
Stimulation of Neurite Outgrowth Using Autologous NGF Bound at the Surface of a Fibrous Substrate





Abstract
:1. Introduction
2. Materials and Methods
2.1. Rat Blood Plasma
2.2. Production of Electrospun Fibrous Meshes
2.3. Nerve Growth Factor Functionalized Electrospun Fibrous Meshes
2.4. Biological In Vitro Assays
2.4.1. PC12 Cells Culture
2.4.2. Cellular Biochemistry Analysis
2.4.3. Scanning Electron Microscopy (SEM)
2.4.4. Gene Expression Analysis
2.4.5. Immunocytochemistry
2.4.6. Neurite Outgrowth
2.5. Statistical Analysis
3. Results
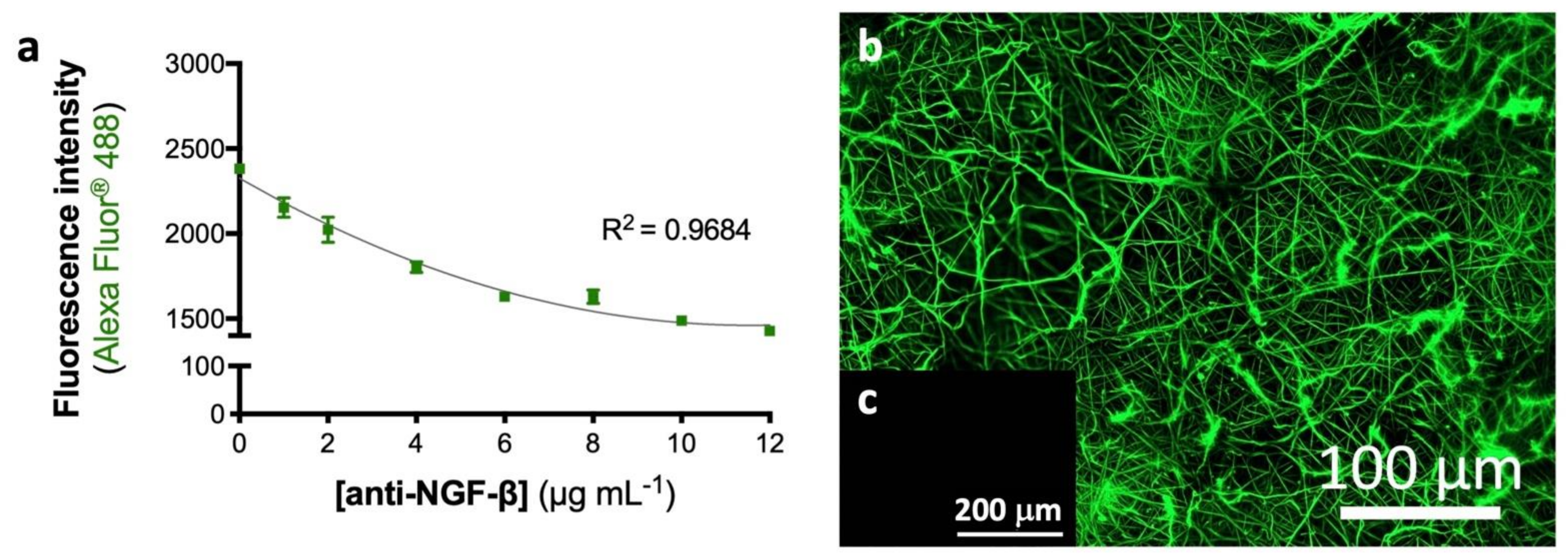
3.1. Physicochemical Properties of the eFM Functionalized with NGF
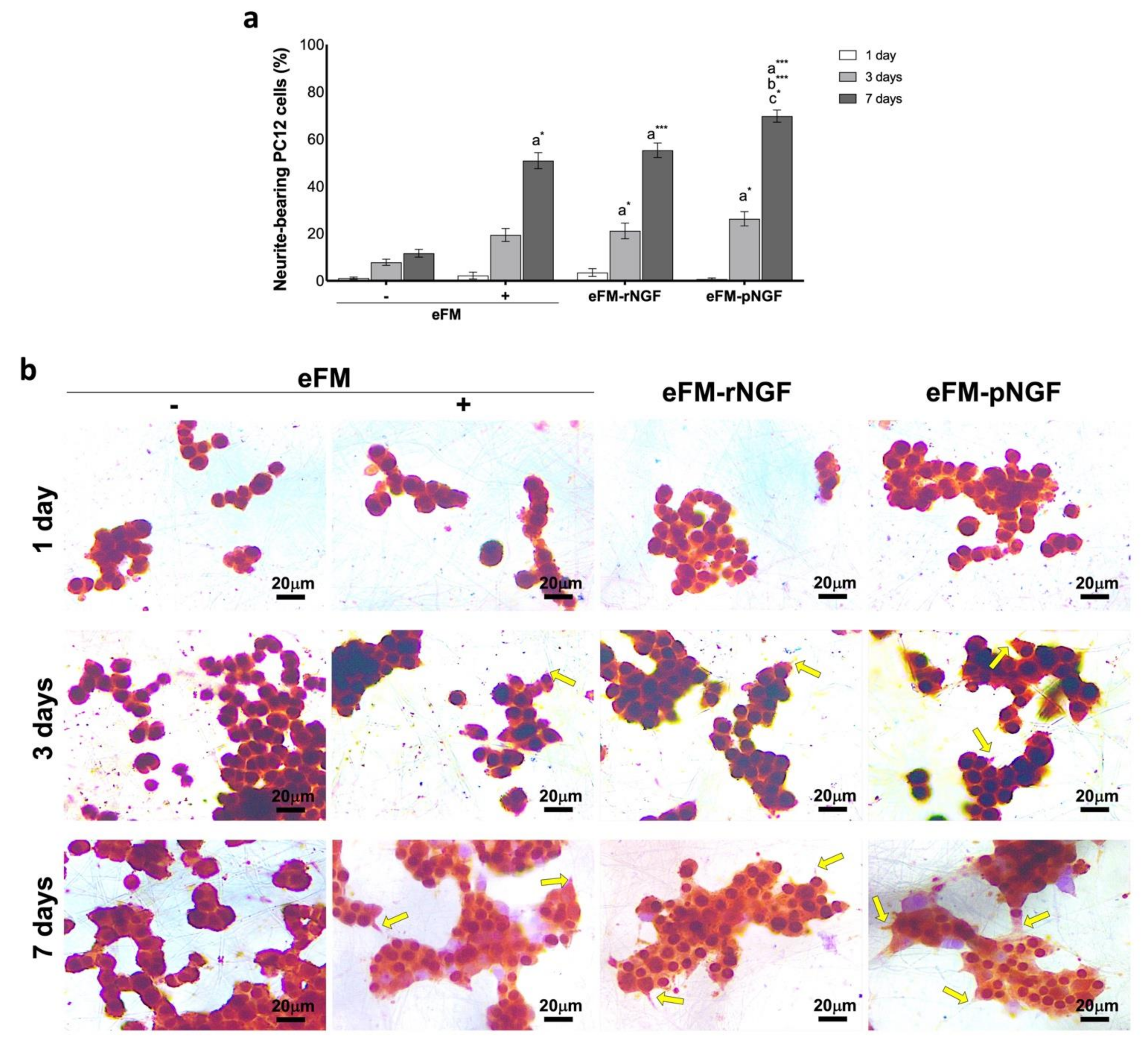
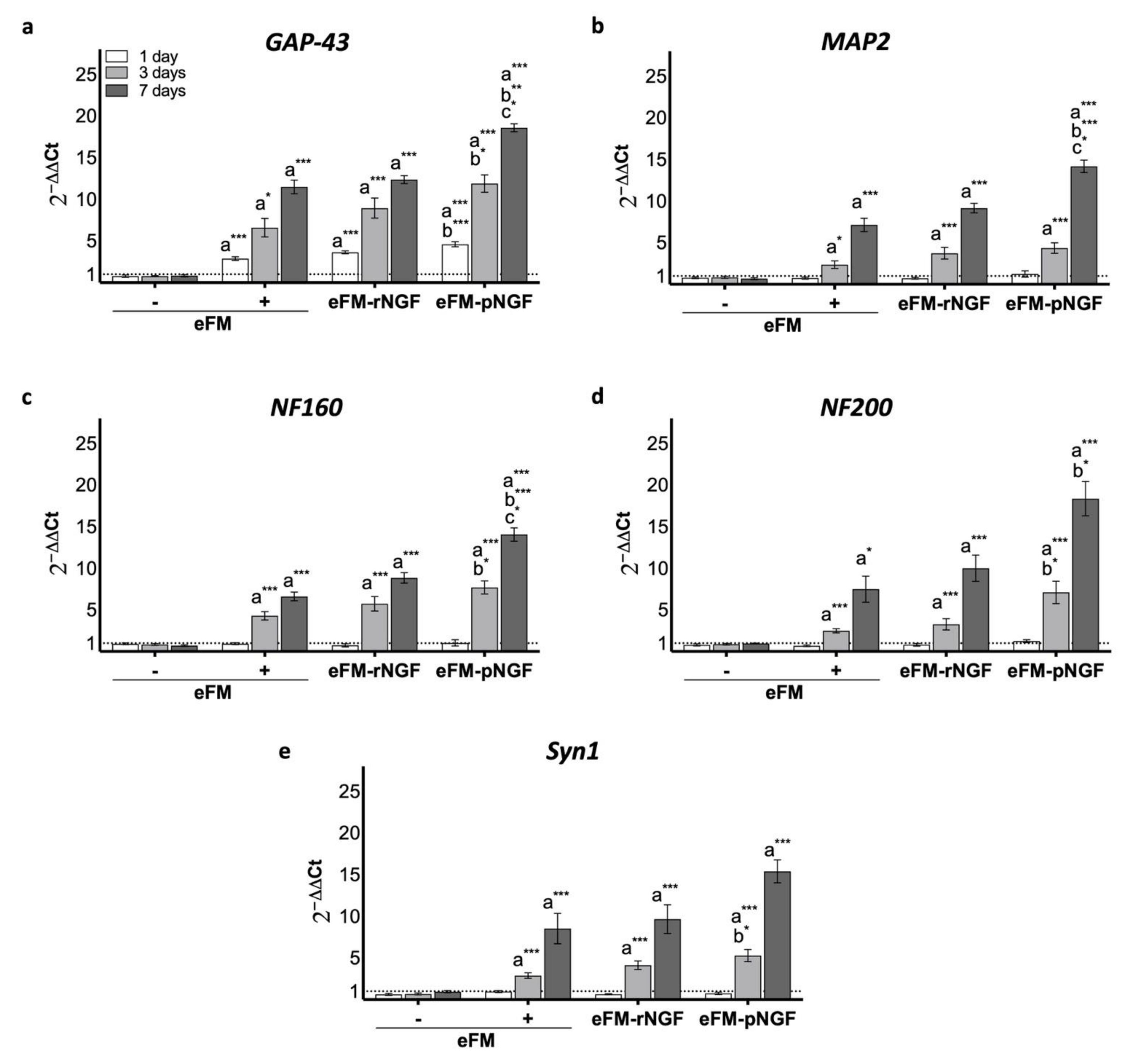
3.2. Performance of PC12 Cells Cultured on eFM-NGF
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, C.A.; Braza, D.; Rice, J.B.; Dillingham, T. The incidence of peripheral nerve injury in extremity trauma. Am. J. Phys. Med. Rehabil. 2008, 87, 381–385. [Google Scholar] [CrossRef]
- Wang, W.; Gao, J.; Na, L.; Jiang, H.; Xue, J.; Yang, Z.; Wang, P. Craniocerebral injury promotes the repair of peripheral nerve injury. Neural Regen. Res. 2014, 9, 1703–1708. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.P.; Freeman, M.R. Wallerian degeneration, wld(s), and nmnat. Annu. Rev. Neurosci. 2010, 33, 245–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, B.; Quan, Q.; Lu, S.; Wang, Y.; Peng, J. Molecular mechanisms in the initiation phase of Wallerian degeneration. Eur. J. Neurosci. 2016, 44, 2040–2048. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nature Commun. 2019, 10, 3879. [Google Scholar] [CrossRef]
- Deumens, R.; Bozkurt, A.; Meek, M.F.; Marcus, M.A.; Joosten, E.A.; Weis, J.; Brook, G.A. Repairing injured peripheral nerves: Bridging the gap. Prog. Neurobiol. 2010, 92, 245–276. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.W.; Hogan, M.V.; Chhabra, A.B.; Deal, D.N. Peripheral nerve repair and reconstruction. J. Bone. Joint. Surg. Am. 2013, 95, 2144–2151. [Google Scholar] [CrossRef]
