
Melittin-Based Nano-Delivery Systems for Cancer Therapy

Abstract
1. Introduction
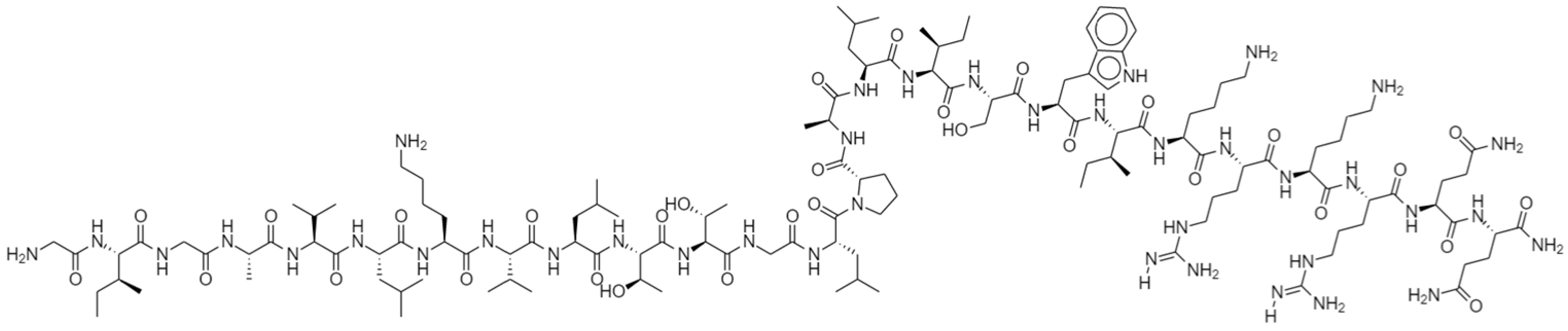
1.1. Structure of Melittin (MEL) and Its Interactions with Membrane
1.2. Pharmacological Effect of MEL
1.3. Anti-Tumor Effects of MEL

{kind=link}
{kind=link}
| Tumor Type | Cell Lines | Treatment | Result or Mechanism | Reference |
|---|---|---|---|---|
| Lung cancer | A549 and NCI-H460 cell | MEL | IC50 values were 2 μg/mL, 3 μg/mL, respectively | [45] |
| A549 cell | Antinucleolin aptamer–MEL conjugate | Viability for A549 cells after treatment was 51.2 ± 3.5%, | [46] | |
| Hepatocellular carcinoma | SMMC-7721 cells | MEL | MEL inhibits G0/G1 cell cycle progression by down-regulating MeCP2 through Shh signaling. | [47] |
| HepG2 cells | MEL | HDAC2-mediated PTEN upregulation, Akt inactivation, and inhibition of PI3K/Akt signaling pathways. | [48] | |
| SMMC-7721 and BEL-7402 cells | MEL | MEL sensitizes human hepatocellular carcinoma cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis by activating CaMKII-TAK1-JNK/p38 and inhibiting IκBαkinase-NFκB. | [49] | |
| Breast cancer | MDA-MB-231 cells | MEL | MEL inhibits the EGF-induced MMP-9 expression via blocking the NF-κB and PI3K/Akt/mTOR pathway | [50] |
| SUM159 and SKBR3 | BV or MEL | MEL reduces the level of the PD-L1 immune-checkpoint protein and the immune-suppressive effects of the tumor microenvironment. IC50 values for MEL was 4.24 ng/μL for SUM159 and 3.59 ng/μL for SKBR3. | [51] | |
| Prostate cancer | LNCaP, DU145, and PC-3 cells | BV or MEL | MEL induces cell apoptosis by activating the caspase pathway via NF-κB inactivation. IC50 for LNCaP cells: MEL 2.9 and BV 14.2 µg/mL, DU145 cells: MEL 1.5 and BV 6.3 µg/mL IC50 for PC-3 cells: MEL 1.8 and BV 6.1 µg/mL, respectively | [52] |
| Leukemia | CCRF-CEM and K-562 cells | MEL | MEL induces apoptosis via the intrinsic/mitochondrial pathway. | [53] |
1.4. Obstacles to the Applications of MEL
2. Delivery Vehicles for Melittin
2.1. Modified MEL and Conjugates
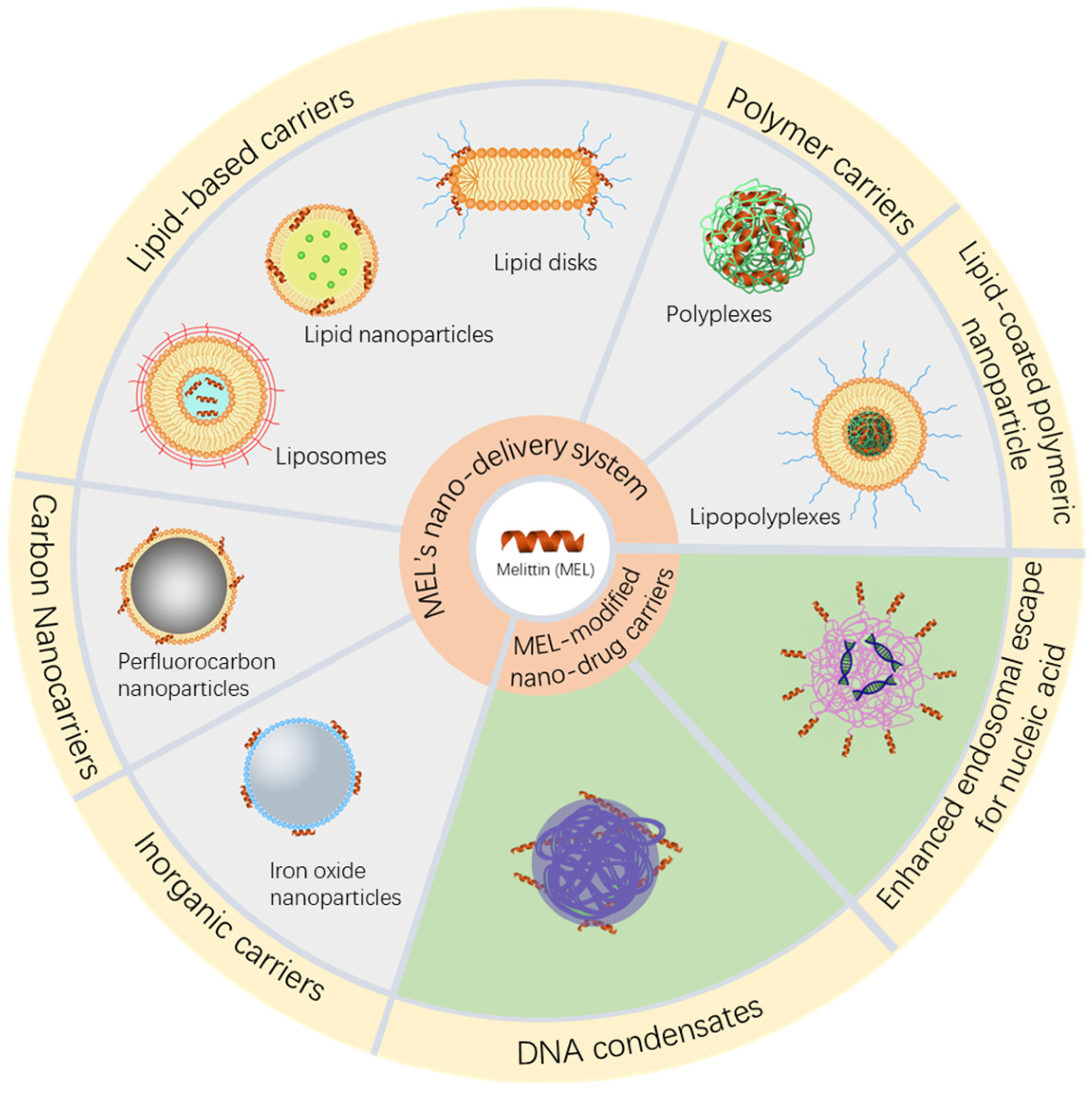
2.2. Nano Delivery Vehicles
2.2.1. Inorganic Carriers
2.2.2. Carbon Nanocarriers
2.2.3. Polymer Carriers
2.2.4. Lipid-Based Carriers
2.2.5. Lipid-Coated Polymeric Nanoparticles
2.2.6. Stimulus-Responsive Delivery Systems
3. Nano Drug Delivery System with Melittin as a Functional Molecule
3.1. Melittin Enables Efficient Vesicular Escape
3.2. Enhanced Drug Delivery of MEL as an Adjuvant
4. Conclusions and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Habermann, E. Bee and Wasp Venoms. Science 1972, 177, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Maulet, Y.; Brodbeck, U.; Fulpius, B.W. Purification from bee venom of melittin devoid of phospholipase A2 contamination. Anal. Biochem. 1982, 127, 61–67. [Google Scholar] [CrossRef]
- Pacáková, V.; Štulík, K.; Thi Hau, P.; Jelínek, I.; Vinš, I.; Sýkora, D. Comparison of high-performance liquid chromatography and capillary electrophoresis for the determination of some bee venom components. J. Chromatogr. A 1995, 700, 187–193. [Google Scholar] [CrossRef]
- Chen, J.; Guan, S.M.; Sun, W.; Fu, H. Melittin, the Major Pain-Producing Substance of Bee Venom. Neurosci. Bull. 2016, 32, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Raghuraman, H.; Chattopadhyay, A. Melittin: A Membrane-active Peptide with Diverse Functions. Biosci. Rep. 2007, 27, 189–223. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, Y.; Kataoka, M.; Aimoto, S.; Goto, Y. Charge repulsion in the conformational stability of melittin. Biochemistry 1992, 31, 11908–11914. [Google Scholar] [CrossRef]
- Bello, J.; Bello, H.R.; Granados, E. Conformation and aggregation of melittin: Dependence on pH and concentration. Biochemistry 1982, 21, 461–465. [Google Scholar] [CrossRef] [PubMed]
- DeGrado, W.F.; Musso, G.F.; Lieber, M.; Kaiser, E.T.; Kézdy, F.J. Kinetics and mechanism of hemolysis induced by melittin and by a synthetic melittin analogue. Biophys. J. 1982, 37, 329–338. [Google Scholar] [CrossRef]
- Zorilă, B.; Necula, G.; Radu, M.; Bacalum, M. Melittin Induces Local Order Changes in Artificial and Biological Membranes as Revealed by Spectral Analysis of Laurdan Fluorescence. Toxins 2020, 12, 705. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu, I.; Lafleur, M. Influence of the lipid composition on the kinetics of concerted insertion and folding of melittin in bilayers. Biochim. Biophys. Acta (BBA)-Biomembr. 2004, 1667, 26–37. [Google Scholar] [CrossRef]
- Sessa, G.; Freer, J.H.; Colacicco, G.; Weissmann, G. Interaction of a Lytic Polypeptide, Melittin, with Lipid Membrane Systems. J. Biol. Chem. 1969, 244, 3575–3582. [Google Scholar] [CrossRef]
- Picoli, T.; Peter, C.M.; Zani, J.L.; Waller, S.B.; Lopes, M.G.; Boesche, K.N.; Vargas, G.D.Á.; Hübner, S.d.O.; Fischer, G. Melittin and its potential in the destruction and inhibition of the biofilm formation by Staphylococcus aureus, Escherichia coli and Pseudomonas aeruginosa isolated from bovine milk. Microb. Pathog. 2017, 112, 57–62. [Google Scholar] [CrossRef]
- Uddin, M.B.; Lee, B.H.; Nikapitiya, C.; Kim, J.H.; Kim, T.H.; Lee, H.C.; Kim, C.G.; Lee, J.S.; Kim, C.J. Inhibitory effects of bee venom and its components against viruses in vitro and in vivo. J. Microbiol. 2016, 54, 853–866. [Google Scholar] [CrossRef]
- Lee, W.; Lee, D.G. Fungicidal mechanisms of the antimicrobial peptide Bac8c. Biochim. Biophys. Acta 2015, 1848, 673–679. [Google Scholar] [CrossRef]
- Lee, D.G.; Park, J.H.; Shin, S.Y.; Lee, S.G.; Lee, M.K.; Kim, K.L.; Hahm, K.S. Design of novel analogue peptides with potent fungicidal but low hemolytic activity based on the cecropin A-melittin hybrid structure. Biochem Mol. Biol. Int. 1997, 43, 489–498. [Google Scholar] [CrossRef]
- Lee, G.; Bae, H. Anti-Inflammatory Applications of Melittin, a Major Component of Bee Venom: Detailed Mechanism of Action and Adverse Effects. Molecules 2016, 21, 616. [Google Scholar] [CrossRef] [PubMed]
