
Irisin Association with Ki-67, MCM3 and MT-I/II in Squamous Cell Carcinomas of the Larynx

Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients Cohort
2.2. Cell Line Culture
2.3. Tissues Microarray (TMA) Preparation
2.4. Immunohistochemistry (IHC)
2.5. Evaluation of Immunohistochemistry (IHC)
2.6. Immunofluorescence (IF)
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
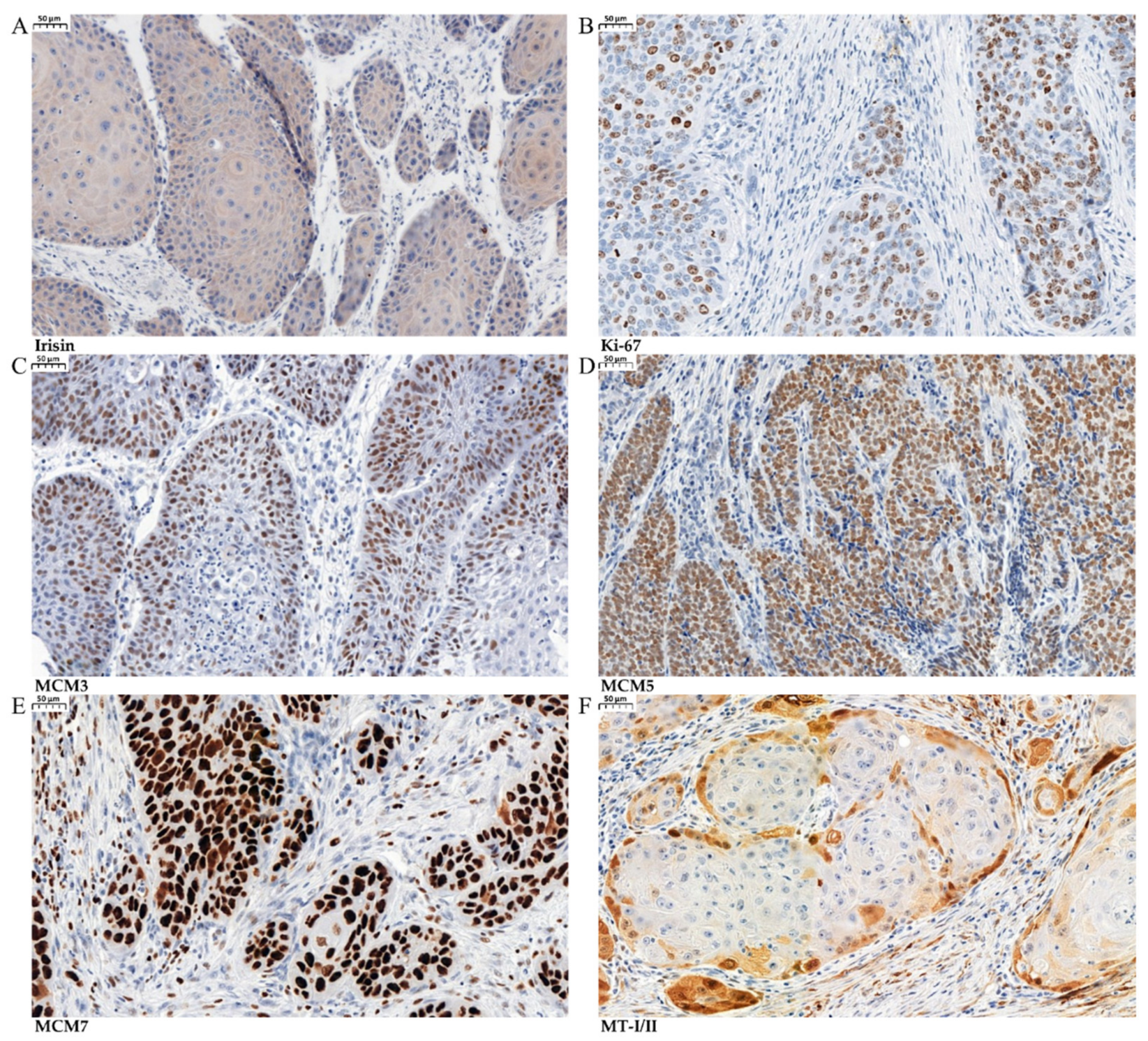
3.1. Immunohistochemical (IHC) Detection of Irisin Expression in Tissue Microarrays (TMAs)
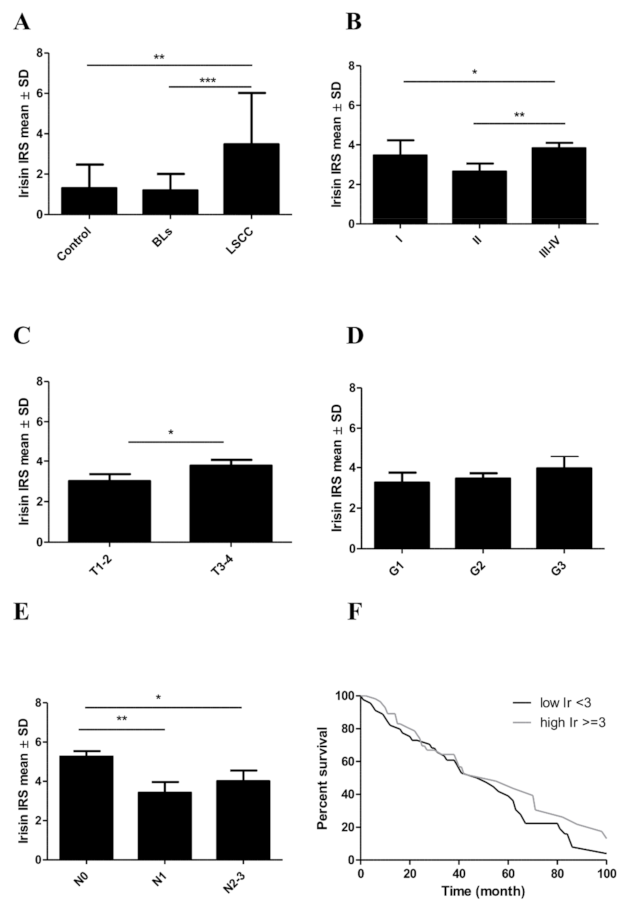
