1. Introduction
Bone, which is the firm organ of the body, establishes the skeleton and supports the musculoskeletal system [
1]. The resorption and formation of bone, which includes osteoclasts and osteoblasts, respectively, are stable for the maintenance of normal skeletal structure, function, and mineral homeostasis. Once the state was disturbed, several bone-related diseases will occur [
2,
3]. The osteoclast is a multinucleated cell that originated from the hematopoietic progenitor of the monocyte–macrophage lineage, which fuse together to become tartrate-resistant acid phosphatase (TRAP)-positive multinucleated cells that act on the bone resorption while osteoblast participates in bone formation and mineralization, which forms a protective layer over the surface of bone [
1].
Reactive oxidant species (ROS), which are known for their cytotoxicity and damage, play a crucial role in regulating the molecular signals of osteoclast differentiation and ROS activate osteoclast differentiation through the RANKL signaling pathway [
4]. The endoplasmic reticulum (ER) is a critical organelle in eukaryotic cells responsible for protein folding, maturation, quality control, and trafficking. ER stress is the state of the protein folding process interruption and unfolded or misfolded protein accumulation by pathologic stimuli responses including oxidative stress, and ROS can regulate the function of ER [
5,
6]. It is reported that during RANKL-induced osteoclastogenesis, the production of intracellular ROS is stimulated by a signal cascade, ROS has a function as an intracellular signaling medium for cell differentiation into osteoclasts, and ER stress results from oxidative stress caused by ROS during osteoclastogenesis [
6]. Besides, inflammation has an important role in skewing bone remodeling toward resorption through the interaction of inflammatory mediators and their related peptides with osteoblasts, osteoclasts, and other immune cells to change the expression of RANK and RANKL, which leads to several bone diseases, such as alveolar bone erosion, periodontitis, rheumatoid arthritis, and septic prosthetic loosening [
7].
Molecular docking is an effective computational method for in silico screening, important for the development of drug design [
8,
9]. Based on the energy and geometrics, docking virtually predicts the binding between two or more binding partners as biological macromolecules (such as protein) or small molecules (such as ligands and drugs) [
8,
9]. For non-covalent binders, different types of protein–ligand interactions include ionic interactions, hydrogen bonds, and van der Waals interactions in which dispersion, polar, and induced interactions are involved [
9]. Among many systemic hormones and cytokines involved in osteoclast differentiation and function, the receptor activator of nuclear factor (NF)-κB ligands (RANKL) is a key osteoclastogenic molecule [
10,
11]. Previous research reported that baicalin specifically binds to the active site of RANKL and inhibits osteoporosis by regulation of the RANK/RANKL/OPG pathway in vitro and in vivo through bone modeling transcription factors, such as nuclear factor κB2 (nfκb2), TNF receptor-associated factor 6 (traf6), forkhead box O1 a2 (foxo1a2), mitogen-activated protein kinase 8b (mapk8b), TNF superfamily member 13b (tnfsl3b), and bone tissue mineralization, growth, and development in the dexamethasone (DEX)-induced osteoporosis zebrafish larvae model [
12]. Therefore, it is important to understand the interaction between RANKL and secondary compounds for further treatment of osteoclast differentiation and bone diseases.
Natural products play an important role in the development for the treatment of many diseases, and several plant extracts and metabolites exhibit pharmaceutical activities [
13].
Prunus armeniaca L., a fragrant and flavorful apricot belonging to the Rosaceae family, has its origins in Central Asia and is widely distributed around the world [
14,
15]. According to published reports, many secondary metabolites, such as steroids, norisoprenoids, terpenoids, proanthocyanidins, and phenolic compounds, have been isolated and structurally elucidated in many parts of
P. armeniaca [
16,
17]. Amygdalin (AD), a cyanogenic glycoside isolated from
P. armeniaca, was reported to exhibit various pharmacological effects, such as anti-fibrosis, anti-inflammation, analgesia, auxiliary anticancer, immunoregulation, anti-atherosclerosis, anti-cardiac hypertrophy, anti-ulcer, hypoglycemia effects, anticholinesterase, and neuroprotective activities [
18,
19,
20,
21]. It is reported that the high concentration of AD (10 mg/mL) reduced the viability and size of osteoblasts, decreased extracellular matrix mineralization by down-regulating the COL1A1 and ALPL genes, and enhanced osteoclast formation and bone resorption by up-regulating the TNFSF11 and WNT5A genes while increasing the expression of BGLAP gene. In lower concentrations (0.1 mg/mL), there is almost no effect on functions of human osteoblasts in vitro [
22]. A recent study reported that by using MSC-specific transforming growth factor- (TGF-) β receptor 2 conditional knockout (KO) mice (Tgfbr2Gli1-Cre) and C3H10 T1/2 murine mesenchymal progenitor cells, the migration and differentiation of mesenchymal stem cells (MSCs) accelerated the fracture healing process by AD through the TGF-β/Smad signaling pathway, supporting the use of amygdalin-based therapy for fracture healing [
23]. However, as reported, there is no information about the effect of AD on osteoclast differentiation and function.
In this study, we explored the binding of AD and RANKL through molecular docking simulation and investigated the effects of AD on osteoclast differentiation and function through the RANKL-induced RAW264.7 in vitro model.