- Wachinger, M.; Kleinschmidt, A.; Winder, D.; von Pechmann, N.; Ludvigsen, A.; Neumann, M.; Holle, R.; Salmons, B.; Erfle, V.; Brack-Werner, R. Antimicrobial peptides melittin and cecropin inhibit replication of human immunodeficiency virus 1 by suppressing viral gene expression. J. Gen. Virol. 1998, 79, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Albiol Matanic, V.C.; Castilla, V. Antiviral activity of antimicrobial cationic peptides against Junin virus and herpes simplex virus. Int. J. Antimicrob. Agents 2004, 23, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Al-Rabia, M.W.; Alhakamy, N.A.; Ahmed, O.A.A.; Eljaaly, K.; Aloafi, A.L.; Mostafa, A.; Asfour, H.Z.; Aldarmahi, A.A.; Darwish, K.M.; Ibrahim, T.S.; et al. Repurposing of Sitagliptin- Melittin Optimized Nanoformula against SARS-CoV-2: Antiviral Screening and Molecular Docking Studies. Pharmaceutics 2021, 13, 307. [Google Scholar] [CrossRef]
- Shams Khozani, R.; Shahbazzadeh, D.; Harzandi, N.; Feizabadi, M.M.; Pooshang Bagheri, K. Kinetics Study of Antimicrobial Peptide, Melittin, in Simultaneous Biofilm Degradation and Eradication of Potent Biofilm Producing MDR Pseudomonas aeruginosa Isolates. Int. J. Pept. Res. Ther. 2019, 25, 329–338. [Google Scholar] [CrossRef]
- Marques Pereira, A.F.; Albano, M.; Bérgamo Alves, F.C.; Murbach Teles Andrade, B.F.; Furlanetto, A.; Mores Rall, V.L.; Delazari dos Santos, L.; de Oliveira Orsi, R.; Fernandes Júnior, A. Influence of apitoxin and melittin from Apis mellifera bee on Staphylococcus aureus strains. Microb. Pathog. 2020, 141, 104011. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Kamysz, W.; D’Amato, G.; Silvestri, C.; Del Prete, M.S.; Łukasiak, J.; Scalise, G. Comparative activities of cecropin A, melittin, and cecropin A–melittin peptide CA(1–7)M(2–9)NH2 against multidrug-resistant nosocomial isolates of Acinetobacter baumannii. Peptides 2003, 24, 1315–1318. [Google Scholar] [CrossRef] [PubMed]
- Saraswat, J.; Aldahmash, B.; AlOmar, S.Y.; Imtiyaz, K.; Rizvi, M.M.A.; Patel, R. Synergistic antimicrobial activity of N-methyl substituted pyrrolidinium–based ionic liquids and melittin against Gram-positive and Gram-negative bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 10465–10479. [Google Scholar] [CrossRef]
- Akbari, R.; Hakemi-Vala, M.; Pashaie, F.; Bevalian, P.; Hashemi, A.; Pooshang Bagheri, K. Highly Synergistic Effects of Melittin with Conventional Antibiotics Against Multidrug-Resistant Isolates of Acinetobacter baumannii and Pseudomonas aeruginosa. Microb. Drug Resist. 2019, 25, 193–202. [Google Scholar] [CrossRef]
- Lee, J.; Hwang, J.-S.; Hwang, I.-s.; Cho, J.; Lee, E.; Kim, Y.; Lee, D.G. Coprisin-induced antifungal effects in Candida albicans correlate with apoptotic mechanisms. Free. Radic. Biol. Med. 2012, 52, 2302–2311. [Google Scholar] [CrossRef]
- Jia, F.; Wang, J.; Peng, J.; Zhao, P.; Kong, Z.; Wang, K.; Yan, W.; Wang, R. The in vitro, in vivo antifungal activity and the action mode of Jelleine-I against Candida species. Amino Acids 2018, 50, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Lange, W. The germicidal effect of bee venom. Muench Med. Wochenschr 1941, 83, 935. [Google Scholar]
- Memariani, H.; Memariani, M.; Shahidi-Dadras, M.; Nasiri, S.; Akhavan, M.M.; Moravvej, H. Melittin: From honeybees to superbugs. Appl. Microbiol. Biotechnol. 2019, 103, 3265–3276. [Google Scholar] [CrossRef]
- Jiang, X.; Qian, K.; Liu, G.; Sun, L.; Zhou, G.; Li, J.; Fang, X.; Ge, H.; Lv, Z. Design and activity study of a melittin-thanatin hybrid peptide. AMB Express 2019, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Memariani, H.; Memariani, M.; Moravvej, H.; Shahidi-Dadras, M. Melittin: A venom-derived peptide with promising anti-viral properties. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Hood, J.L.; Jallouk, A.P.; Campbell, N.; Ratner, L.; Wickline, S.A. Cytolytic nanoparticles attenuate HIV-1 infectivity. Antivir. Ther. 2013, 18, 95–103. [Google Scholar] [CrossRef]
- Yasin, B.; Pang, M.; Turner, J.S.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I.; Wagar, E.A. Evaluation of the inactivation of infectious Herpes simplex virus by host-defense peptides. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Isaacs Charles, E.; Jia Jun, H.; Xu, W. A Lipid-Peptide Microbicide Inactivates Herpes Simplex Virus. Antimicrob. Agents Chemother. 2004, 48, 3182–3184. [Google Scholar] [CrossRef][Green Version]
- Mousavi Maleki, M.S.; Rostamian, M.; Madanchi, H. Antimicrobial peptides and other peptide-like therapeutics as promising candidates to combat SARS-CoV-2. Expert Rev. Anti-Infect. Ther. 2021, 19, 1205–1217. [Google Scholar] [CrossRef]
- Kasozi, K.I.; Niedbała, G.; Alqarni, M.; Zirintunda, G.; Ssempijja, F.; Musinguzi, S.P.; Usman, I.M.; Matama, K.; Hetta, H.F.; Mbiydzenyuy, N.E.; et al. Bee Venom-A Potential Complementary Medicine Candidate for SARS-CoV-2 Infections. Front. Public Health 2020, 8, 755. [Google Scholar] [CrossRef]
- Yang, W.; Hu, F.-L.; Xu, X.-F. Bee venom and SARS-CoV-2. Toxicon 2020, 181, 69–70. [Google Scholar] [CrossRef]
- Kwon, Y.B.; Lee, J.D.; Lee, H.J.; Han, H.J.; Mar, W.C.; Kang, S.K.; Beitz, A.J.; Lee, J.H. Bee venom injection into an acupuncture point reduces arthritis associated edema and nociceptive responses. Pain 2001, 90, 271–280. [Google Scholar] [CrossRef]
- Saini, S.S.; Peterson, J.W.; Chopra, A.K. Melittin binds to secretory phospholipase A2 and inhibits its enzymatic activity. Biochem. Biophys. Res. Commun. 1997, 238, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Son, D.J.; Lee, C.W.; Choi, M.S.; Lee, U.S.; Song, H.S.; Lee, J.M.; Hong, J.T. Melittin inhibits inflammatory target gene expression and mediator generation via interaction with IkappaB kinase. Biochem. Pharmacol. 2007, 73, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.R.; Kim, K.H.; An, H.J.; Kim, J.Y.; Chang, Y.C.; Chung, H.; Park, Y.Y.; Lee, M.L.; Park, K.K. The protective effects of melittin on Propionibacterium acnes-induced inflammatory responses in vitro and in vivo. J. Investig. Dermatol. 2014, 134, 1922–1930. [Google Scholar] [CrossRef]
- Han, S.M.; Kim, J.M.; Park, K.K.; Chang, Y.C.; Pak, S.C. Neuroprotective effects of melittin on hydrogen peroxide-induced apoptotic cell death in neuroblastoma SH-SY5Y cells. BMC Complement. Altern. Med. 2014, 14, 286. [Google Scholar] [CrossRef]
