
Lumican in Carcinogenesis—Revisited



 ,
, 

Abstract
:1. Introduction-Cancer and ECM
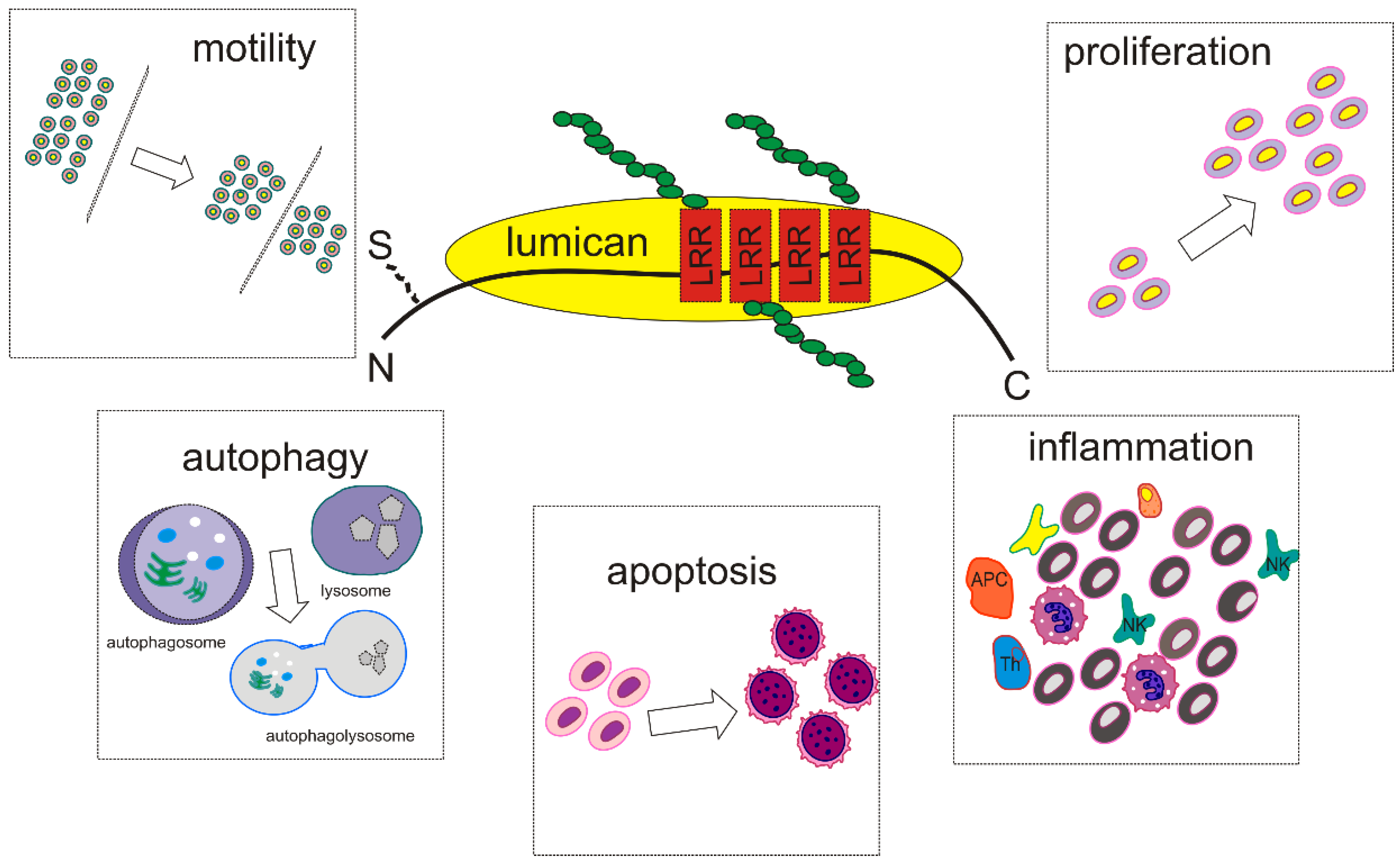