2. Materials and Methods
2.1. Reagents and Chemicals
Recombinant soluble RANKL ligand (sRANKL) was acquired from PeproTech EC Ltd. (London, UK). An Acid Phosphatase Assay Kit, 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), leukocyte acid phosphatase, 4-6-diamidino-2-phenylindole (DAPI), ascorbic acid, and β-glycerophosphate were purchased from Sigma-Aldrich Fine Chemicals (Saint Louis, MO, USA). Dulbecco’s modified Eagle’s medium (DMEM), minimum essential medium alpha (α-MEM), and fetal bovine serum (FBS) were purchased from Welgene Bioscience (Daegu, Korea). TRIZOL reagent was purchased from JBI (Seoul, Korea) and TOPscript™ RT DryMIX (dT18 plus) was purchased from Enzynomics (Daejeon, Korea). TB Green® Pre-mix Ex Taq™ II (Tli RNaseH Plus) was purchased from Takara (Tokyo, Japan). Primary antibodies for phospho-JNK, JNK, phospho-p38, p38, phospho-ERK, ERK, BIP, p-eIF2α, eIF2α, and rabbit polyclonal antibodies were purchased from Cell Signaling Technology Inc. (Danvers, MA, USA). Catalase (CAT) and superoxide dismutase (SOD) antibodies were acquired from BD biosciences (San Jose, CA, USA). An anti-nuclear factor of activated T cells-c1 (NFATc1) antibody was acquired from BD biosciences (San Jose, CA, USA). The enhanced chemiluminescence (ECL) Western blotting detection system was purchased from Advansta Inc. (San Jose, CA, USA).
2.2. Isolation and Identification of Amygdalin from the Seeds of Prunus armeniaca L.
The dried seeds of
P. armeniaca L. (1.0 kg) were extracted three times with EtOH (1 L × 3 h) under reflux. After concentrating under reduced pressure, the EtOH extract (89.7 g) was suspended in distilled water and partitioned with n-hexane, CH
2Cl
2, EtOAc, and n-butanol to yield an n-hexane extract (50 g), a CH
2Cl
2 extract (8.8 g), an EtOAc extract (9.6 g), an n-butanol extract (6.5 g), and a water layer. The n-butanol extract was chromatographed with column chromatography (CC) on silica gel using stepwise eluent of CH
2Cl
2–acetone (gradient 10:1–1:1,
v/
v) and then CH
2Cl
2–MeOH–H
2O (gradient 6:1:0.1–1:1:0,
v/
v) to obtain 6 fractions (A–F). Compound 1 (35 mg) was isolated from fraction F (177.5 mg) by reserved phase C18 (RP-18) CC using MeOH–H
2O (3:1,
v/
v) as an eluent, followed by preparative high-performance liquid chromatography (HPLC) with an isocratic mixture solvent, 60% MeOH in H
2O, at 5 mL/min for 60 min. The nuclear magnetic resonance (NMR) spectra of compound 1 (
1H and
13C) were conducted using a 500 MHz NMR spectrometer (JEOL, JNM-ECA 500) with tetramethylsilane (TMS) as an internal standard. NMR solvent DMSO-
d6 was purchased from Sigma-Aldrich (MO, USA). The structure of compound 1 was confirmed as amygdalin using spectroscopic analysis of the NMR data and by comparison with other studies [