- Son, D.J.; Kang, J.; Kim, T.J.; Song, H.S.; Sung, K.J.; Yun, D.Y.; Hong, J.T. Melittin, a major bioactive component of bee venom toxin, inhibits PDGF receptor beta-tyrosine phosphorylation and downstream intracellular signal transduction in rat aortic vascular smooth muscle cells. J. Toxicol. Environ. Health A 2007, 70, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, H.J.; Choi, M.S.; Son, D.J.; Song, H.S.; Song, M.J.; Lee, J.M.; Han, S.B.; Kim, Y.; Hong, J.T. JNK pathway is involved in the inhibition of inflammatory target gene expression and NF-kappaB activation by melittin. J. Inflamm. (Lond.) 2008, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, S.H.; Son, D.J.; Oh, K.W.; Kim, K.H.; Song, H.S.; Kim, G.J.; Oh, G.T.; Yoon, D.Y.; Hong, J.T. Antiarthritic effect of bee venom: Inhibition of inflammation mediator generation by suppression of NF-kappaB through interaction with the p50 subunit. Arthritis Rheum. 2004, 50, 3504–3515. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.E.; Hwang, C.J.; Gu, S.M.; Park, M.H.; Kim, J.H.; Park, J.H.; Ahn, Y.J.; Kim, J.Y.; Song, M.J.; Song, H.S.; et al. Cancer Cell Growth Inhibitory Effect of Bee Venom via Increase of Death Receptor 3 Expression and Inactivation of NF-kappa B in NSCLC Cells. Toxins 2014, 6, 2210–2228. [Google Scholar] [CrossRef] [PubMed]
- Rajabnejad, S.H.; Mokhtarzadeh, A.; Abnous, K.; Taghdisi, S.M.; Ramezani, M.; Razavi, B.M. Targeted delivery of melittin to cancer cells by AS1411 anti-nucleolin aptamer. Drug Dev. Ind. Pharm. 2018, 44, 982–987. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, B.; Cheng, Y.; Yang, Y.; Huang, C.; Meng, X.; Wu, B.; Zhang, L.; Lv, X.; Li, J. Melittin induces PTCH1 expression by down-regulating MeCP2 in human hepatocellular carcinoma SMMC-7721 cells. Toxicol. Appl. Pharmacol. 2015, 288, 74–83. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, B.; Huang, C.; Meng, X.M.; Bian, E.B.; Li, J. Melittin restores PTEN expression by down-regulating HDAC2 in human hepatocelluar carcinoma HepG2 cells. PLoS ONE 2014, 9, e95520. [Google Scholar] [CrossRef]
- Wang, C.; Chen, T.; Zhang, N.; Yang, M.; Li, B.; Lü, X.; Cao, X.; Ling, C. Melittin, a major component of bee venom, sensitizes human hepatocellular carcinoma cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis by activating CaMKII-TAK1-JNK/p38 and inhibiting IkappaBalpha kinase-NFkappaB. J. Biol. Chem. 2009, 284, 3804–3813. [Google Scholar] [CrossRef]
- Jeong, Y.J.; Choi, Y.; Shin, J.M.; Cho, H.J.; Kang, J.H.; Park, K.K.; Choe, J.Y.; Bae, Y.S.; Han, S.M.; Kim, C.H.; et al. Melittin suppresses EGF-induced cell motility and invasion by inhibiting PI3K/Akt/mTOR signaling pathway in breast cancer cells. Food Chem. Toxicol. 2014, 68, 218–225. [Google Scholar] [CrossRef]
- Duffy, C.; Sorolla, A.; Wang, E.; Golden, E.; Woodward, E.; Davern, K.; Ho, D.; Johnstone, E.; Pfleger, K.; Redfern, A.; et al. Honeybee venom and melittin suppress growth factor receptor activation in HER2-enriched and triple-negative breast cancer. NPJ Precis. Oncol. 2020, 4, 24. [Google Scholar] [CrossRef]
- Park, M.H.; Choi, M.S.; Kwak, D.H.; Oh, K.-W.; Yoon, D.Y.; Han, S.B.; Song, H.S.; Song, M.J.; Hong, J.T. Anti-cancer effect of bee venom in prostate cancer cells through activation of caspase pathway via inactivation of NF-κB. Prostate 2011, 71, 801–812. [Google Scholar] [CrossRef]
- Ceremuga, M.; Stela, M.; Janik, E.; Gorniak, L.; Synowiec, E.; Sliwinski, T.; Sitarek, P.; Saluk-Bijak, J.; Bijak, M. Melittin-A Natural Peptide from Bee Venom Which Induces Apoptosis in Human Leukaemia Cells. Biomolecules 2020, 10, 247. [Google Scholar] [CrossRef]
- Jang, M.-H.; Shin, M.-C.; Lim, S.; Han, S.-M.; Park, H.-J.; Shin, I.; Lee, J.-S.; Kim, K.-A.; Kim, E.-H.; Kim, C.-J. Bee Venom Induces Apoptosis and Inhibits Expression of Cyclooxygenase-2 mRNA in Human Lung Cancer Cell Line NCI-H1299. J. Pharmacol. Sci. 2003, 91, 95–104. [Google Scholar] [CrossRef]
- Sharma, S.V. Melittin-induced hyperactivation of phospholipase A2 activity and calcium influx in ras-transformed cells. Oncogene 1993, 8, 939–947. [Google Scholar] [PubMed]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.-W. NF-κB in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef]
- Liu, S.; Yu, M.; He, Y.; Xiao, L.; Wang, F.; Song, C.; Sun, S.; Ling, C.; Xu, Z. Melittin prevents liver cancer cell metastasis through inhibition of the Rac1-dependent pathway. Hepatology 2008, 47, 1964–1973. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.-J.; Cho, H.-J.; Whang, K.; Lee, I.-S.; Park, K.-K.; Choe, J.-Y.; Han, S.-M.; Kim, C.-H.; Chang, H.-W.; Moon, S.-K.; et al. Melittin has an inhibitory effect on TNF-α-induced migration of human aortic smooth muscle cells by blocking the MMP-9 expression. Food Chem. Toxicol. 2012, 50, 3996–4002. [Google Scholar] [CrossRef]
- Park, J.H.; Jeong, Y.-J.; Park, K.-K.; Cho, H.-J.; Chung, I.-K.; Min, K.-S.; Kim, M.; Lee, K.-G.; Yeo, J.-H.; Park, K.-K.; et al. Melittin suppresses PMA-induced tumor cell invasion by inhibiting NF-κB and AP-1-dependent MMP-9 expression. Mol. Cells 2010, 29, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.K.; Ahmad, A.; Asthana, N.; Azmi, S.; Srivastava, R.M.; Srivastava, S.; Verma, R.; Vishwakarma, A.L.; Ghosh, J.K. Cell-Selective Lysis by Novel Analogues of Melittin against Human Red Blood Cells and Escherichia coli. Biochemistry 2010, 49, 7920–7929. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jang, A.Y.; Lin, S.; Lim, S.; Kim, D.; Park, K.; Han, S.M.; Yeo, J.H.; Seo, H.S. Melittin, a honeybee venomderived antimicrobial peptide, may target methicillinresistant Staphylococcus aureus. Mol. Med. Rep. 2015, 12, 6483–6490. [Google Scholar] [CrossRef]
- Rauen, H.M.; Schriewer, H.; Ferié, F. Alkylans-alkylandum reactions. 10. Anti-alkylanting effect of bee poison, melittine and apamine. Arzneimittelforschung 1972, 22, 1921–1922. [Google Scholar]
- Wolfgang Bücherl, E.E.B. Venomous Animals and Their Venoms: Venomous Vertebrates; Academic Press: Cambridge, MA, USA, 1971; Volume 3, pp. 157–158. [Google Scholar]
- Askari, P.; Namaei, M.H.; Ghazvini, K.; Hosseini, M. In vitro and in vivo toxicity and antibacterial efficacy of melittin against clinical extensively drug-resistant bacteria. BMC Pharmacol. Toxicol. 2021, 22, 42. [Google Scholar] [CrossRef]