3.2. Associations between Irisin Expression in Cancer Cells and Clinicopathological Parameters
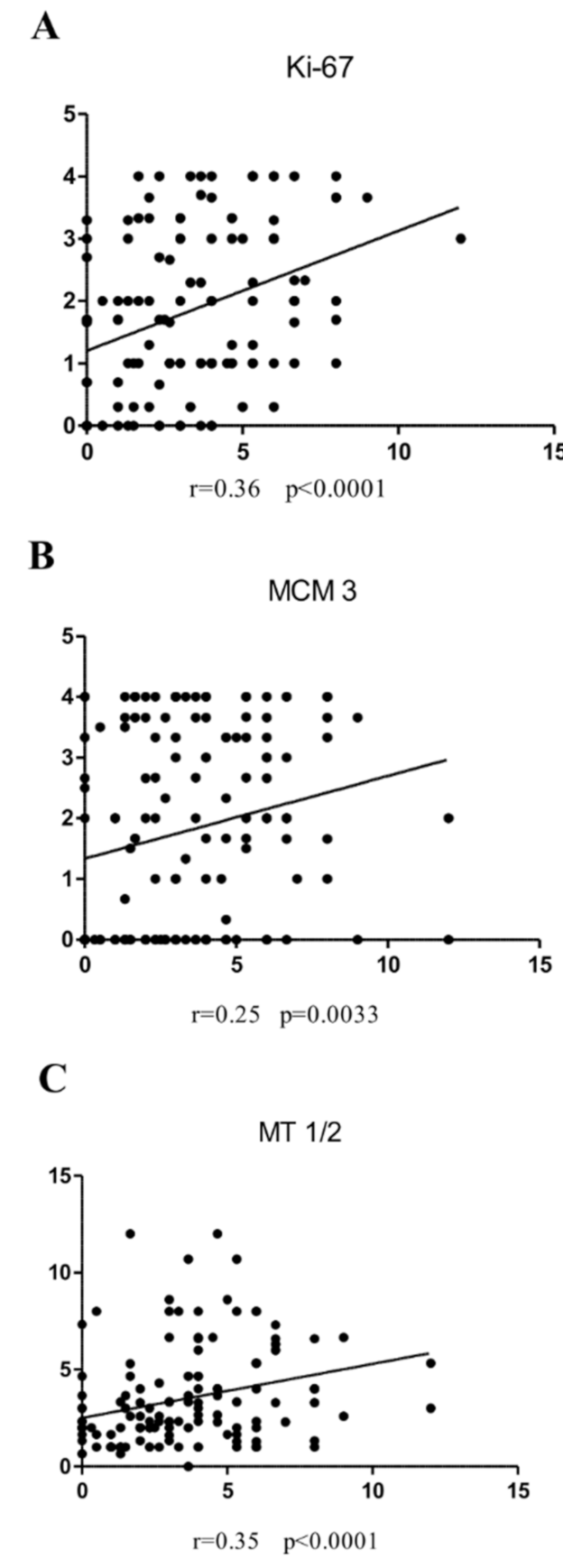
3.3. Associations between Irisin and Cancer Cell Proliferation
3.4. Irisin Expression Levels in Cancer Cell Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaczmarczyk, D.; Bruzgielewicz, A.; Osuch-Wójcikiewicz, E. Histopatologia i Zmiany Przedrakowe w Raku Krtani. Pol. Przegląd Otorynolaryngologiczny 2014, 3, 132–139. [Google Scholar] [CrossRef]
- Krajowy Rejestr Nowotwórów. Available online: http://onkologia.org.pl/nowotwory-zlosliwe-krtani-c32/ (accessed on 23 November 2021).