24,
25] (
Table S1, Figures S1 and S2).
2.3. Molecular Docking Simulation
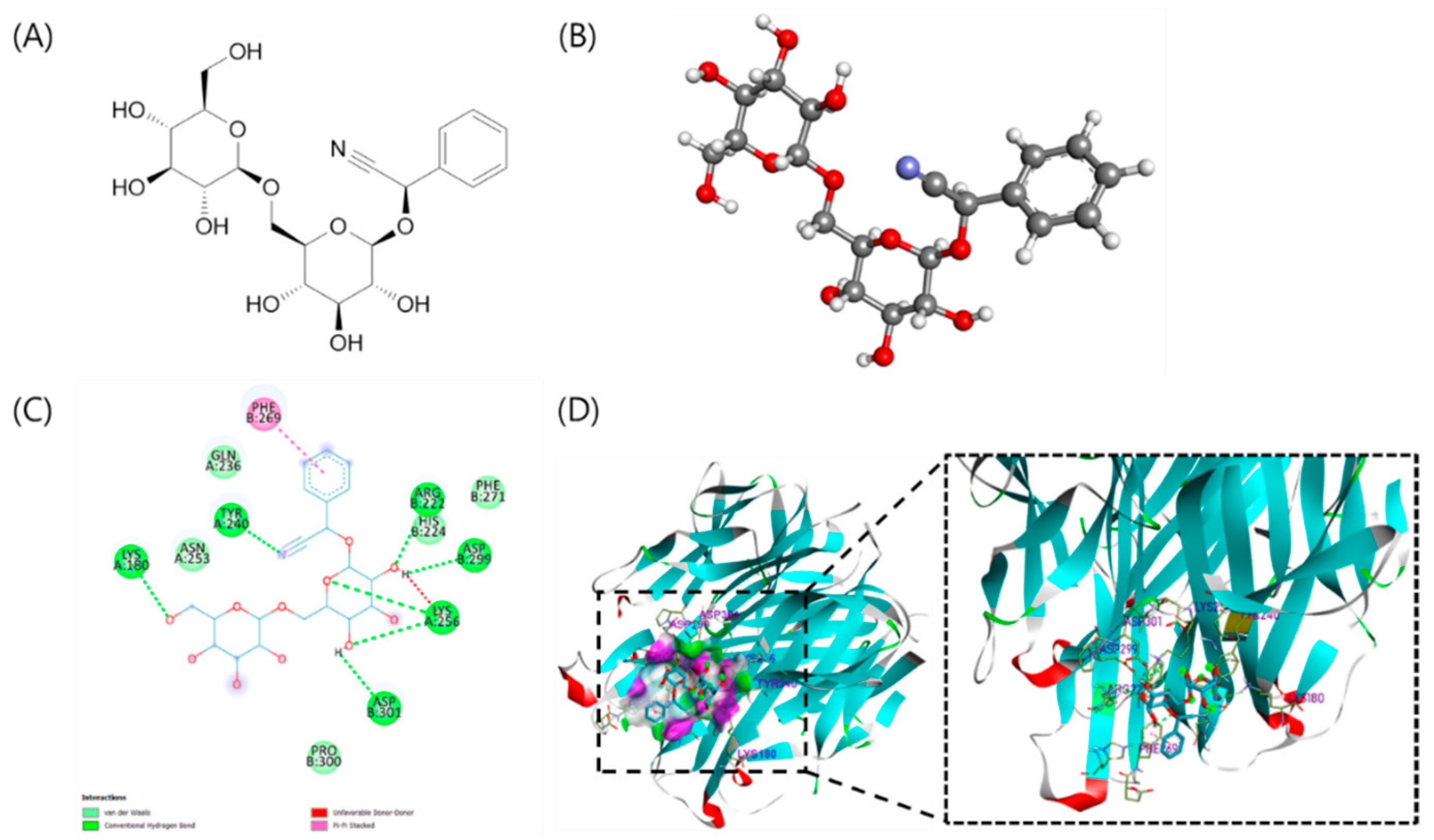
The crystal structure of the extracellular domain of mouse RANK ligand with the resolution of 2.20 Å was downloaded from the RSCB Protein Data Bank website (PDB ID: 1IQA) [
26]. The crystal structure was prepared for docking using Discovery Studio v21.1 (Accelrys, Inc., San Diego, CA, USA). The binding of RANKL and water molecules were removed from the structure for docking simulation using AutoDockTools (ADT). AutoDock Vina 1.1.2 was utilized to perform molecular docking to understand the interaction and binding of AD with RANKL. For the docking procedure, the 3D structure of AD was constructed using ChemDraw 20.1 and Chem3D 20.1 (PerkinElmer, INC., MA, USA) for 2D and 3D conformations, respectively. Automated docking simulation was performed using ADT to assess the appropriate binding orientations and conformations of the ligand molecules with the protein inhibitor. The Lamarckian genetic algorithm (LGA) method implemented in AutoDock Vina was employed. For docking calculations, Gasteiger charges were added by default, the rotatable bonds were set by the AutoDock tools, and all torsions were allowed to rotate. The binding effect of RANKL residues, following the binding affinity score and the root-mean-square deviation (RMSD), was considered as the best molecular interaction. Discovery Studio v21.1 was used for graphic display and analysis of the interactions between AD and the target protein. Various interactions, such as hydrogen bonds, van der Waals interactions, π–π bonds, and the interaction distance between amino acids and the active sites of compounds, were plotted. Van der Waals interactions were detected between AD and the bonding point by hydrophilic and hydrophobic contact.
2.4. Cell Culture and Osteoclast Differentiation of RAW 264.7 Cells
RAW 264.7 murine macrophage cells were acquired from the American Type Culture Collection (ATCC, Rockville, MD, USA). RAW 264.7 cells were seeded at 5 × 103 cells/well in 24-well plates and grown in Dulbecco’s modified Eagle’s medium (DMEM) high glucose supplemented with 10% fetal bovine serum (FBS), 100 U/mL of penicillin, and streptomycin. After then, RAW 264.7 cells were incubated for 24 h at 37°C in a humidified atmosphere containing 5% CO2. Then, each well was changed to minimum essential medium alpha (α-MEM) containing 10% FBS, 100U/mL penicillin, and streptomycin. After being treated or untreated with RANKL (50 ng/mL), cells were processed with AD at different concentrations (5, 10, 20, 40 µM) for 5 days. The medium was refreshed every day. Osteoclast differentiation was identified by TRAP staining, activity, and other experimental methods.