- Park, J.H.; Yim, B.K.; Lee, J.-H.; Lee, S.; Kim, T.-H. Risk Associated with Bee Venom Therapy: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0126971. [Google Scholar] [CrossRef] [PubMed]
- Korošec, P.; Žiberna, K.; Šilar, M.; Dežman, M.; Čelesnik Smodiš, N.; Rijavec, M.; Kopač, P.; Eržen, R.; Lalek, N.; Bajrović, N.; et al. Immunological and clinical factors associated with adverse systemic reactions during the build-up phase of honeybee venom immunotherapy. Clin. Exp. Allergy 2015, 45, 1579–1589. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Soman, N.R.; Schlesinger, P.H.; Lanza, G.M.; Wickline, S.A. Cytolytic peptide nanoparticles (’NanoBees’) for cancer therapy. Wiley Interdiscip Rev. Nanomed Nanobiotechnol. 2011, 3, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Maher, S.; McClean, S. Melittin exhibits necrotic cytotoxicity in gastrointestinal cells which is attenuated by cholesterol. Biochem. Pharmacol. 2008, 75, 1104–1114. [Google Scholar] [CrossRef]
- Resende, V.M.F.; Vasilj, A.; Santos, K.S.; Palma, M.S.; Shevchenko, A. Proteome and phosphoproteome of Africanized and European honeybee venoms. Proteomics 2013, 13, 2638–2648. [Google Scholar] [CrossRef]
- Jallouk, A.P.; Palekar, R.U.; Marsh, J.N.; Pan, H.; Pham, C.T.N.; Schlesinger, P.H.; Wickline, S.A. Delivery of a Protease-Activated Cytolytic Peptide Prodrug by Perfluorocarbon Nanoparticles. Bioconjug. Chem. 2015, 26, 1640–1650. [Google Scholar] [CrossRef]
- Hou, K.K.; Pan, H.; Lanza, G.M.; Wickline, S.A. Melittin derived peptides for nanoparticle based siRNA transfection. Biomaterials 2013, 34, 3110–3119. [Google Scholar] [CrossRef]
- Hewish, D.R.; Barnham, K.J.; Werkmeister, J.A.; Kirkpatrick, A.; Bartone, N.; Liu, S.T.; Norton, R.S.; Curtain, C.; Rivetta, D.E. Structure and activity of D-Pro14 melittin. J. Protein. Chem. 2002, 21, 243–253. [Google Scholar] [CrossRef]
- Asthana, N.; Yadav, S.P.; Ghosh, J.K. Dissection of antibacterial and toxic activity of melittin: A leucine zipper motif plays a crucial role in determining its hemolytic activity but not antibacterial activity. J. Biol. Chem. 2004, 279, 55042–55050. [Google Scholar] [CrossRef]
- Rex, S. A Pro→Ala substitution in melittin affects self-association, membrane binding and pore-formation kinetics due to changes in structural and electrostatic properties. Biophys. Chem. 2000, 85, 209–228. [Google Scholar] [CrossRef]
- Guha, S.; Ferrie, R.P.; Ghimire, J.; Ventura, C.R.; Wu, E.; Sun, L.; Kim, S.Y.; Wiedman, G.R.; Hristova, K.; Wimley, W.C. Applications and evolution of melittin, the quintessential membrane active peptide. Biochem. Pharmacol. 2021, 193, 114769. [Google Scholar] [CrossRef]
- Liu, M.; Wang, H.; Liu, L.; Wang, B.; Sun, G. Melittin-MIL-2 fusion protein as a candidate for cancer immunotherapy. J. Transl. Med. 2016, 14, 155. [Google Scholar] [CrossRef] [PubMed]
- Wiedman, G.; Kim, S.Y.; Zapata-Mercado, E.; Wimley, W.C.; Hristova, K. pH-Triggered, Macromolecule-Sized Poration of Lipid Bilayers by Synthetically Evolved Peptides. J. Am. Chem. Soc. 2017, 139, 937–945. [Google Scholar] [CrossRef]
- Krauson, A.J.; He, J.; Wimley, W.C. Gain-of-function analogues of the pore-forming peptide melittin selected by orthogonal high-throughput screening. J. Am. Chem. Soc. 2012, 134, 12732–12741. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wu, W.; Sun, D.; Dai, H.B.; Wang, Y.; Zhong, Y.; Wang, J.X.; Maruf, A.; Nurhidayah, D.; Zhang, X.J.; et al. Acid-Activated Melittin for Targeted and Safe Antitumor Therapy. Bioconjug. Chem. 2018, 29, 2936–2944. [Google Scholar] [CrossRef]
- Shin, S.Y.; Lee, M.K.; Kim, K.L.; Hahm, K.S. Structure-antitumor and hemolytic activity relationships of synthetic peptides derived from cecropin A-magainin 2 and cecropin A-melittin hybrid peptides. J. Pept. Res. 1997, 50, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jin, H.; Qian, Y.; Qi, S.; Luo, H.; Luo, Q.; Zhang, Z. Hybrid melittin cytolytic Peptide-driven ultrasmall lipid nanoparticles block melanoma growth in vivo. ACS Nano 2013, 7, 5791–5800. [Google Scholar] [CrossRef]
- Lu, X.; Liu, J.; Gou, L.; Li, J.; Yuan, B.; Yang, K.; Ma, Y. Designing Melittin-Graphene Hybrid Complexes for Enhanced Antibacterial Activity. Adv. Healthc. Mater. 2019, 8, 1801521. [Google Scholar] [CrossRef]
- Dang, Y.-Q.; Li, H.-W.; Wu, Y. Construction of a Supramolecular Förster Resonance Energy Transfer System and Its Application Based on the Interaction between Cy3-Labeled Melittin and Phosphocholine Encapsulated Quantum Dots. ACS Appl. Mater. Interfaces 2012, 4, 1267–1272. [Google Scholar] [CrossRef] [PubMed]
- Hematyar, M.; Soleimani, M.; Es-Haghi, A.; Rezaei Mokarram, A. Synergistic co-delivery of doxorubicin and melittin using functionalized magnetic nanoparticles for cancer treatment: Loading and in vitro release study by LC-MS/MS. Artif. Cells Nanomed. Biotechnol. 2018, 46, S1226–S1235. [Google Scholar] [CrossRef]
- Soman, N.R.; Lanza, G.M.; Heuser, J.M.; Schlesinger, P.H.; Wickline, S.A. Synthesis and Characterization of Stable Fluorocarbon Nanostructures as Drug Delivery Vehicles for Cytolytic Peptides. Nano Lett. 2008, 8, 1131–1136. [Google Scholar] [CrossRef]
- Soman, N.R.; Baldwin, S.L.; Hu, G.; Marsh, J.N.; Lanza, G.M.; Heuser, J.E.; Arbeit, J.M.; Wickline, S.A.; Schlesinger, P.H. Molecularly targeted nanocarriers deliver the cytolytic peptide melittin specifically to tumor cells in mice, reducing tumor growth. J. Clin. Investig. 2009, 119, 2830–2842. [Google Scholar] [CrossRef] [PubMed]
- Jeong, I.; Kim, B.-S.; Lee, H.; Lee, K.-M.; Shim, I.; Kang, S.-K.; Yin, C.-S.; Hahm, D.-H. Prolonged analgesic effect of PLGA-encapsulated bee venom on formalin-induced pain in rats. Int. J. Pharm. 2009, 380, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cui, F.; Shi, K.; Cun, D.; Wang, R. Design of high payload PLGA nanoparticles containing melittin/sodium dodecyl sulfate complex by the hydrophobic ion-pairing technique. Drug Dev. Ind. Pharm. 2009, 35, 959–968. [Google Scholar] [CrossRef]