- Kruk-Zagajewska, A.; Werzbicka, M.; Leszczyńska, M.; Kordylewska, M.; Szyfter, W. Diagnosis and Treatment of Larynx Cancer. Adv. Head Neck Surg. 2006, 5, 5–15. [Google Scholar]
- Badwal, J.S. Total Laryngectomy for Treatment of T4 Laryngeal Cancer: Trends and Survival Outcomes. Polish J. Surg. 2019. [Google Scholar] [CrossRef]
- Souza, F.G.R.; Santos, I.C.; Bergmann, A.; Thuler, L.C.S.; Freitas, A.S.; Freitas, E.Q.; Dias, F.L. Quality of Life after Total Laryngectomy: Impact of Different Vocal Rehabilitation Methods in a Middle Income Country. Health Qual. Life Outcomes 2020, 18, 92. [Google Scholar] [CrossRef]
- van Sluis, K.E.; Kornman, A.F.; van der Molen, L.; van den Brekel, M.W.M.; Yaron, G. Women’s Perspective on Life after Total Laryngectomy: A Qualitative Study. Int. J. Lang. Commun. Disord. 2020. [Google Scholar] [CrossRef] [Green Version]
- Ugur, K.; Aydin, S.; Kuloglu, T.; Artas, G.; Kocdor, M.A.; Sahin, İ.; Yardim, M.; Hanifi Ozercan, İ. Comparison of Irisin Hormone Expression between Thyroid Cancer Tissues and Oncocytic Variant Cells. Cancer Manag. Res. 2019, 11, 2595–2603. [Google Scholar] [CrossRef]
- Nowinska, K.; Jablonska, K.; Pawelczyk, K.; Piotrowska, A.; Partynska, A.; Gomulkiewicz, A.; Ciesielska, U.; Katnik, E.; Grzegrzolka, J.; Glatzel-Plucinska, N.; et al. Expression of Irisin/FNDC5 in Cancer Cells and Stromal Fibroblasts of Non-Small Cell Lung Cancer. Cancers 2019, 11, 1538. [Google Scholar] [CrossRef] [Green Version]
- Shao, L.; Li, H.; Chen, J.; Song, H.; Zhang, Y.; Wu, F.; Wang, W.; Zhang, W.; Wang, F.; Li, H.; et al. Irisin Suppresses the Migration, Proliferation, and Invasion of Lung Cancer Cells via Inhibition of Epithelial-to-Mesenchymal Transition. Biochem. Biophys. Res. Commun. 2017, 485, 598–605. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-Dependent Myokine That Drives Brown-Fat-like Development of White Fat and Thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Maak, S.; Norheim, F.; Drevon, C.A.; Erickson, H.P. Progress and Challenges in the Biology of FNDC5 and Irisin. Endocr. Rev. 2021, 42, 436–456. [Google Scholar] [CrossRef] [PubMed]
- Pinkowska, A.; Podhorska-Okołów, M.; Dzięgiel, P.; Nowińska, K. The Role of Irisin in Cancer Disease. Cells 2021, 10, 1479. [Google Scholar] [CrossRef]
- Nie, Y.; Liu, D. N-Glycosylation Is Required for FDNC5 Stabilization and Irisin Secretion. Biochem. J. 2017, 474, 3167–3177. [Google Scholar] [CrossRef]
- Ząbczyńska, M.; Pocheć, E. The Role of Protein Glycosylation in Immune System. Postepy Biochem. 2015, 61, 129–137. [Google Scholar] [PubMed]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic Peroxisome Proliferator-Activated Receptor γ (PPARγ) Activation of Epididymally Derived White Adipocyte Cultures Reveals a Population of Thermogenically Competent, UCP1-Containing Adipocytes Molecularly Distinct from Classic Brown Adipocytes. J. Biol. Chem. 2010, 285, 7153–7164. [Google Scholar] [CrossRef] [Green Version]