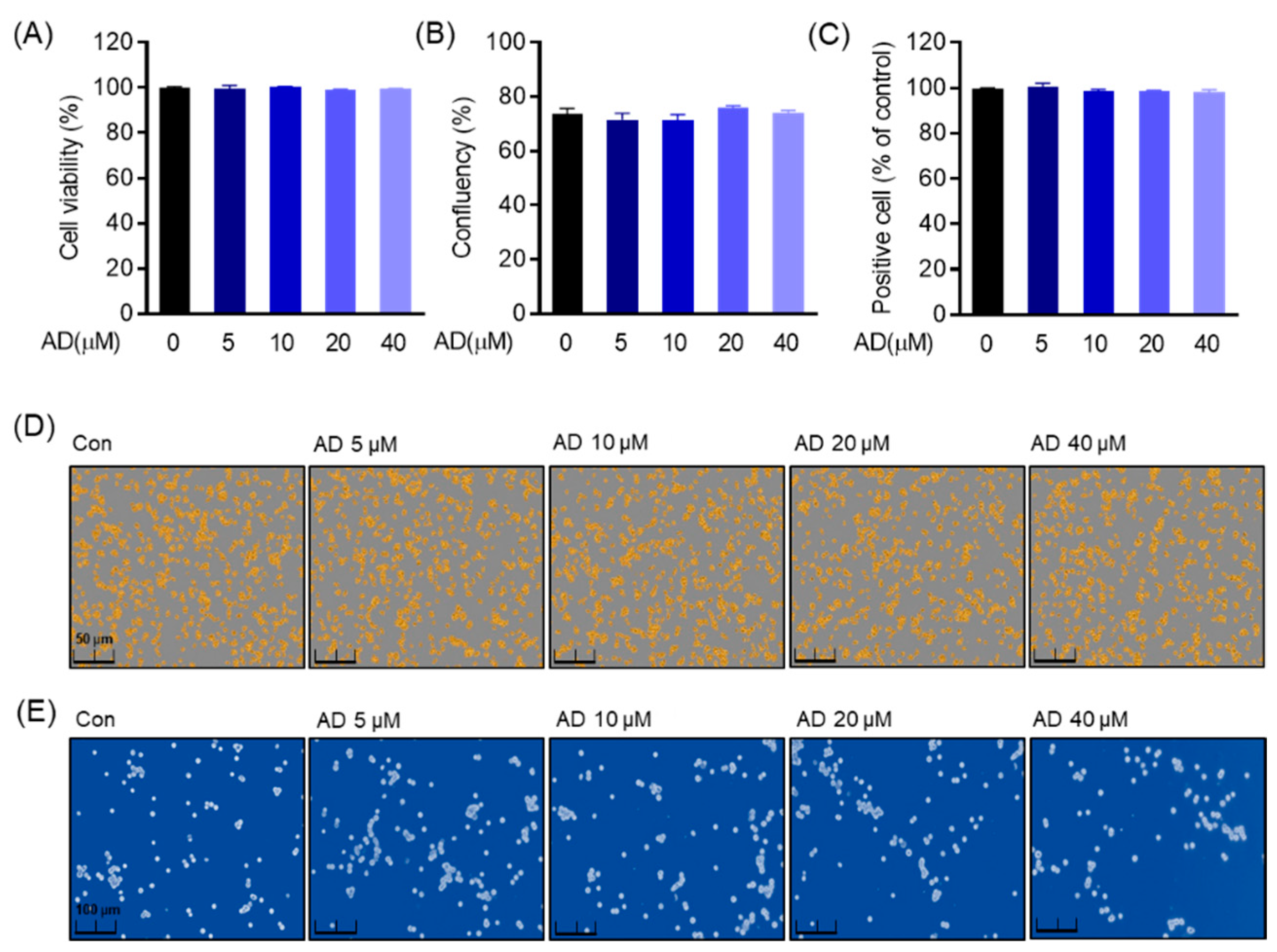
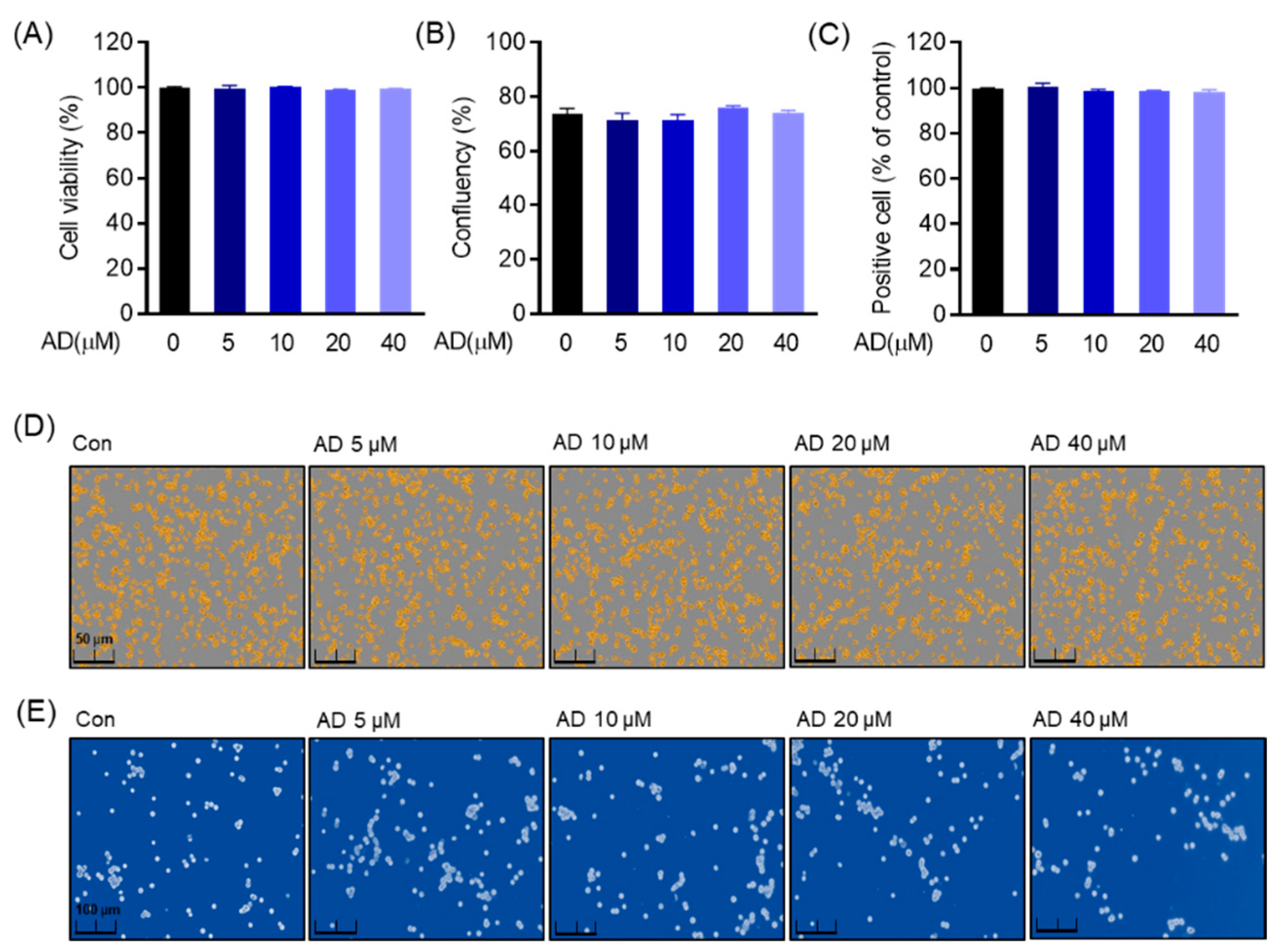
2.5. Cell Viability
The cytotoxic effect of AD on RAW 264.7 cells was detected by an MTT assay according to the following procedure. RAW 264.7 (5 × 103 cells/well) cells were seeded in 96-well plates and maintained at 37 °C in a humidified 5% CO2 incubator for 24 h. Cells were treated in the presence or absence of different concentrations of AD for 5 days. Then, 4,5-dimethylthiazol-2-thiazolyl)-5-diphenyltetrazolium bromide (MTT) (5 mg/mL) was added to each well. Next, cells were incubated for 4 h before the supernatant was replaced with dimethyl sulfoxide (DMSO) to dissolve formazan crystals. Cell viability was exhibited at the 540 nm absorbance in a microplate reader (TECAN infinity pro 2000, Männedorf, Switzerland). The results shown are representative of three independent experiments. After trypan blue staining was also treated in the presence or absence of different concentrations of AD for 5 days, RAW 264.7 cells were stained with 0.4% trypan blue and photographed under an optical microscope (CKX53, Olympus, Tokyo, Japan). The amount of positive cells was estimated by counting stained and unstained cells.
2.6. Cell Migration
RAW 264.7 cells were seeded at the density of 1 × 103 cells/well in 6-well plates. Then, cells were incubated for 24 h followed by a constant scratching before being treated or untreated with RANKL (50 ng/mL) and the indicated AD concentration. The amount of migration of yellow marked cells in the indicated red line was measured on the first day and fifth day after treatment using Incucyte® Live Cell analysis systems. The only RANKL-treated group was used as a normalized percentage.
2.7. TRAP Staining and Activity
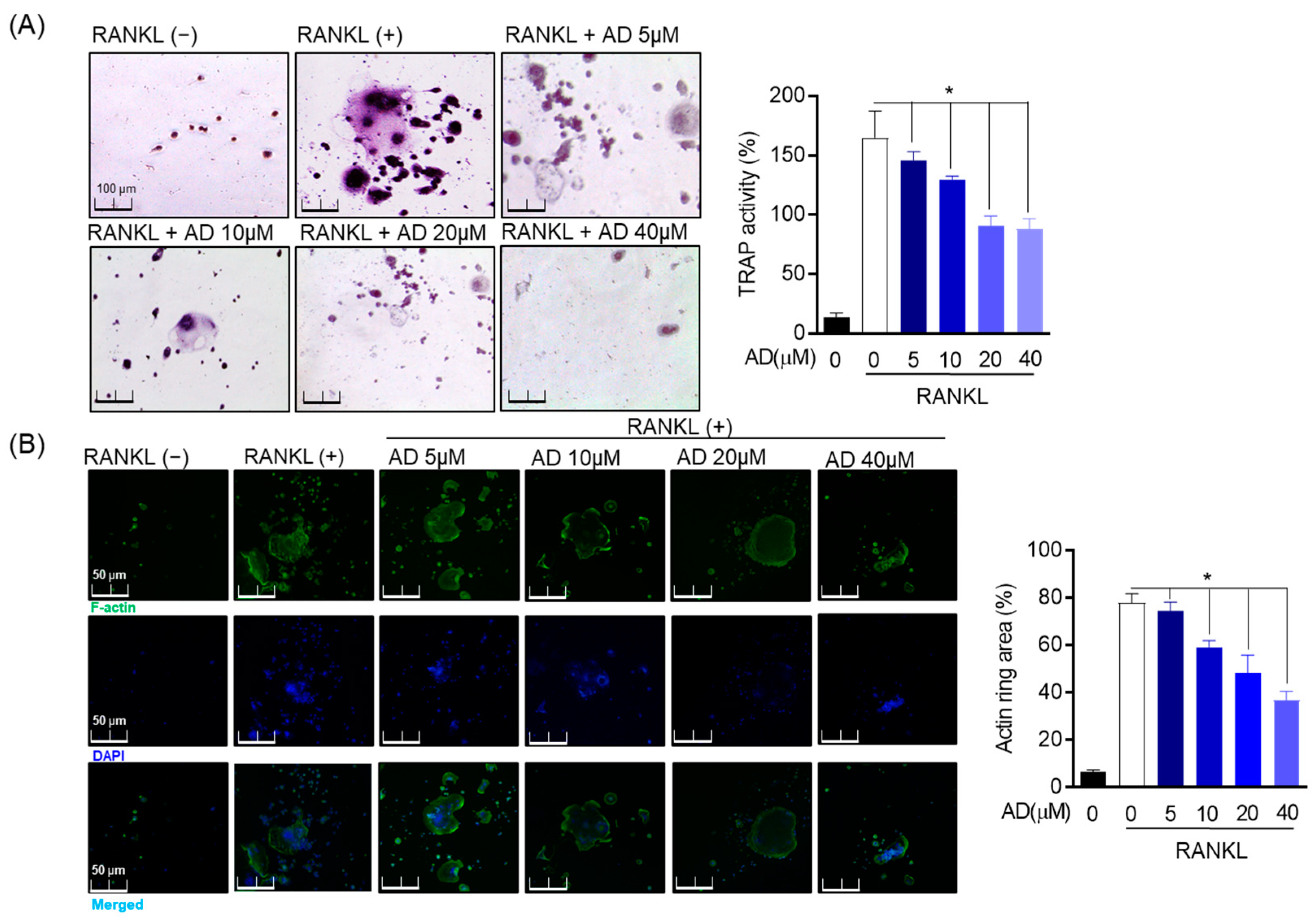
