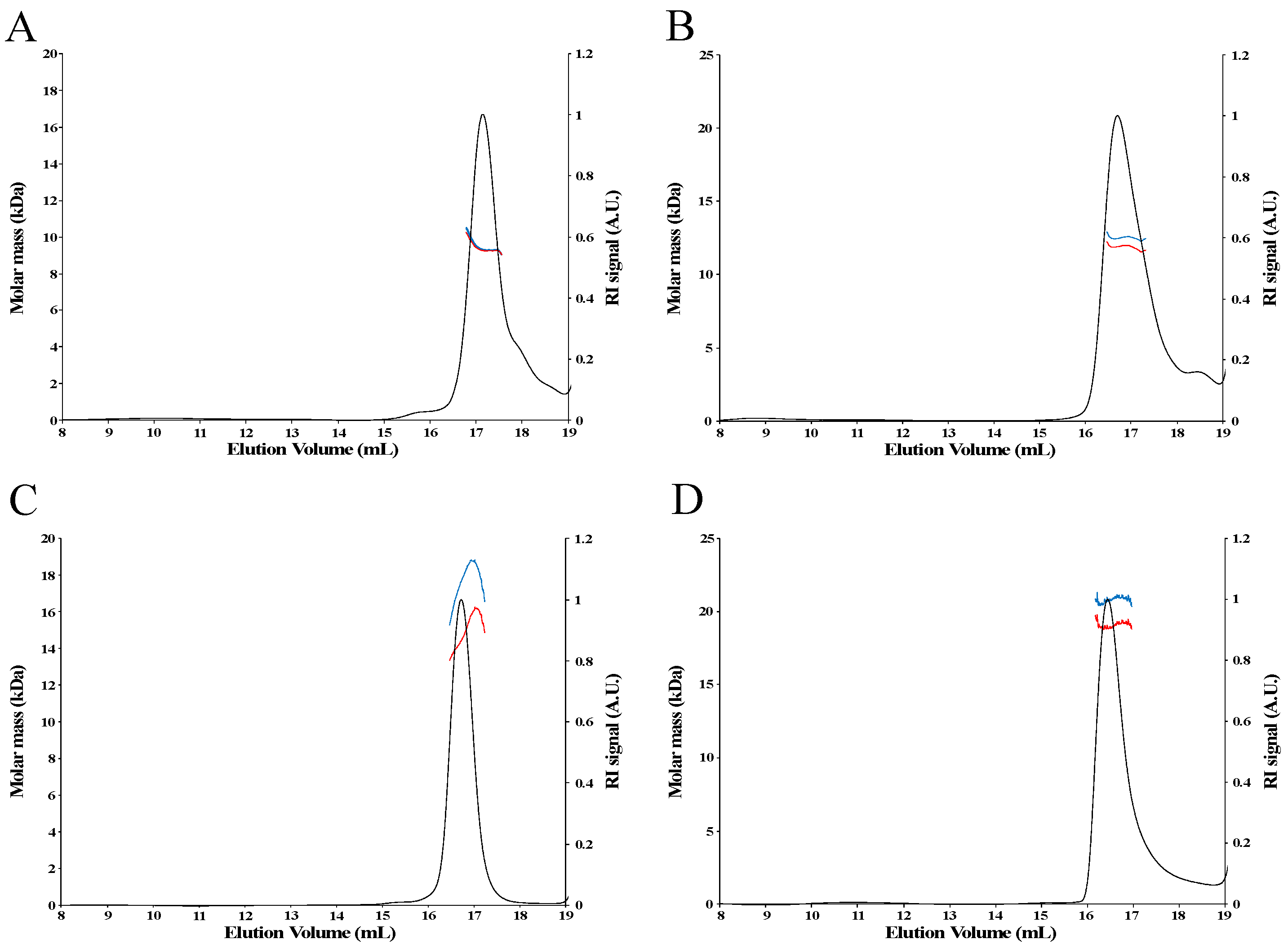{kind=link}
| Experimental Molar Mass | ||||||||
|---|---|---|---|---|---|---|---|---|
| Protein | Theoretical Molar Mass (kDa) 1 | No Correction (kDa) | Oligomerization State 1 | LM 2 (kDa) | Mass Difference with Theoretical Value Using LM 3 (%) | FM 2 (kDa) | Mass Difference with Theoretical Value Using FM 3 (%) | |
| mitoNEET | 8.6 (M); 17.2 (D) | Apo | 9.3 | M | 9.3 | −8.1 | 9.3 | −8.1 |
| Holo | 16.2 | D | 16.2 | 5.8 | 18.8 | 9.3 | ||
| CISD2 | 11.4 (M); 22.8 (D) | Apo | 11.8 | M | 11.9 | −4.4 | 12.6 | −10.5 |
| Holo | 19 | D | 19 | 16.7 | 20.9 | 8.3 | ||
| MnmA | 40.6 (M) | Apo | 36.3 | M 4 | 36.3 | 10.6 | 36.3 | 10.6 |
| Holo | 36.1 | M | 36.5 | 10.1 | 40.1 | 1.2 | ||
| LarE | 31.1 (M); 62.2 (D) | Apo | 54.1 | D | 54.2 | 12.9 | 54.2 | 12.9 |
| Holo | 58.8 | D | 58.8 | 5.5 | 60.5 | 2.7 | ||
| CyuA | 43.7 (M); 87.4 (D) | Apo | 77.9 | D | 77.9 | 10.9 | 78.6 | 10.1 |
| Holo | 78.9 | D | 78.9 | 9.7 | 80.8 | 7.6 | ||
| TudS | 16.5 (M) | Holo | 4.5 | M | 3.3 | 80.0 | 17.9 | −8.5 |
| Protein Name | Theoretical Extinction Coefficient mL/mg·cm | Experimental Extinction Coefficient mL/mg·cm | |
|---|---|---|---|
| mitoNEET | Holo | 0.815 | 1.545 |
| Apo | 0.846 | ||
| CISD2 | Holo | 0.616 | 1.2262 |
| Apo | 0.768 | ||
| MnmA | Holo | 0.993 | 1.46 |
| Apo | 1.1 | ||
| LarE | Holo | 0.473 1 | 0.638 2 |
| Apo | 0.565 | ||
| CyuA | Holo | 0.583 | 0.858 |
| Apo | 0.733 | ||