- Arhire, L.I.; Mihalache, L.; Covasa, M. Irisin: A Hope in Understanding and Managing Obesity and Metabolic Syndrome. Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Aydin, S.; Kuloglu, T.; Aydin, S.; Kalayci, M.; Yilmaz, M.; Cakmak, T.; Albayrak, S.; Gungor, S.; Colakoglu, N.; Ozercan, İ.H. A Comprehensive Immunohistochemical Examination of the Distribution of the Fat-Burning Protein Irisin in Biological Tissues. Peptides 2014, 61, 130–136. [Google Scholar] [CrossRef]
- Estell, E.G.; Le, P.T.; Vegting, Y.; Kim, H.; Wrann, C.; Bouxsein, M.L.; Nagano, K.; Baron, R.; Spiegelman, B.M.; Rosen, C.J. Irisin Directly Stimulates Osteoclastogenesis and Bone Resorption in Vitro and in Vivo. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Wrann, C.D.; Jedrychowski, M.; Vidoni, S.; Kitase, Y.; Nagano, K.; Zhou, C.; Chou, J.; Parkman, V.-J.A.; Novick, S.J.; et al. Irisin Mediates Effects on Bone and Fat via AV Integrin Receptors. Cells 2018, 175, 1756–1768. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.; Zhang, J.; Ren, Y.; Du, Z.; Li, T.; Wang, T.; Zhang, L.; Wang, M.; Wu, Z.; Lv, Y.; et al. Irisin Reverses Intestinal Epithelial Barrier Dysfunction during Intestinal Injury via Binding to the Integrin AVβ5 Receptor. J. Cell. Mol. Med. 2020, 24, 996–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuloglu, T.; Celik, O.; Aydin, S.; Hanifi Ozercan, I.; Acet, M.; Aydin, Y.; Artas, G.; Turk, A.; Yardim, M.; Ozan, G.; et al. Irisin Immunostaining Characteristics of Breast and Ovarian Cancer Cells. Cell. Mol. Biol 2016, 62, 40–44. [Google Scholar] [CrossRef]
- Tekin, S.; Erden, Y.; Sandal, S.; Yilmaz, B. Is Irisin an Anticarcinogenic Peptide? Med. Sci. 2015, 4, 2172–2180. [Google Scholar] [CrossRef]
- Aydin, S.; Kuloglu, T.; Ozercan, M.; Albayrak, S.; Aydin, S.; Bakal, U.; Yilmaz, M.; Kalayci, M.; Yardim, M.; Sarac, M.; et al. Irisin Immunohistochemistry in Gastrointestinal System Cancers. Biotech. Histochem. 2016, 91, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Kong, G.; Jiang, Y.; Sun, X.; Cao, Z.; Zhang, G.; Zhao, Z.; Zhao, Y.; Yu, Q.; Cheng, G. Irisin Reverses the IL-6 Induced Epithelial-Mesenchymal Transition in Osteosarcoma Cell Migration and Invasion through the STAT3/Snail Signaling Pathway. Oncol. Rep. 2017, 38, 2647–2656. [Google Scholar] [CrossRef] [Green Version]
- Catalano, V.; Turdo, A.; Di Franco, S.; Dieli, F.; Todaro, M.; Stassi, G. Tumor and Its Microenvironment: A Synergistic Interplay. Semin. Cancer Biol. 2013, 23, 522–532. [Google Scholar] [CrossRef]
- Gannon, N.P.; Vaughan, R.A.; Garcia-Smith, R.; Bisoffi, M.; Trujillo, K.A. Effects of the Exercise-Inducible Myokine Irisin on Malignant and Non-Malignant Breast Epithelial Cell Behavior in Vitro. Int. J. Cancer 2015, 136, E197–E202. [Google Scholar] [CrossRef]
- Liu, J.; Song, N.; Huang, Y.; Chen, Y. Irisin Inhibits Pancreatic Cancer Cell Growth via the AMPK-MTOR Pathway. Sci. Rep. 2018, 8, 15247. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.-S.; Mantzoros, C.S. Regulation of Cell Proliferation and Malignant Potential by Irisin in Endometrial, Colon, Thyroid and Esophageal Cancer Cell Lines. Metabolism 2014, 63, 188–193. [Google Scholar] [CrossRef]