RAW 264.7 cells (1 × 103 cells/well) were cultured in 24-well culture plates in DMEM containing 10% FBS, 100 U/mL of penicillin, and streptomycin. After being incubated for 24 h, the medium was changed to α-MEM containing RANKL (50 ng/mL), and cells were treated with AD at different concentrations. The medium was refreshed every day. After 5 days of incubation, the medium was removed and washed twice with phosphate-buffered saline (PBS). Then, cells were fixed with 4% formaldehyde for 15 min and washed with PBS. Osteoclast differentiation was measured by the Acid Phosphatase Assay Lit (Cat. No. CS 0740) and acid phosphatase leukocyte (Cat. No. 387A-1KT) from Sigma-Aldrich Fine Chemicals (Saint Louis, MO, USA) according to the instruction of the manufacturer. Briefly, fixed cells were reacted with a leukocyte acid phosphatase kit at 37 °C, 5% CO2 for 1 h, and then the reaction was conducted in a shaded state. Next, cells were washed three times with PBS and TRAP-positive multinuclear cells, which have three or more nuclei, according to optical microscopic observations.
2.8. Actin Ring and DAPI Staining
Alexa 488 Phalloidin (Invitrogen, Carlsbad, CA, USA) was used to measure the degree of differentiation of osteoclasts by staining the ring part. RAW 264.7 cells were maintained in the medium including processed or unprocessed AD for 5 days. Cells were fixed in 4% formaldehyde for 15 min followed by a treatment with 0.5% Triton X-100 solution to permeabilize the cells. Afterward, the experiment was conducted in the dark by staining cells with Alexa 488 for 1 h, then contrasted staining for nuclei with DAPI for 30 min followed by cold PBS washing. Actin rings and DAPI staining in mature osteoclasts were observed with a fluorescence microscope (Nikon Co., Tokyo, Japan).
2.9. Bone Resorption Assay
RAW 264.7 cells (1 × 103 cells/well) were cultured on a Corning osteo assay surface well (Corning, NY, USA). After 24 h of incubation, cells were refreshed with α-MEM medium containing 5% FBS and treated or untreated with RANKL (50 ng/mL) and the indicated AD. The cells were maintained for 7 days at 37 °C in a humidified incubator containing 5% CO2. The medium was changed every day. Subsequently, the medium was removed, cells were washed twice with PBS. Cells were detached by 5% sodium hypochlorite for 5 min, washed with PBS, and dried. The surface of each well was visualized using a microscope (Olympus, Tokyo, Japan) and measured with an ImageJ program.