- Bulstra, L.F.; Rbia, N.; Kircher, M.F.; Spinner, R.J.; Bishop, A.T.; Shin, A.Y. Spinal accessory nerve to triceps muscle transfer using long autologous nerve grafts for recovery of elbow extension in traumatic brachial plexus injuries. J. Neurosurg. 2018, 129, 1041–1047. [Google Scholar] [CrossRef] [Green Version]
- Geuna, S.; Papalia, I.; Ronchi, G.; d’Alcontres, F.S.; Natsis, K.; Papadopulos, N.A.; Colonna, M.R. The reasons for end-to-side coaptation: How does lateral axon sprouting work? Neural Regen. Res. 2017, 12, 529–533. [Google Scholar] [CrossRef]
- Millesi, H. Bridging defects: Autologous nerve grafts. Acta Neurochir. Suppl. 2007, 100, 37–38. [Google Scholar]
- Sarker, M.D.; Naghieh, S.; McInnes, A.D.; Schreyer, D.J.; Chen, X. Regeneration of peripheral nerves by nerve guidance conduits: Influence of design, biopolymers, cells, growth factors, and physical stimuli. Prog. Neurobiol. 2018, 171, 125–150. [Google Scholar] [CrossRef]
- Forbes, S.J.; Rosenthal, N. Preparing the ground for tissue regeneration: From mechanism to therapy. Nat. Med. 2014, 20, 857–869. [Google Scholar] [CrossRef]
- Marquardt, L.M.; Sakiyama-Elbert, S.E. Engineering peripheral nerve repair. Curr. Opin. Biotechnol. 2013, 24, 887–892. [Google Scholar] [CrossRef] [Green Version]
- Grinsell, D.; Keating, C.P. Peripheral Nerve Reconstruction after Injury: A Review of Clinical and Experimental Therapies. Biomed. Res. Int. 2014, 2014, 698256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.L.; Rivlin, M.; Graham, J.G.; Beredjiklian, P.K. Peripheral nerve injury, scarring, and recovery. Connect. Tissue Res. 2019, 60, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Belanger, K.; Dinis, T.M.; Taourirt, S.; Vidal, G.; Kaplan, D.L.; Egles, C. Recent Strategies in Tissue Engineering for Guided Peripheral Nerve Regeneration. Macromol. Biosci. 2016, 16, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, Y.L.; Kong, D.; Qu, B.; Su, X.J.; Li, H.; Pi, H.Y. Nerve autografts and tissue-engineered materials for the repair of peripheral nerve injuries: A 5-year bibliometric analysis. Neural. Regen. Res. 2015, 10, 1003–1008. [Google Scholar] [CrossRef]