- Juríková, M.; Danihel, Ľ.; Polák, Š.; Varga, I. Ki67, PCNA, and MCM Proteins: Markers of Proliferation in the Diagnosis of Breast Cancer. Acta Histochem. 2016, 118, 544–552. [Google Scholar] [CrossRef]
- Vukelic, J.; Dobrila-Dintinjana, R.; Dekanic, A.; Marijic, B.; Cubranic, A.; Braut, T. The Relevance of Assessing the Cell Proliferation Factor Ki-67 in Squamous Cell Carcinoma of the Larynx. Biomed Res. Int. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, U.; Zatonski, T.; Nowinska, K.; Ratajczak-Wielgomas, K.; Grzegrzolka, J.; Piotrowska, A.; Olbromski, M.; Pula, B.; Podhorska-Okolow, M.; Dziegiel, P. Expression of Cell Cycle-Related Proteins P16, P27 and Ki-67 Proliferating Marker in Laryngeal Squamous Cell Carcinomas and in Laryngeal Papillomas. Anticancer Res. 2017, 37, 2407–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowinska, K.; Dziegiel, P. The Role of MCM Proteins in Cell Proliferation and Tumorigenesis. Postep. Hig. Med. Dosw. 2010, 64, 627–635. [Google Scholar]
- Nowinska, K.; Ciesielska, U.; Piotrowska, A.; Jablonska, K.; Partynska, A.; Paprocka, M.; Zatonski, T.; Podhorska-Okolow, M.; Dziegiel, P. MCM5 Expression Is Associated With the Grade of Malignancy and Ki-67 Antigen in LSCC. Anticancer Res. 2019, 39, 2325–2335. [Google Scholar] [CrossRef] [Green Version]
- Nowinska, K.; Chmielewska, M.; Piotrowska, A.; Pula, B.; Pastuszewski, W.; Krecicki, T.; Podhorska-Okolow, M.; Zabel, M.; Dziegiel, P. Correlation between Levels of Expression of Minichromosome Maintenance Proteins, Ki-67 Proliferation Antigen and Metallothionein I/II in Laryngeal Squamous Cell Cancer. Int. J. Oncol. 2016, 48, 635–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywoszyńska, K.; Kozłowski, H. Metallothioneins and Polythiol Motifs: Interactions with Metal Ions. Wiadomości Chem. 2018, 72, 383–395. [Google Scholar]
- Bizoń, A.; Jędryczko, K.; Milnerowicz, H. Rola Metalotioneiny w Procesie Nowotworzenia Oraz w Leczeniu Chorób NowotworowychThe Role of Metallothionein in Oncogenesis and Cancer Treatment. Postep. Hig. Med. Dosw. 2017, 71, 98–109. [Google Scholar] [CrossRef]
- Werynska, B.; Pula, B.; Kobierzycki, C.; Dziegiel, P.; Podhorska-Okolow, M. Metallothioneins in the Lung Cancer. Folia Histochem. Cytobiol. 2015, 53, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, M.Ø.; Larsen, A.; Stoltenberg, M.; Penkowa, M. The Role of Metallothionein in Oncogenesis and Cancer Prognosis. Prog. Histochem. Cytochem. 2009, 44, 29–64. [Google Scholar] [CrossRef] [PubMed]
- Brielrey, J.D.; Gospodarowicz, M.K.; Wittekind, C. Head and Neck Tumours. In TNM Classification of Malignant Tumours, 8th ed.; Brielrey, J.D., Gospodarowicz, M.K., Wittekind, C., Eds.; John Wiley & Sons: Oxford, UK, 2017; Volume 8, pp. 48–52. [Google Scholar]
- Remmele, W.; Stegner, H.E. Recommendation for Uniform Definition of an Immunoreactive Score (IRS) for Immunohistochemical Estrogen Receptor Detection (ER-ICA) in Breast Cancer Tissue. Pathologe 1987, 8, 138–140. [Google Scholar]