2.10. Measurement of Intracellular Reactive Oxygen Species
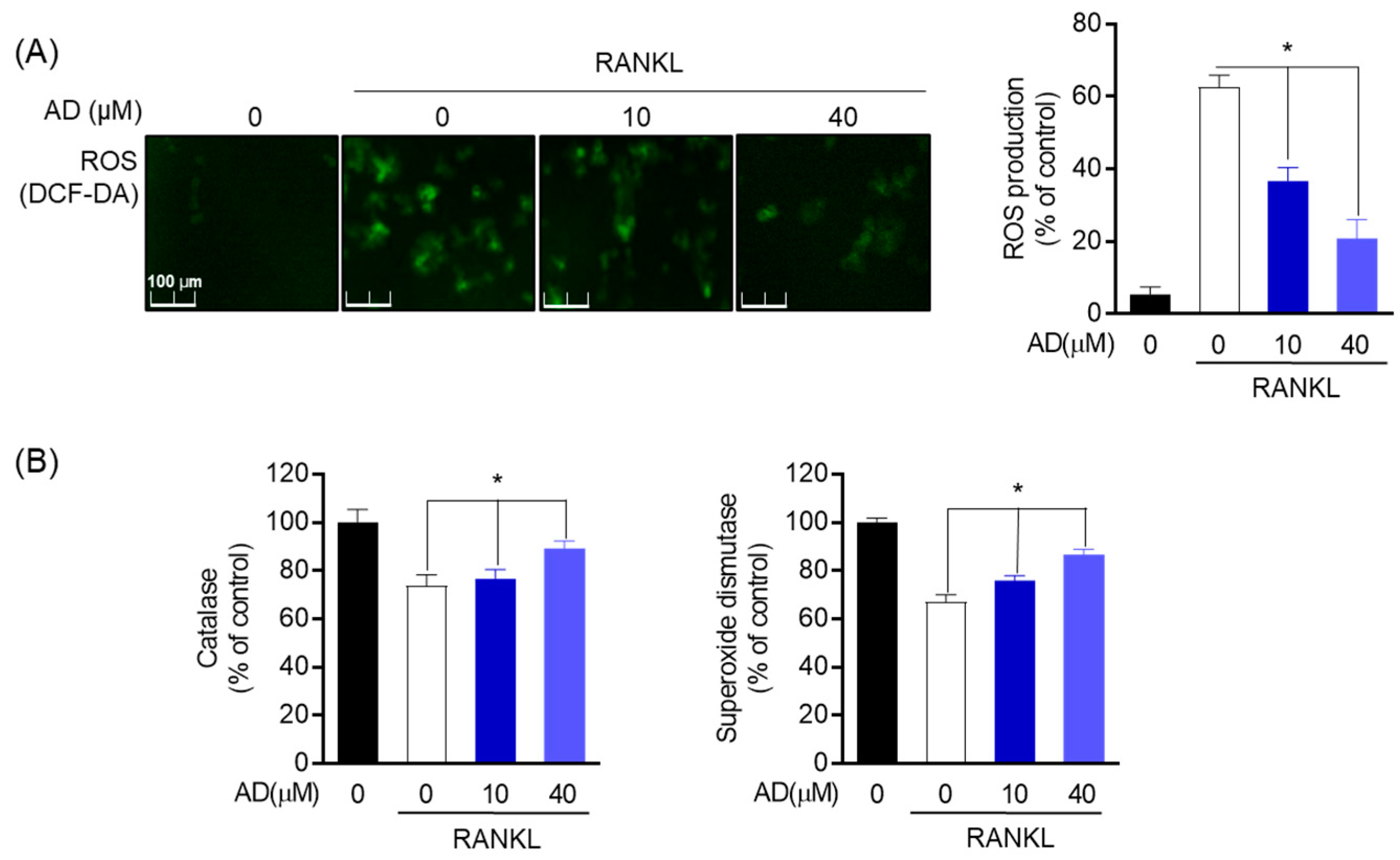
RAW 264.7 (5 × 103 cells/well) was dispended in 24-well plates for 24 h. After treatment with RANKL (50 ng/mL), cells were treated with AD at different concentrations and maintained with α-MEM supplemented with 10% FBS, 100U/mL penicillin and streptomycin for 24 h. 2′, 7′-dichlorodihydrofluorescein diacetate (DCF-DA) was utilized to study the production of ROS. After incubation for 20 min at 37 °C in the dark, cells were washed with PBS and fixed with 4% paraformaldehyde (pH 7.4) for 20 min. ROS was detected with a fluorescent Olympus IX microscope 71-F3 2PH (Tokyo, Japan).
2.11. Measurement of Antioxidant Enzyme Activity
Antioxidant enzymes were measured using a Superoxide Dismutase Assay Kit (Cayman chemical, Ann arbor, MI, USA) and a Catalase Assay Kit (Cayman chemical, Ann arbor, MI, USA), respectively. RAW 264.7 cells treated with AD and RANKL were recovered using extraction buffer (20 mM HEPES buffer, pH 7.2, 1 mM EGTA, 210 mM mannitol, 70 mM sucrose) and centrifuged, while SOD was measured in a 450 nm ELISA plate reader from the recovered supernatant. Then, CAT was measured at 540 nm with an ELISA plate reader from the supernatant recovered using the extraction buffer (50 mM potassium phosphate, pH 7.0, 1 mM EDTA) according to the manufacturer’s instructions.
2.12. Real-Time Quantitative PCR
Real-time PCR was used to detect the expression of
dcstamp,
acp5,
atp6d0v2, and
ctsk. The cell lysate was placed in a TRIZOL reagent to extract the total RNA. NanoDrop (Thermo Scientific, Waltham, MA, USA) and TOPscript™ RT DryMIX (dT18 plus) were used to analyze the concentration of mRNA and synthesize cDNA, respectively. Real-time PCR reactions operated in a LightCycler 480 (Roche, Basel, Switzerland) instrument using TB Green