- Moattari, M.; Kouchesfehani, H.M.; Kaka, G.; Sadraie, S.H.; Naghdi, M. Evaluation of nerve growth factor (NGF) treated mesenchymal stem cells for recovery in neurotmesis model of peripheral nerve injury. J. Cranio. Maxill. Surg. 2018, 46, 898–904. [Google Scholar] [CrossRef]
- Li, G.C.; Xiao, Q.Z.; Zhang, L.Z.; Zhao, Y.H.; Yang, Y.M. Nerve growth factor loaded heparin/chitosan scaffolds for accelerating peripheral nerve regeneration. Carbohyd. Polym. 2017, 171, 39–49. [Google Scholar] [CrossRef]
- Zeng, W.; Rong, M.Y.; Hu, X.Y.; Xiao, W.; Qi, F.Y.; Huang, J.H.; Luo, Z.J. Incorporation of Chitosan Microspheres into Collagen-Chitosan Scaffolds for the Controlled Release of Nerve Growth Factor. PLoS ONE 2014, 9, e101300. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, C.; Pedro, J.; Ng, K.W.; Neves, N.; Reis, R.; Oliveira, M. Keratin/chitosan as novel grafts for peripheral nerve regeneration. J. Tissue. Eng. Regen. M 2014, 8, 434. [Google Scholar]
- Salgado, A.J.; Sousa, N.; Silva, N.A.; Neves, N.M.; Reis, R.L. Hydrogels for spinal cord injury regeneration. Woodhead Publ. Mater. 2008, 22, 570–594. [Google Scholar] [CrossRef]
- Qian, Y.; Zhao, X.T.; Han, Q.X.; Chen, W.; Li, H.; Yuan, W.E. An integrated multi-layer 3D-fabrication of PDA/RGD coated graphene loaded PCL nanoscaffold for peripheral nerve restoration. Nat. Commun. 2018, 9, 323. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Asikainen, S.; Teotia, A.K.; Shiekh, P.A.; Huotilainen, E.; Qayoom, I.; Partanen, J.; Seppala, J.; Kumar, A. Biomimetic Photocurable Three-Dimensional Printed Nerve Guidance Channels with Aligned Cryomatrix Lumen for Peripheral Nerve Regeneration. ACS Appl. Mater. Inter. 2018, 10, 43327–43342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, W.; Tao, K.X.; Song, Y.; Xie, H.J.; Wang, J.; Li, X.L.; Shuai, X.M.; Gao, J.B.; Chang, P.P.; et al. Sustained Local Release of NGF from a Chitosan-Sericin Composite Scaffold for Treating Chronic Nerve Compression. ACS Appl. Mater. Inter. 2017, 9, 3432–3444. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.C.; Tseng, T.C.; Chen, J.R.; Hsu, S.H.; Chiu, I.M. Fabrication of bioactive conduits containing the fibroblast growth factor 1 and neural stem cells for peripheral nerve regeneration across a 15 mm critical gap. Biofabrication 2013, 5, 035010. [Google Scholar] [CrossRef]