- Gonzalez-Horta, A.; Matamoros-Acosta, A.; Chavez-Montes, A.; Castro-Rios, R.; Lara-Arias, J. Biodegradable nanoparticles loaded with tetrameric melittin: Preparation and membrane disruption evaluation. Gen. Physiol. Biophys. 2017, 36, 373–381. [Google Scholar] [CrossRef]
- Xu, Y.; Deng, M.; Zhang, H.; Tan, S.; Li, D.; Li, S.; Luo, L.; Liao, G.; Wang, Q.; Huang, J.; et al. Selection of Affinity Reagents to Neutralize the Hemolytic Toxicity of Melittin Based on a Self-Assembled Nanoparticle Library. ACS Appl. Mater. Interfaces 2020, 12, 16040–16049. [Google Scholar] [CrossRef]
- Zetterberg, M.M.; Reijmar, K.; Pränting, M.; Engström, Å.; Andersson, D.I.; Edwards, K. PEG-stabilized lipid disks as carriers for amphiphilic antimicrobial peptides. J. Control. Release 2011, 156, 323–328. [Google Scholar] [CrossRef]
- Gao, J.; Xie, C.; Zhang, M.; Wei, X.; Yan, Z.; Ren, Y.; Ying, M.; Lu, W. RGD-modified lipid disks as drug carriers for tumor targeted drug delivery. Nanoscale 2016, 8, 7209–7216. [Google Scholar] [CrossRef] [PubMed]
- Ahlgren, S.; Reijmar, K.; Edwards, K. Targeting lipodisks enable selective delivery of anticancer peptides to tumor cells. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2325–2328. [Google Scholar] [CrossRef]
- Wang, H.; Wang, S.; Wang, R.; Wang, X.; Jiang, K.; Xie, C.; Zhan, C.; Wang, H.; Lu, W. Co-delivery of paclitaxel and melittin by glycopeptide-modified lipodisks for synergistic anti-glioma therapy. Nanoscale 2019, 11, 13069–13077. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Dai, Y.; Zhao, Y.; Qi, S.; Liu, L.; Lu, L.; Luo, Q.; Zhang, Z. Melittin-lipid nanoparticles target to lymph nodes and elicit a systemic anti-tumor immune response. Nat. Commun. 2020, 11, 1110. [Google Scholar] [CrossRef] [PubMed]
- Barrajón-Catalán, E.; Menéndez-Gutiérrez, M.P.; Falco, A.; Carrato, A.; Saceda, M.; Micol, V. Selective death of human breast cancer cells by lytic immunoliposomes: Correlation with their HER2 expression level. Cancer Lett. 2010, 290, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Falco, A.; Barrajón-Catalán, E.; Menéndez-Gutiérrez, M.P.; Coll, J.; Micol, V.; Estepa, A. Melittin-loaded immunoliposomes against viral surface proteins, a new approach to antiviral therapy. Antivir. Res. 2013, 97, 218–221. [Google Scholar] [CrossRef]
- Mao, J.; Liu, S.; Ai, M.; Wang, Z.; Wang, D.; Li, X.; Hu, K.; Gao, X.; Yang, Y. A novel melittin nano-liposome exerted excellent anti-hepatocellular carcinoma efficacy with better biological safety. J. Hematol. Oncol. 2017, 10, 71. [Google Scholar] [CrossRef]
- Li, Y.; Ruan, S.; Wang, Z.; Feng, N.; Zhang, Y. Hyaluronic Acid Coating Reduces the Leakage of Melittin Encapsulated in Liposomes and Increases Targeted Delivery to Melanoma Cells. Pharmaceutics 2021, 13, 1235. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Zheng, Y.; Chen, Y.; Wei, X.; Shi, S.; Chen, Y.; Zhu, W.; Wang, A.; Yang, L.; Xu, Y.; et al. Stable Loading and Delivery of Melittin with Lipid-Coated Polymeric Nanoparticles for Effective Tumor Therapy with Negligible Systemic Toxicity. ACS Appl. Mater. Interfaces 2021, 13, 55902–55912. [Google Scholar] [CrossRef]
- Lai, H.; Chen, F.; Lu, M.; Stenzel, M.H.; Xiao, P. Polypeptide-Grafted Nanodiamonds for Controlled Release of Melittin to Treat Breast Cancer. ACS Macro Lett. 2017, 6, 796–801. [Google Scholar] [CrossRef]
- Lv, S.; Sylvestre, M.; Song, K.; Pun, S.H. Development of D-melittin polymeric nanoparticles for anti-cancer treatment. Biomaterials 2021, 277, 121076. [Google Scholar] [CrossRef]
- Jia, H.-R.; Zhu, Y.-X.; Liu, X.; Pan, G.-Y.; Gao, G.; Sun, W.; Zhang, X.; Jiang, Y.-W.; Wu, F.-G. Construction of Dually Responsive Nanotransformers with Nanosphere–Nanofiber–Nanosphere Transition for Overcoming the Size Paradox of Anticancer Nanodrugs. ACS Nano 2019, 13, 11781–11792. [Google Scholar] [CrossRef]
- Liu, H.; Hu, Y.; Sun, Y.; Wan, C.; Zhang, Z.; Dai, X.; Lin, Z.; He, Q.; Yang, Z.; Huang, P.; et al. Co-delivery of Bee Venom Melittin and a Photosensitizer with an Organic–Inorganic Hybrid Nanocarrier for Photodynamic Therapy and Immunotherapy. ACS Nano 2019, 13, 12638–12652. [Google Scholar] [CrossRef]
- Cheng, B.; Xu, P. Redox-Sensitive Nanocomplex for Targeted Delivery of Melittin. Toxins 2020, 12, 582. [Google Scholar] [CrossRef]
- Clapp, A.R.; Medintz, I.L.; Mauro, J.M.; Fisher, B.R.; Bawendi, M.G.; Mattoussi, H. Fluorescence Resonance Energy Transfer Between Quantum Dot Donors and Dye-Labeled Protein Acceptors. J. Am. Chem. Soc. 2004, 126, 301–310. [Google Scholar] [CrossRef]
- Li, X.; Vinothini, K.; Ramesh, T.; Rajan, M.; Ramu, A. Combined photodynamic-chemotherapy investigation of cancer cells using carbon quantum dot-based drug carrier system. Drug Deliv. 2020, 27, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Kim, Y.J.; Lee, J.W.; Lee, Y.; Lim, Y.B.; Chung, H.W. Cyto-/genotoxic effect of CdSe/ZnS quantum dots in human lung adenocarcinoma cells for potential photodynamic UV therapy applications. J. Nanosci. Nanotechnol. 2012, 12, 2160–2168. [Google Scholar] [CrossRef]
- Cai, X.; Luo, Y.; Zhang, W.; Du, D.; Lin, Y. pH-Sensitive ZnO Quantum Dots–Doxorubicin Nanoparticles for Lung Cancer Targeted Drug Delivery. ACS Appl. Mater. Interfaces 2016, 8, 22442–22450. [Google Scholar] [CrossRef] [PubMed]
- He, S.J.; Cao, J.; Li, Y.S.; Yang, J.C.; Zhou, M.; Qu, C.Y.; Zhang, Y.; Shen, F.; Chen, Y.; Li, M.M.; et al. CdSe/ZnS quantum dots induce photodynamic effects and cytotoxicity in pancreatic cancer cells. World J. Gastroenterol. 2016, 22, 5012–5022. [Google Scholar] [CrossRef] [PubMed]
- Maleki, H.; Rai, A.; Pinto, S.; Evangelista, M.; Cardoso, R.M.S.; Paulo, C.; Carvalheiro, T.; Paiva, A.; Imani, M.; Simchi, A.; et al. High Antimicrobial Activity and Low Human Cell Cytotoxicity of Core–Shell Magnetic Nanoparticles Functionalized with an Antimicrobial Peptide. ACS Appl. Mater. Interfaces 2016, 8, 11366–11378. [Google Scholar] [CrossRef]
- Vu, H.D.; Huynh, P.T.; Ryu, J.; Kang, U.R.; Youn, S.W.; Kim, H.; Ahn, H.J.; Park, K.; Hwang, S.K.; Chang, Y.C.; et al. Melittin-loaded Iron Oxide Nanoparticles Prevent Intracranial Arterial Dolichoectasia Development through Inhibition of Macrophage-mediated Inflammation. Int. J. Biol. Sci. 2021, 17, 3818–3836. [Google Scholar] [CrossRef]