® Premix Ex Taq™ II (Tli RNaseH Plus).
gapdh was used as a housekeeping gene, and the mRNA level of each gene was normalized compared to
gapdh. The calculation equation for gene expression is as follows: 2−ΔΔCT, where ΔΔCT = (CT
target−CT
gapdh) at time x−(CT
target−CT
gapdh) at time 0, time x represents any time point, and time 0 represents the 1 X expression of the gene in the untreated group normalized to
gapdh. The experiment was conducted three times separately. The primers are displayed in
Table S2.
2.13. Western Blot Analysis
RAW 264.7 cells were pretreated with or without AD (5–40 µM) and stimulated with RANKL (50 ng/mL) to detect the activation of MAPK. The total lysate was prepared using RIPA buffer containing protease inhibitors for 30 min on ice followed by centrifugation at 14,000 rpm for 20 min at 4 °C. Then, the Bradford assay was used to measure the protein concentration according to the instruction of the manufacturer. An equal amount of protein for each sample was separated in 12% SDS-PAGE gel. After electrophoresis, separated proteins were transferred onto the PVDF membrane (Bio-Rad, Hercules, CA, USA). The membrane was blocked in 5% skim milk for 1 h at room temperature. The membranes were incubated with anti-BIP, anti-phospho-eIF2α, and anti-eIF2α to detect the expression of ER stress; anti-NFATc1 and anti-c-fos to detect osteoclast transcription factors; and anti-phospho-ERK, anti-ERK, anti-phospho-JNK, JNK, anti-phospho-p38, and anti-p38 to detect expression of MAPK. Furthermore, β-actin was used as the housekeeping gene. Then, the membranes were incubated overnight at 4oC before being washed in TBS-T followed by the incubation with secondary antibodies. The antibody-bounded proteins were visualized with ECL Western blotting detection reagents (Thermo Fisher Scientific, Waltham, MA, USA) using ImageQuant LAS 4000 (GE Healthcare, Chicago, IL, USA). ImageJ software was employed to quantify the observed band and normalized with control.