- Koh, H.S.; Yong, T.; Chan, C.K.; Ramakrishna, S. Enhancement of neurite outgrowth using nano-structured scaffolds coupled with laminin. Biomaterials 2008, 29, 3574–3582. [Google Scholar] [CrossRef]
- Quan, Q.; Meng, H.Y.; Chang, B.; Liu, G.B.; Cheng, X.Q.; Tang, H.; Wang, Y.; Peng, J.; Zhao, Q.; Lu, S.B. Aligned fibers enhance nerve guide conduits when bridging peripheral nerve defects focused on early repair stage. Neural Regen. Res. 2019, 14, 903–912. [Google Scholar] [CrossRef]
- Wang, J.; Tian, L.L.; He, L.M.; Chen, N.; Ramakrishna, S.; So, K.F.; Mo, X.M. Lycium barbarum polysaccharide encapsulated Poly lactic-co-glycolic acid Nanofibers: Cost effective herbal medicine for potential application in peripheral nerve tissue engineering. Sci. Rep. 2018, 8, 8669. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.J.; Li, H.X.; Xia, Y.N. Nanofiber-Based Multi-Tubular Conduits with a Honeycomb Structure for Potential Application in Peripheral Nerve Repair. Macromol. Biosci. 2018, 18, 1800090. [Google Scholar] [CrossRef]
- Tajdaran, K.; Gordon, T.; Wood, M.D.; Shoichet, M.S.; Borschel, G.H. A glial cell line-derived neurotrophic factor delivery system enhances nerve regeneration across acellular nerve allografts. Acta. Biomater. 2016, 29, 62–70. [Google Scholar] [CrossRef]
- Terenghi, G. Peripheral nerve regeneration and neurotrophic factors. J. Anat. 1999, 194, 1–14. [Google Scholar] [CrossRef]
- Afrash, H.; Nazeri, N.; Davoudi, P.; Faridi Majidi, R.; Ghanbari, H. Development of a Bioactive Scaffold based on NGF Containing PCL/Chitosan Nanofibers for Nerve Regeneration. Biointerface Res. App. 2021, 11, 12606–12617. [Google Scholar] [CrossRef]
- Liu, C.Y.; Li, X.H.; Xu, F.Y.; Cong, H.B.; Li, Z.X.; Song, Y.; Wang, M. Spatio-temporal release of NGF and GDNF from multi-layered nanofibrous bicomponent electrospun scaffolds. J. Mater. Sci.-Mater. M 2018, 29, 102. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y.; Li, X.H.; Zhao, Q.L.; Xie, Y.C.; Yao, X.M.; Wang, M.; Cao, F.J. Nanofibrous bicomponent scaffolds for the dual delivery of NGF and GDNF: Controlled release of growth factors and their biological effects. J. Mater. Sci.-Mater. M 2021, 32, 9. [Google Scholar] [CrossRef] [PubMed]