- Dziegiel, P.; Salwa-Zurawska, W.; Zurawski, J.; Wojnar, A.; Zabel, M. Prognostic Significance of Augmented Metallothionein (MT) Expression Correlated with Ki-67 Antigen Expression in Selected Soft Tissue Sarcomas. Histol. Histopathol. 2005, 20, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Pastuszewski, W.; Dziegiel, P.; Krecicki, T.; Podhorska-Okolow, M.; Ciesielska, U.; Gorzynska, E.; Zabel, M. Prognostic Significance of Metallothionein, P53 Protein and Ki-67 Antigen Expression in Laryngeal Cancer. Anticancer Res. 2007, 27, 335–342. [Google Scholar]
- Kuloğlu, T.; Artaş, G.; Yardim, M.; Sahin, I.; Aydin, Y.; Beyoğlu, N.; Özercan, İ.H.; Yalcin, M.H.; Ugur, K.; Aydin, S. Immunostaining Characteristics of Irisin in Benign and Malignant Renal Cancers. Biotech. Histochem. 2019, 94, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Altay, D.U.; Keha, E.E.; Karagüzel, E.; Menteşe, A.; Yaman, S.O.; Alver, A. The Diagnostic Value of FNDC5/Irisin in Renal Cell Cancer. Int. braz. J. Urol. 2018, 44, 734–739. [Google Scholar] [CrossRef]
- Provatopoulou, X.; Georgiou, G.P.; Kalogera, E.; Kalles, V.; Matiatou, M.A.; Papapanagiotou, I.; Sagkriotis, A.; Zografos, G.C.; Gounaris, A. Serum Irisin Levels Are Lower in Patients with Breast Cancer: Association with Disease Diagnosis and Tumor Characteristics. BMC Cancer 2015, 15, 898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.; Tang, N.; Qiu, J.; Zhang, D.; Huang, F.; Cheng, Y.; Ding, K.; Li, W.; Zhang, P.; Tan, X. Irisin Stimulates Cell Proliferation and Invasion by Targeting the PI3K/AKT Pathway in Human Hepatocellular Carcinoma. Biochem. Biophys. Res. Commun. 2017, 493, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tan, X.; Tang, N.; Huang, F.; Chen, Z.; Shi, G. Review of Research on the Role of Irisin in Tumors. Onco. Targets. Ther. 2020, 13, 4423–4430. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, G.; Triantafyllidou, S.; Tarlatzis, B.C.; Papakonstantinou, E. Serum Levels of Irisin and Omentin-1 in Breast Neoplasms and Their Association with Tumor Histology. Int. J. Endocrinol. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- Jóźwiak, P.; Lipińska, A. The Role of Glucose Transporter 1 (GLUT1) in the Diagnosis and Therapy of Tumor. Postep. Hig. Med. Dosw. 2012, 66, 165–174. [Google Scholar]
- Xin, C.; Liu, J.; Zhang, J.; Zhu, D.; Wang, H.; Xiong, L.; Lee, Y.; Ye, J.; Lian, K.; Xu, C.; et al. Irisin Improves Fatty Acid Oxidation and Glucose Utilization in Type 2 Diabetes by Regulating the AMPK Signaling Pathway. Int. J. Obes. 2016, 40, 443–451. [Google Scholar] [CrossRef]
- Gos, M.; Miloszewska, J.; Przybyszewska, M. Epithelial-Mesenchymal Transition in Cancer Progression. Postepy Biochem. 2009, 55, 121–128. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Points | Percentage of Cells with Positive Reaction | Points | Color Intensity of Positive Reaction in Cells |
|---|---|---|---|
| 0 | 0% | 0 | lack |
| 1 | 1–10% | 1 | weak |
| 2 | 11–50% | 2 | moderate |