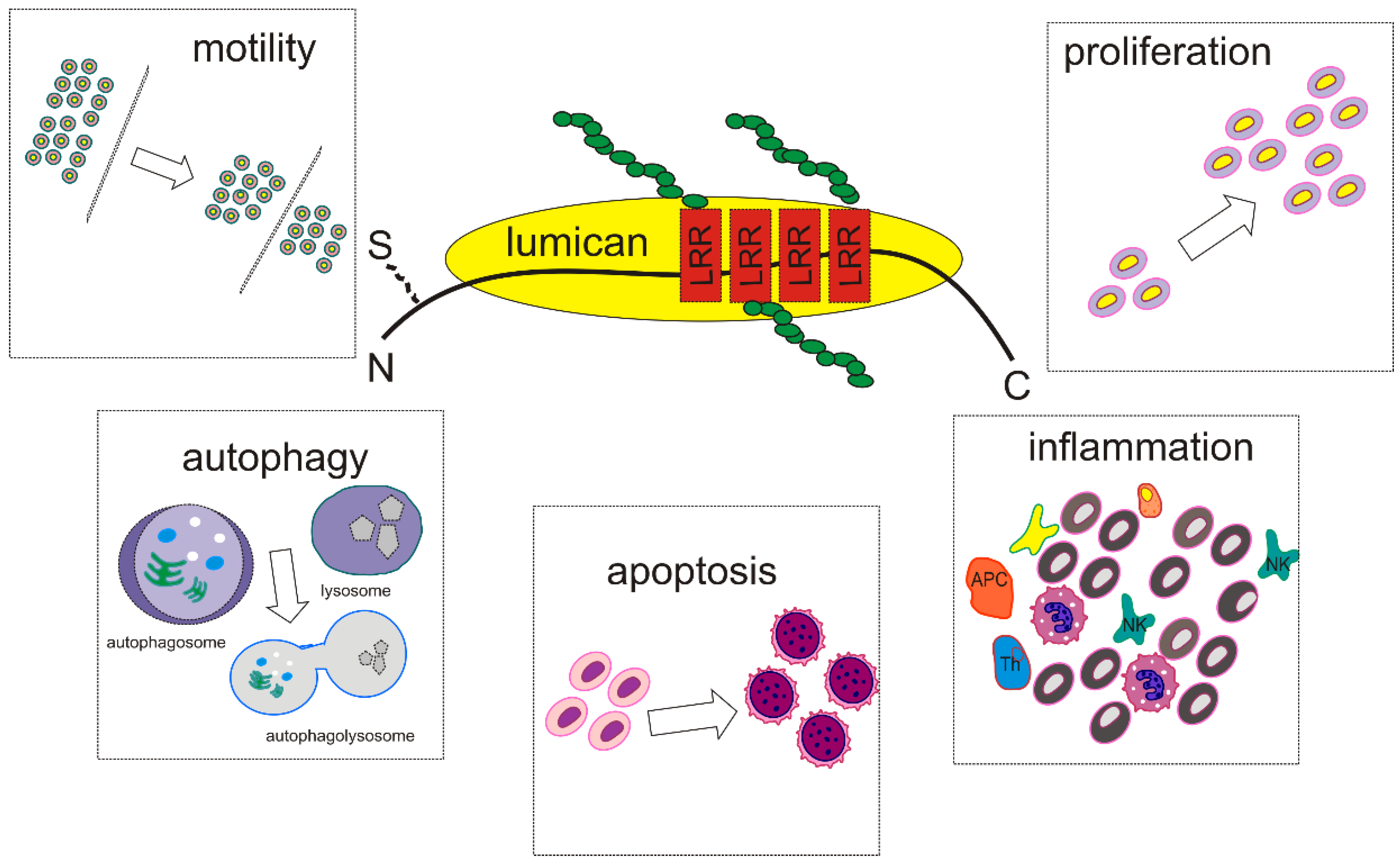
2. SLRPs Structure and Function—Focus on Lumican