- Okur, Z.; Senturk, O.I.; Yilmaz, C.; Gulseren, G.; Mammadov, B.; Guler, M.O.; Tekinay, A.B. Promotion of neurite outgrowth by rationally designed NGF-beta binding peptide nanofibers. Biomater. Sci. 2018, 6, 1777–1790. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Castellanos, A.M.; Claeyssens, F.; Haycock, J.W. Bioactive 3D Scaffolds for the Delivery of NGF and BDNF to Improve Nerve Regeneration. Front. Mater. 2021, 8, 734683. [Google Scholar] [CrossRef]
- Zhang, R.R.; Zhang, Y.; Yi, S. Identification of critical growth factors for peripheral nerve regeneration. Rsc. Adv. 2019, 9, 10760–10765. [Google Scholar] [CrossRef] [Green Version]
- Santosa, K.B.; Jesuraj, N.J.; Viader, A.; MacEwan, M.; Newton, P.; Hunter, D.A.; Mackinnon, S.E.; Johnson, P.J. Nerve allografts supplemented with schwann cells overexpressing glial-cell-line-derived neurotrophic factor. Muscle. Nerve 2013, 47, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Etulain, J.; Mena, H.A.; Meiss, R.P.; Frechtel, G.; Gutt, S.; Negrotto, S.; Schattner, M. An optimised protocol for plateletrich plasma preparation to improve its angiogenic and regenerative properties. Sci. Rep. 2018, 8, 1513. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.R.; da Silva, M.A.; Costa-Pinto, A.R.; Reis, R.L.; Martins, A.; Neves, N.M. Chondrogenesis-inductive nanofibrous substrate using both biological fluids and mesenchymal stem cells from an autologous source. Mat. Sci. Eng. C Mater. 2019, 98, 1169–1178. [Google Scholar] [CrossRef]
- da Silva, M.A.; Crawford, A.; Mundy, J.; Martins, A.; Araujo, J.V.; Hatton, P.V.; Reis, R.L.; Neves, N.M. Evaluation of Extracellular Matrix Formation in Polycaprolactone and Starch-Compounded Polycaprolactone Nanofiber Meshes When Seeded with Bovine Articular Chondrocytes. Tissue. Eng. Pt. A 2009, 15, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, N.; Martins, A.; Pires, R.; Faria, S.; Fonseca, N.A.; Moreira, J.N.; Reisa, R.L.; Neves, N.M. Immobilization of bioactive factor-loaded liposomes on the surface of electrospun nanofibers targeting tissue engineering. Biomater. Sci. 2014, 2, 1195–1209. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.M.; Zhao, W.J.; He, J.H.; Zhao, Y.H.; Ding, F.; Gu, X.S. Nerve conduits based on immobilization of nerve growth factor onto modified chitosan by using genipin as a crosslinking agent. Eur. J. Pharm. Biopharm. 2011, 79, 519–525. [Google Scholar] [CrossRef]
- Gu, X.; Ding, F.; Williams, D.F. Neural tissue engineering options for peripheral nerve regeneration. Biomaterials 2014, 35, 6143–6156. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, H.; Hiraka, K.; Nagaoka, S.; Suzuki, Y.; Iwaki, M. Neuronal attachment and outgrowth on a micropatterned fluorinated polyimide surface. J. Artif. Organs. 2004, 7, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Costa-Pinto, A.R.; Reis, R.L.; Martins, A.; Neves, N.M. Biofunctional nanofibrous substrate comprising immobilized antibodies and selective binding of autologous growth factors. Biomacromolecules 2014, 15, 2196–2205. [Google Scholar] [CrossRef]
- Casanova, M.R.; Oliveira, C.; Fernandes, E.; Reis, R.; Silva, T.H.; Martins, A.; Neves, N.M. Spatial immobilization of endogenous growth factors to control vascularization in bone tissue engineering. Biomater. Sci. 2020, 8, 2577–2589. [Google Scholar] [CrossRef]
- Casanova, M.R.; Osório, H.; Reis, R.L.; Martins, A.; Neves, N.M. Chondrogenic differentiation induced by extracellular vesicles bound to a nanofibrous substrate. NPJ Regen. Med. 2021, 6, 79. [Google Scholar] [CrossRef]
- Casanova, M.R.; Reis, R.L.; Martins, A.; Neves, N.M. Fibronectin Bound to a Fibrous Substrate Has Chondrogenic Induction Properties. Biomacromolecules 2020, 21, 1368–1378. [Google Scholar] [CrossRef]
- Bacelo, E.; da Silva, M.A.; Cunha, C.; Faria, S.; Carvalho, A.; Reis, R.L.; Martins, A.; Neves, N.M. Biofunctional Nanofibrous Substrate for Local TNF-Capturing as a Strategy to Control Inflammation in Arthritic Joints. Nanomaterials 2019, 9, 567. [Google Scholar] [CrossRef] [Green Version]