- Rai, A.; Pinto, S.; Evangelista, M.B.; Gil, H.; Kallip, S.; Ferreira, M.G.; Ferreira, L. High-density antimicrobial peptide coating with broad activity and low cytotoxicity against human cells. Acta Biomater. 2016, 33, 64–77. [Google Scholar] [CrossRef]
- Brand, I.; Khairalla, B. Structural changes in the model of the outer cell membrane of Gram-negative bacteria interacting with melittin: An in situ spectroelectrochemical study. Faraday Discuss. 2021, 232, 68–85. [Google Scholar] [CrossRef]
- Chugh, H.; Sood, D.; Chandra, I.; Tomar, V.; Dhawan, G.; Chandra, R. Role of gold and silver nanoparticles in cancer nano-medicine. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Jallouk, A.P.; Moley, K.H.; Omurtag, K.; Hu, G.; Lanza, G.M.; Wickline, S.A.; Hood, J.L. Nanoparticle incorporation of melittin reduces sperm and vaginal epithelium cytotoxicity. PLoS ONE 2014, 9, e95411. [Google Scholar] [CrossRef]
- Cabral, H.; Matsumoto, Y.; Mizuno, K.; Chen, Q.; Murakami, M.; Kimura, M.; Terada, Y.; Kano, M.R.; Miyazono, K.; Uesaka, M.; et al. Accumulation of sub-100 nm polymeric micelles in poorly permeable tumours depends on size. Nat. Nanotechnol. 2011, 6, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Kim, J.H.; Jeon, J.W.; Park, J.K.; Lee, B.J.; Suh, G.H.; Cho, C.W. Preformulation Studies of Bee Venom for the Preparation of Bee Venom-Loaded PLGA Particles. Molecules 2015, 20, 15072–15083. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Cun, D.; Tao, A.; Yang, M.; Shi, K.; Zhao, M.; Guan, Y. Preparation and characterization of melittin-loaded poly (DL-lactic acid) or poly (DL-lactic-co-glycolic acid) microspheres made by the double emulsion method. J. Control. Release 2005, 107, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Zhan, W.; Wei, T.; Yu, Q.; Chen, H. Fabrication of Supramolecular Bioactive Surfaces via β-Cyclodextrin-Based Host–Guest Interactions. ACS Appl. Mater. Interfaces 2018, 10, 36585–36601. [Google Scholar] [CrossRef]
- Lundquist, A.; Wessman, P.; Rennie, A.R.; Edwards, K. Melittin–Lipid interaction: A comparative study using liposomes, micelles and bilayerdisks. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1778, 2210–2216. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, W.; Jin, H.; Lovell, J.F.; Yang, M.; Ding, L.; Chen, J.; Corbin, I.; Luo, Q.; Zheng, G. Biomimetic Nanocarrier for Direct Cytosolic Drug Delivery. Angew. Chem. Int. Ed. 2009, 48, 9171–9175. [Google Scholar] [CrossRef]
- Wessman, P.; Strömstedt, A.A.; Malmsten, M.; Edwards, K. Melittin-Lipid Bilayer Interactions and the Role of Cholesterol. Biophys. J. 2008, 95, 4324–4336. [Google Scholar] [CrossRef]
- Wessman, P.; Morin, M.; Reijmar, K.; Edwards, K. Effect of α-helical peptides on liposome structure: A comparative study of melittin and alamethicin. J. Colloid Interface Sci. 2010, 346, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013, 8, 102. [Google Scholar] [CrossRef]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef]
- Tian, J.L.; Ke, X.; Chen, Z.; Wang, C.J.; Zhang, Y.; Zhong, T.C. Melittin liposomes surface modified with poloxamer 188: In vitro characterization and in vivo evaluation. Pharmazie 2011, 66, 362–367. [Google Scholar]
- Esim, O.; Hascicek, C. Lipid-Coated Nanosized Drug Delivery Systems for an Effective Cancer Therapy. Curr. Drug Deliv. 2021, 18, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, N.A.N.; Quarta, A.; Di Corato, R.; Dini, L.; Nobile, C.; Tasco, V.; Carallo, S.; Cascione, M.; Malfettone, A.; Soukupova, J.; et al. Hybrid polymeric-protein nano-carriers (HPPNC) for targeted delivery of TGFβ inhibitors to hepatocellular carcinoma cells. J. Mater. Sci. Mater. Med. 2017, 28, 120. [Google Scholar] [CrossRef]
- Zylberberg, C.; Gaskill, K.; Pasley, S.; Matosevic, S. Engineering liposomal nanoparticles for targeted gene therapy. Gene Ther. 2017, 24, 441–452. [Google Scholar] [CrossRef]
- Erazo-Oliveras, A.; Muthukrishnan, N.; Baker, R.; Wang, T.-Y.; Pellois, J.-P. Improving the Endosomal Escape of Cell-Penetrating Peptides and Their Cargos: Strategies and Challenges. Pharmaceuticals 2012, 5, 1177–1209. [Google Scholar] [CrossRef] [PubMed]
- Paray, B.A.; Ahmad, A.; Khan, J.M.; Taufiq, F.; Pathan, A.; Malik, A.; Ahmed, M.Z. The role of the multifunctional antimicrobial peptide melittin in gene delivery. Drug Discov. Today 2021, 26, 1053–1059. [Google Scholar] [CrossRef]
- Ogris, M.; Carlisle, R.C.; Bettinger, T.; Seymour, L.W. Melittin Enables Efficient Vesicular Escape and Enhanced Nuclear Access of Nonviral Gene Delivery Vectors. J. Biol. Chem. 2001, 276, 47550–47555. [Google Scholar] [CrossRef]
- Overhoff, M.; Sczakiel, G. Phosphorothioate-stimulated uptake of short interfering RNA by human cells. EMBO Rep. 2005, 6, 1176–1181. [Google Scholar] [CrossRef]
- Boeckle, S.; Wagner, E.; Ogris, M. C- versus N-terminally linked melittin-polyethylenimine conjugates: The site of linkage strongly influences activity of DNA polyplexes. J. Gene Med. 2005, 7, 1335–1347. [Google Scholar] [CrossRef]
- Zhang, W.; Song, J.; Liang, R.; Zheng, X.; Chen, J.; Li, G.; Zhang, B.; Yan, X.; Wang, R. Stearylated Antimicrobial Peptide Melittin and Its Retro Isomer for Efficient Gene Transfection. Bioconjug. Chem. 2013, 24, 1805–1812. [Google Scholar] [CrossRef]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Oude Blenke, E.; Sleszynska, M.; Evers, M.J.W.; Storm, G.; Martin, N.I.; Mastrobattista, E. Strategies for the Activation and Release of the Membranolytic Peptide Melittin from Liposomes Using Endosomal pH as a Trigger. Bioconjug. Chem. 2017, 28, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Dohmen, C.; Philipp, A.; Kiener, D.; Maiwald, G.; Scheu, C.; Ogris, M.; Wagner, E. Synthesis and Biological Evaluation of a Bioresponsive and Endosomolytic siRNA−Polymer Conjugate. Mol. Pharm. 2009, 6, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Keil, T.W.M.; Baldassi, D.; Merkel, O.M. T-cell targeted pulmonary siRNA delivery for the treatment of asthma. Wiley Interdiscip. Reviews. Nanomed. Nanobiotechnol. 2020, 12, e1634. [Google Scholar] [CrossRef]