2.14. Statistics
The Sigma Plot 12.1 Student’s t-test was used to calculate statistical significance. The data were expressed as means ± standard deviation (SD) for at least three independent experiments, with p < 0.05 being considered as statistically significant.
4. Discussion
Under physiological conditions, through a balance between bone formation by osteoblast and bone resorption by osteoclast, the maintenance of bone mass is achieved [
30]. Osteoclast, the principle of bone resorption in the body, plays an important role in skeletal development and maintenance regulates osteoblast differentiation, promotes the mobilization of hematopoietic stem cell from the bone marrow to the bloodstream, and is involved in the response of the immune system. In some cases, this balance is disrupted, and the activity of osteoclast is over-expressed or abnormal, leading to the activation of numerous bone metabolic disorders and localized bone diseases [
30,
31,
32,
33]. Thus, understanding the mechanism underlying osteoclast and bone-related disorder is crucial for the development of natural products as a treatment for osteoclastogenesis and bone diseases.
In this study, since the in silico study of AD and osteoclast is currently unknown, the computational screening of AD and its properties related to osteoclast differentiation was first assessed. The AA” loop in protein RANKL was reported to have an important role in osteoclast differentiation [
26,
29]. By molecular docking, the result predicted that AD effectively binds to the active site of RANKL, which has been known as the key regulator of osteoclast precursors. Additionally, in 2021, Abbas et al. reported that resveratrol, a common secondary metabolite found in red grapes, red wine, and peanuts, was exhibited as a potential compound for drug preparation against bone loss by using in vitro and in silico methods. By using molecular docking simulation between resveratrol and eight RANKL receptors (PDB ID: 1JTZ, 1S55, 3ME2, 3QBQ, 4E4D, 4GIQ, 1IQA, and 1TJZ), resveratrol showed the high binding affinities from −7.6 to −6.7 kcal/mol [
34]. Besides, in 2019, El-Baz et al. revealed that astaxanthin-rich fraction from
Heamatococcus pluvialis shows benefits in the control of age-related osteoporosis, not only through preserving bone mass and serum calcium/phosphorus level but also through an increase in the bone mineralization rate. Especially, all-trans astaxanthin, the major carotenoid found in
H. pluvialis, possessed a high binding affinity towards the OPG-RANKL complex protein with the free energy of binding −6.3 kcal/mol [
35]. Our study revealed that AD showed high binding affinities with 1IQA, the crystal structure of the extracellular domain of mouse RANK ligand, of −6.4 kcal/mol. These results suggested that AD may be a potential compound for the inhibition of RANKL.
Then, the in vitro experiment on RAW264.7 cell model was carried out to study the effect of AD on osteoclast differentiation. RANKL, a central positive regulator of osteoclast produced by osteoblast-lineage cells, binds to its receptor RANK on the surface of myeloid cells, stimulating their differentiation into osteoclasts which results in the activation of osteoclast formation along with the rapid recruitment of multiple intracellular signaling molecules [
32,
33,
36,
37]. TRAP is the mature osteoclast marker [
30]. The data revealed that RANKL-induced cells were able to form TRAP-positive osteoclast and that this process was inhibited by AD at this indicated experimental concentration. Osteoclast differentiation is regulated by various upstream signaling of MAPK, including JNK, ERK, and p38 [
38]. RANKL-induced bone resorption is critically regulated through MAPK induced c-Fos expression [
36]. NFATc1 is the crucial downstream signaling event in RANKL-mediated ROS signaling, and its expression is regulated by c-Fos/c-Jun. Thus, increased NFATc1 induces the formation of TRAP-positive osteoclast cells [
36]. In this study, AD suppresses the phosphorylation of MAPK activation and osteoclast differentiate downstream signal, including c-Fos and NFATc1. RANKL-induced osteoclast differentiation through oxidative stress and ER stress in the regulation of osteoclast differentiation is reported, relating to the inflammatory process [
36,
39]. ROS is known as a pivotal regulator of ER function along with ER stress, and increased ROS production occurs concurrently [
6,
40]. The results suggested that the inhibitory effect of AD was exhibited by suppressed ROS accumulation by DCF-DA staining and the activation of antioxidant enzymes, such as CAT and SOD. In addition, in the presence of AD, the expression of cytosolic protein, including BIP and eIF2α, was significantly inhibited.
In conclusion, these data demonstrated that AD inhibited osteoclast by RANKL-induced osteoclast in the RAW264.7 cell model. The treatment of AD not only suppresses the structure and function of osteoclast but also the expression osteoclast-specific genes and transcription factors, thereby positively influencing the treatment of osteoporosis for further study. Additionally, the generation of ROS and ER stress was strongly inhibited by AD. Finally, AD downregulates the activation of MAPK which is an old cell signaling pathway related to osteoclastogenesis. Our study suggests that amygdalin (AD), a secondary compound isolated from seeds of P. armeniaca L., has the potential to become a natural-derived treatment for osteoclastogenesis and bone disorder diseases.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}