- Bastami, F.; Vares, P.; Khojasteh, A. Healing Effects of Platelet-Rich Plasma on Peripheral Nerve Injuries. J. Craniofac. Surg. 2017, 28, e49–e57. [Google Scholar] [CrossRef]
- Sanchez, M.; Garate, A.; Delgado, D.; Padilla, S.P. Platelet-rich plasma, an adjuvant biological therapy to assist peripheral nerve repair. Neural. Regen. Res. 2017, 12, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.P.; Lyon, K.A.; Huang, J.H. An update-tissue engineered nerve grafts for the repair of peripheral nerve injuries. Neural. Regen. Res. 2018, 13, 764–774. [Google Scholar] [CrossRef]
- Sanchez, M.; Anitua, E.; Delgado, D.; Sanchez, P.; Prado, R.; Orive, G.; Padilla, S. Platelet-rich plasma, a source of autologous growth factors and biomimetic scaffold for peripheral nerve regeneration. Expert. Opin. Biol. Ther. 2017, 17, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Hadji Constantinou, M.; McGuire, L.; Duchemin, A.M.; Laskowski, B.; Kiecolt-Glaser, J.; Glaser, R. Changes in plasma nerve growth factor levels in older adults associated with chronic stress. J. Neuroimmunol. 2001, 116, 102–106. [Google Scholar] [CrossRef]
- Lee, B.C.; Choi, I.G.; Kim, Y.K.; Ham, B.J.; Yang, B.H.; Roh, S.; Choi, J.; Lee, J.S.; Oh, D.Y.; Chai, Y.G. Relation between plasma brain-derived neurotrophic factor and nerve growth factor in the male patients with alcohol dependence. Alcohol 2009, 43, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Emanuele, E.; Politi, P.; Bianchi, M.; Minoretti, P.; Bertona, M.; Geroldi, D. Raised plasma nerve growth factor levels associated with early-stage romantic love. Psychoneuroendocrinology 2006, 31, 288–294. [Google Scholar] [CrossRef]
- Levanti, M.B.; Germana, A.; de Carlos, F.; Ciriaco, E.; Vega, J.A.; Germana, G. Effects of increased nerve growth factor plasma levels on the expression of TrkA and p75 in rat testicles. J. Anat. 2006, 208, 373–379. [Google Scholar] [CrossRef]
- Aloe, L.; Rossi, S.; Manni, L. Altered expression of nerve growth factor and its receptors in the kidneys of diabetic rats. J. Nephrol. 2011, 24, 798–805. [Google Scholar] [CrossRef]