- Meyer, M.; Zintchenko, A.; Ogris, M.; Wagner, E. A dimethylmaleic acid-melittin-polylysine conjugate with reduced toxicity, pH-triggered endosomolytic activity and enhanced gene transfer potential. J. Gene. Med. 2007, 9, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Yumul, R.C.; Pun, S.H. Virus-Inspired Polymer for Efficient In Vitro and In Vivo Gene Delivery. Angew. Chem. (Int. Ed. Engl. ) 2016, 55, 12013–12017. [Google Scholar] [CrossRef]
- Feldmann, D.P.; Cheng, Y.; Kandil, R.; Xie, Y.; Mohammadi, M.; Harz, H.; Sharma, A.; Peeler, D.J.; Moszczynska, A.; Leonhardt, H.; et al. In vitro and in vivo delivery of siRNA via VIPER polymer system to lung cells. J. Control. Release 2018, 276, 50–58. [Google Scholar] [CrossRef]
- Zhang, S.-K.; Song, J.-W.; Li, S.-B.; Gao, H.-W.; Chang, H.-Y.; Jia, L.-l.; Gong, F.; Tan, Y.-X.; Ji, S.-P. Design of pH-sensitive peptides from natural antimicrobial peptides for enhancing polyethylenimine-mediated gene transfection. J. Gene Med. 2017, 19, e2955. [Google Scholar] [CrossRef]
- Ahmad, A.; Ranjan, S.; Zhang, W.; Zou, J.; Pyykkö, I.; Kinnunen, P.K.J. Novel endosomolytic peptides for enhancing gene delivery in nanoparticles. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1848, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Ventura, C.R.; Wiedman, G.R. Substituting azobenzene for proline in melittin to create photomelittin: A light-controlled membrane active peptide. Biochim. Biophys. Acta (BBA)-Biomembr. 2021, 1863, 183759. [Google Scholar] [CrossRef]
- Chen, C.-P.; Kim, J.-s.; Steenblock, E.; Liu, D.; Rice, K.G. Gene transfer with poly-melittin peptides. Bioconjug. Chem. 2006, 17, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.K.; Pan, H.; Ratner, L.; Schlesinger, P.H.; Wickline, S.A. Mechanisms of Nanoparticle-Mediated siRNA Transfection by Melittin-Derived Peptides. ACS Nano 2013, 7, 8605–8615. [Google Scholar] [CrossRef]
- Pan, H.; Myerson, J.W.; Ivashyna, O.; Soman, N.R.; Marsh, J.N.; Hood, J.L.; Lanza, G.M.; Schlesinger, P.H.; Wickline, S.A. Lipid membrane editing with peptide cargo linkers in cells and synthetic nanostructures. FASEB J. 2010, 24, 2928–2937. [Google Scholar] [CrossRef]
- Jafari, E.; Mahmoodi, S. Design, expression, and purification of a multi-epitope vaccine against Helicobacter Pylori based on Melittin as an adjuvant. Microb. Pathog. 2021, 157, 104970. [Google Scholar] [CrossRef]
- Bramwell, V.W.; Somavarapu, S.; Outschoorn, I.; Alpar, H.O. Adjuvant Action of Melittin Following Intranasal Immunisation with Tetanus and Diphtheria Toxoids. J. Drug Target. 2003, 11, 525–530. [Google Scholar] [CrossRef] [PubMed]


| Type of Microbial | Treatment or Method | Result | Reference | |
|---|---|---|---|---|
| Virus | HIV-1 | MEL | ID50 values was in the range 0.9–1.5 μM | [17] |
| HSV-1 and HSV-2 | MEL | CC50 ranges 1.35–2.05 μM | [18] | |
| SARS-CoV-2 | Sitagliptin-MEL nano-conjugate | IC50 values 8.439 μM | [19] | |
| Bacteria | Pseudomonas aeruginosa | MEL | MIC 10 µg/mL and MBC 20 µg/mL | [20] |
| Methicillin-resistant Staphylococcus aureus | MEL | MIC 6.7 μg/mL and MBC 26 μg/mL. | [21] | |
| Multidrug-resistant Acinetobacter baumannii | MEL | MIC ranges 0.50–32 μg/mL | [22] | |
| E. coli and Staphylococcus aureus | MEL and ionic liquids combination | E. coli: MIC value was 0.52 μM MEL with 10 μM [Pyr C12]Br− S. aureus: MIC value was 0.62 μM MEL with 20 μM [Pyr C10] Br−. | [23] | |
| Multidrug-resistant Acinetobacter baumannii and Pseudomonas aeruginosa | Combination of MEL and conventional antibiotics | MDR A. baumannii isolates: MIC for MEL and doripenem were reduced by 61.5 and 51.5 folds, respectively. MDR P. aeruginosa isolates: MIC was reduced by 63.5 and 58 folds for MEL–doripenem, respectively, and by 16 and 11 folds for MEL–ceftazidime, respectively. | [24] | |
| Fungus | Aspergillus flavus, Aspergillus fumigatus, and Aspergillus parasiticus | MEL | MIC values was 1.25 μM, 1.25 μM, and 2.5 μM for Aspergillus flavus, Aspergillus fumigatus, and Aspergillus parasiticus strains respectively. | [25] |
| Candida albicans | MEL | MIC values for different strains of Candida albicans ranges from 8 μM to 32 μM. | [26] | |
| Type | Loading Strategy | Size | Applications | Reference |
|---|---|---|---|---|
| Quantum dots | MEL was modified to CdSe/ZnS core quantum dots | 5–10 nm | Quantum dots were used to study the interaction between protein and membrane, and had potential to deliver MEL in vivo. | [83] |
| Iron oxide nanoparticles | MEL and doxorubicin (DOX) were co-loaded to citric acid-functionalized Fe3O4 magnetic nanoparticles (CA-MNPs) | 20 nm | The release of both MEL and DOX was strongly enhanced at pH 4.5 and the nanoparticles were potentially applied in magnetically targeted cancer therapy. | [84] |
| Perfluorocarbon (PFC) nanoparticles | MEL was added to the PFC nanoparticles | ~290 nm | PFC nanoparticles retained their structural integrity after the addition and contribute to the stability and slow dissociation of MEL from the stabilizing monolayer. | [85] |
| MEL was mixed and incubated with PFC nanoparticles | 227 nm | The growth of the tumors was inhibited by 24.68% in MDA-MB-435 human breast cancer. | [86] | |
| MEL derivative peptide was incubated with PFC nanoparticles composed of egg phosphatidylcholineand dipalmitoylphosphatidylglycerol | ~280 nm | This MEL derivative is activated by matrix metalloproteinase-9 (MMP-9), a protease overexpressed in many tumor cells. In addition, treatment of PFC nanoparticles resulted in ~54% reduction in melanoma tumor size in vivo. | [70] | |
| Poly (d,l-lactic acid-coglycolic acid) (PLGA) nanoparticles | BV-loaded PLGA/PVA nanoparticles | 180 nm | PLGA nanoparticles reduced side effects by slowing down BV release, and prolonged suppression of nociceptive behavior in rats with formalin-induced pain. | [87] |
| MEL was modified with sodium dodecyl sulfate and then formulated into PLGA nanoparticles | ~130 nm | MEL was loaded with a high encapsulation efficiency in the nanoparticles and the concentration of half the cell growth (GI50) in breast cancer MCF-7 cells was 4.42 μg/mL in vitro. | [88] | |
| Tetrameric MEL binds avidly to PLGA-NPs | 110 nm | Biodegradable tetrameric MEL is encapsulated in nanoparticles at efficiency of 97% and retains lytic activity. | [89] | |