| TudS | Holo | 0.514 | 1.406 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velours, C.; Zhou, J.; Zecchin, P.; He, N.; Salameh, M.; Golinelli-Cohen, M.-P.; Golinelli-Pimpaneau, B. Determination of the Absolute Molar Mass of [Fe-S]-Containing Proteins Using Size Exclusion Chromatography-Multi-Angle Light Scattering (SEC-MALS). Biomolecules 2022, 12, 270. https://doi.org/10.3390/biom12020270
Velours C, Zhou J, Zecchin P, He N, Salameh M, Golinelli-Cohen M-P, Golinelli-Pimpaneau B. Determination of the Absolute Molar Mass of [Fe-S]-Containing Proteins Using Size Exclusion Chromatography-Multi-Angle Light Scattering (SEC-MALS). Biomolecules. 2022; 12(2):270. https://doi.org/10.3390/biom12020270
Chicago/Turabian StyleVelours, Christophe, Jingjing Zhou, Paolo Zecchin, Nisha He, Myriam Salameh, Marie-Pierre Golinelli-Cohen, and Béatrice Golinelli-Pimpaneau. 2022. "Determination of the Absolute Molar Mass of [Fe-S]-Containing Proteins Using Size Exclusion Chromatography-Multi-Angle Light Scattering (SEC-MALS)" Biomolecules 12, no. 2: 270. https://doi.org/10.3390/biom12020270
APA StyleVelours, C., Zhou, J., Zecchin, P., He, N., Salameh, M., Golinelli-Cohen, M.-P., & Golinelli-Pimpaneau, B. (2022). Determination of the Absolute Molar Mass of [Fe-S]-Containing Proteins Using Size Exclusion Chromatography-Multi-Angle Light Scattering (SEC-MALS). Biomolecules, 12(2), 270. https://doi.org/10.3390/biom12020270