- Liu, J.; Li, J.D.; Lu, J.; Xing, J.; Li, J. Contribution of nerve growth factor to upregulation of P2X(3) expression in DRG neurons of rats with femoral artery occlusion. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H1070–H1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strategies | Pros | Cons | |
|---|---|---|---|
| Nerve Grafts | Autograft |
|
|
| Allograft |
|
| |
| Xenograft |
|
| |
| Tissue Engineered Nerve Guidance Grafts |
|
| |
| Condition | Description | |
|---|---|---|
| eFM | − | eFM in BM 1 without rNGF |
| + | eFM in BM supplemented with 100 ng/mL of rNGF | |
| eFM-rNGF | eFM functionalized with rNGF in BM | |
| eFM-pNGF | eFM functionalized with plasma-derived NGF in BM | |
| Gene 1 | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| GAPDH | CAACTCCCTCAAGATTGTCAGCAA | GGCATGGACTGTGGTCATGA |
| GAP-43 | TTTCCTCTCCTGTCCTGCTC | TGGACTTGGGATCTTTCCTG |
| MAP2 | GGCACTCCTCCAAGCTACTCT | CTTGACGTTCTTCAGGTCTGG |
| NF-160 | AGCATTGAGCTCGAGTCGGTG | CTGCTGGATGGTGTCCTGGTAG |
| NF-200 | AAAGTGAACACGGATGCTATGC | GTGCTTTTCAGTGCCTCCAAC |
| Syn1 | GTGTCAGGGAACTGGAAGACC | AGGAGCCCACCACCTCAATA |
| Recombinant (ng/mL) | Plasma (ρg/mL) | |
|---|---|---|
| [NGF] | 99.9 ± 0.01 | 597 ± 40 |
| bound | 98.7 ± 0.2 | 567 ± 23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casanova, M.R.; Reis, R.L.; Martins, A.; Neves, N.M. Stimulation of Neurite Outgrowth Using Autologous NGF Bound at the Surface of a Fibrous Substrate. Biomolecules 2022, 12, 25. https://doi.org/10.3390/biom12010025
Casanova MR, Reis RL, Martins A, Neves NM. Stimulation of Neurite Outgrowth Using Autologous NGF Bound at the Surface of a Fibrous Substrate. Biomolecules. 2022; 12(1):25. https://doi.org/10.3390/biom12010025
Chicago/Turabian StyleCasanova, Marta R., Rui L. Reis, Albino Martins, and Nuno M. Neves. 2022. "Stimulation of Neurite Outgrowth Using Autologous NGF Bound at the Surface of a Fibrous Substrate" Biomolecules 12, no. 1: 25. https://doi.org/10.3390/biom12010025
APA StyleCasanova, M. R., Reis, R. L., Martins, A., & Neves, N. M. (2022). Stimulation of Neurite Outgrowth Using Autologous NGF Bound at the Surface of a Fibrous Substrate. Biomolecules, 12(1), 25. https://doi.org/10.3390/biom12010025