| β-cyclodextrin(β-CDP) nanoparticles | 5 different functional monomer adamantane derivatives (Ad-Ds) incubated with β-CDPs respectively, and then mixed with MEL | 30–200 nm | The percentage of hemolytic toxicity neutralization reached 100% at the concentration of 100 μM. The cytotoxicity of 30 μg/mL MEL with 2 mmol/mL nanoparticle decreased by sixfold compared with that of free MEL in CCRF-CEM cells. | [90] |
| Lipodisks | MEL incubated with PEG-stabilized lipid disks which composed of POPC/cholesterol/ceramide-PEG5000 | 20–100 nm | PEGylated lipodisks allowed stable loading of MEL, and retained anti-bacterial activity of MEL in E. coli, but extended the actions by slowing down releasing rate. | [91] |
| Lipid disks was modified by c(RGDyK)-PEG3400-DSPE | 50 nm | The disks induced no hemoglobin release at maximum tested concentration (100 μg/mL) and presented significate targeting and in vivo anti-tumor effect towards U87 glioma cells. | [92] | |
| MEL loaded lipodisks contained EGF-conjugated PEG-lipids. | ~20 nm | The EGF-targeted lipodisks binded specifically to A-431 tumor cells, and resulted in a improved cell-killing effect, as cell viability decreased 20% compared to free MEL. | [93] | |
| MEL and paclitaxel were co-loaded within 9G-A7R modified lipodisks. | ~50 nm | Co-loading prevented leakage of MEL from the disks and improved cytotoxicity on U87 cells in vitro and anti-tumor effect in intracranial glioma models. The synergistic effect of MEL and paclitaxel was proved as combination index values was 0.45. | [94] | |
| Lipid nanoparticles | MEL was linked to an amphipathic peptide then loaded in ultrasmall lipid nanoparticles | 14 nm | The ultrasmall lipid nanoparticles significantly reduced the hemolysis of MEL and showed obvious anti-tumor effect in malignant melanoma B16F10 cells, with IC50 values being 11.26 μM. | [81] |
| MEL-lipid conjugate nanoparticles | MEL-phospholipid scaffold | 10–20 nm | The nanoparticles induced tumor cell apoptosis, releasing whole-tumor antigens in situ, and targeting to lymph nodes. | [95] |
| Liposomes | MEL was loaded in PEGylated anti-HER2 immunoliposomes modified by the complete antibody (trastuzumab) | 139 nm | The immunoliposomes decreased cancer cells viability in a dose–response manner and in correlation to the level of HER2 expression in human breast cancer cells. | [96] |
| MEL loaded liposomes was modified by antibodies against the fish viral hemorrhagic septicemia rhabdovirus (VHSV) glycoprotein G (gpG) | ~140 nm | The in vitro antiviral studies showed that the liposomes inhibited the infectivity by 95.2% through inactivating VHSV. | [97] | |
| MEL was modified with 2% poloxamer 188 then loaded in nano-liposomes. | NA | Multiple hepatic carcinoma cell lines (Bel-7402, BMMC-7721, HepG2, LM-3, and Hepa 1–6 cells) were sensitive to the liposomes, and the IC50 value was close to free MEL, indicating efficient anti-tumor effect. | [98] | |
| Hyaluronic acid (HA) modified MEL-loading liposomes | 133 nm | HA enhanced the sustained-release effect of MEL from the liposomes and provide targeting ability via specific binding with CD44, which is highly expressed on the surface of melanoma B16F10 cells. | [99] | |
| Lipid-coated polymeric Nanoparticles | MEL and poly γ-glutamic acid (γ-PGA) formed nanoparticles which then coated by cationic liposomes modified by PEG and DSPE-PEG-RGD | ~100 nm | The hemolytic activity and nonspecific cytotoxicity of MEL were remarkably reduced by the lipid-coated polymeric nanoparticles and the RGD-modified RGD modified nanoparticles effectively induced apoptosis in A549 cells. | [100] |
| Stimulus-responsive delivery systems | MEL was grafted to nanodiamonds coated with PEGylated PGA. | 220 nm | The nanoparticles were pH sensitive and steady able to released MEL in an acidic environment. Toxicity to breast cancer MFC-7 cells was enhanced than free MEL in a concentration-dependent manner. | [101] |
| D-MEL was conjugated with PEG which is polymerized with DIPAMA and PDSEMA, to form micelles. | 33 nm | The pH sensitive micellar formulations unsheathes MEL only at endosomal pH, remarkably reducing hemolytic effects of MEL, and IC50 for the micelles in 3T3, A549, CT26 cancer cells were 8.5 μM, 6.9 μM, 11.6 μM, respectively. | [102] | |
| MEL was loaded in negatively charged nanospheres consisting of NIR-absorbing molecule cypate and HA. | ∼50 nm | The nanospheres responsive to both pH and near-infrared (NIR) laser irradiation changes into net-like nanofibers and small nanospheres (~25 nm) when stimulated and induce cancer cell death, inhibit the metastatic dissemination of tumor cells, and facilitated deep tumor penetration o | [103] | |
| Serum albumin (SA)-coated boehmite scaffold was loaded with photosensitizer chlorin e6 (Ce6) and MEL. | 184 nm | The nanocarrier exerted high encapsulation efficiency of MEL and low hemocompatibility. In vivo phototreatment of the scaffold eliminated 4T1 cells remarkably in subcutaneous breast tumor models. | [104] | |
| MEL loaded in redox-sensitive nanocomplexes | 357 nm | The nanocomplexes decreased hemolysis of MEL and released MEL responding to high redox potential environment, and showed an enhanced cytotoxicity on both HCT 116 colon cancer cells and MCF-7 breast cancer cells. | [105] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, A.; Zheng, Y.; Zhu, W.; Yang, L.; Yang, Y.; Peng, J. Melittin-Based Nano-Delivery Systems for Cancer Therapy. Biomolecules 2022, 12, 118. https://doi.org/10.3390/biom12010118
Wang A, Zheng Y, Zhu W, Yang L, Yang Y, Peng J. Melittin-Based Nano-Delivery Systems for Cancer Therapy. Biomolecules. 2022; 12(1):118. https://doi.org/10.3390/biom12010118
Chicago/Turabian StyleWang, Anqi, Yuan Zheng, Wanxin Zhu, Liuxin Yang, Yang Yang, and Jinliang Peng. 2022. "Melittin-Based Nano-Delivery Systems for Cancer Therapy" Biomolecules 12, no. 1: 118. https://doi.org/10.3390/biom12010118
APA StyleWang, A., Zheng, Y., Zhu, W., Yang, L., Yang, Y., & Peng, J. (2022). Melittin-Based Nano-Delivery Systems for Cancer Therapy. Biomolecules, 12(1), 118. https://doi.org/10.3390/biom12010118