| 3 | 51–80% | 3 | strong |
| 4 | >80% |
| Clinicopathological Parameter | n 140 (%) | Irisin Expression in LSCC Cancer Cells | |||
|---|---|---|---|---|---|
| Low 1–6 | High >6 | Chi2 Test p Value | |||
| Age * | ≤60 >60 | 76 (56,3) 59 (43,7) | 66 (47,10) 51 (36,4) | 10 (7,1) 8 (5,7) | 0.9457 |
| Sex | Male Female | 120 (85,7) 20 (14,3) | 49 (35,0) 9 (6,4) | 71 (50,7) 11 (7,8) | 0.7262 |
| Tumor size (T) | T1–T2 T3–4 | 54 (38,6) 86 (61,4) | 50 (35,7) 70 (50,0) | 4 (2,8) 16 (11,4) | 0.0834 |
| Lymph nodes * (N) | N0 N1 N2–N3 | 98 (72,6) 15 (11,1) 24 (17,7) | 86 (61,4) 13 (9,3) 20 (14,3) | 12 (8,6) 2 (1,4) 4 (2,8) | 0.8475 |
| Stage * | I II III–IV | 7 (5,1) 37 (27,2) 94 (69,1) | 6 (4,3) 37 (26,4) 77 (55,0) | 1 (0,7) 0 (0,0) 17 (12,1) | 0.0216 |
| Grade of malignance (G) | G1 G2 G3 | 30 (21,5) 92 (65,7) 18 (12,8) | 27 (19,3) 79 (56,4) 13 (9,3) | 3 (2,1) 11 (7,8) 4 (2,8) | 0.3803 |
| LSCC | p Value (Mann–Whitney U Test) | Mean Value ± SD | |
|---|---|---|---|
| Lymph nodes N0 vs. N1 N0 vs. N2–3 N1 vs. N2–3 | 0.0031 0.0457 0.5101 | Lymph nodes N0 N1 N2–3 | 5.25 ± 2.02 3.41 ± 2.10 4.00 ± 2.68 |
| Tumor size T1–2 vs. T3–4 | 0.0348 | Tumor size T1–2 T3–4 | 3.00 ± 2.45 3.78 ± 2.58 |
| Stage I vs. II I vs. III–IV II vs. III–IV | 0.2785 0.7991 0.0083 | Stage I II III–IV | 3.46 ± 1.86 2.65 ± 2.52 3.83 ± 2.55 |
| Grade of malignancy G1 vs. G2 G1 vs. G3 G2 vs. G3 | 0.5354 0.2857 0.4415 | Grade of malignancy G1 G2 G3 | 3.26 ± 2.70 3.45 ± 2.47 3.95 ± 2.64 |
| Clinicopathological Parameter | Univariate Analysis HR (95% CI) p |
|---|---|
| pT T1–T2 vs. T3–T4 | 0.98 (0.60–1.62) 0.9787 |
| pN N0 vs. N+ | 0.76 (0.43–1.35) 0.3705 |
| Grade G1 vs. G2–G3 | 0.90 (0.50–1.60) 0.7320 |
| Stage I–II vs. III–IV | 0.80 (0.48–1.33) 0.3883 |
| Irisin <25% vs. ≥25% | 1.46 (0.90–2.40) 0.1183 |
| Ki-67 <25% vs. ≥25% | 1.16 (0.68–1.98) 0.5779 |
| MT- I/II <25% vs. ≥25% | 1.18 (0.72–1.88) 0.5000 |
| MCM3 <25% vs. ≥25% | 1.37 (0.82–2.24) 0.2153 |
| MCM5 <25% vs. ≥25 % | 1.42 (0.89–2.27) 0.1442 |
| MCM7 <25% vs. ≥25% | 3.32 (0.60–18.33) 0.1665 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinkowska, A.; Nowinska, K.; Ciesielska, U.; Podhorska-Okolow, M. Irisin Association with Ki-67, MCM3 and MT-I/II in Squamous Cell Carcinomas of the Larynx. Biomolecules 2022, 12, 52. https://doi.org/10.3390/biom12010052
Pinkowska A, Nowinska K, Ciesielska U, Podhorska-Okolow M. Irisin Association with Ki-67, MCM3 and MT-I/II in Squamous Cell Carcinomas of the Larynx. Biomolecules. 2022; 12(1):52. https://doi.org/10.3390/biom12010052
Chicago/Turabian StylePinkowska, Agnieszka, Katarzyna Nowinska, Urszula Ciesielska, and Marzenna Podhorska-Okolow. 2022. "Irisin Association with Ki-67, MCM3 and MT-I/II in Squamous Cell Carcinomas of the Larynx" Biomolecules 12, no. 1: 52. https://doi.org/10.3390/biom12010052
APA StylePinkowska, A., Nowinska, K., Ciesielska, U., & Podhorska-Okolow, M. (2022). Irisin Association with Ki-67, MCM3 and MT-I/II in Squamous Cell Carcinomas of the Larynx. Biomolecules, 12(1), 52. https://doi.org/10.3390/biom12010052