3. Lumican Structure, Function, and Expression-Correlation with Carcinogenesis
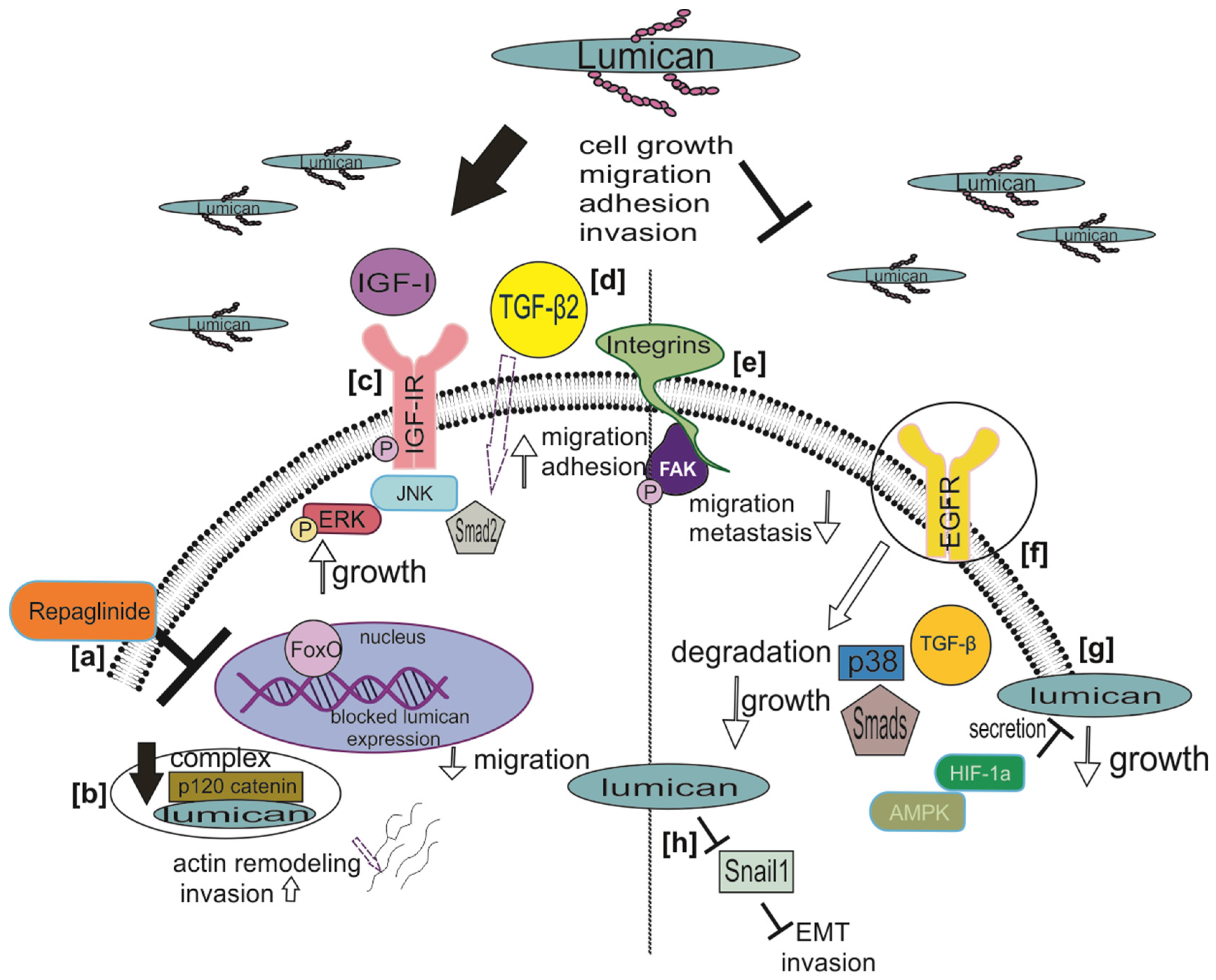
Lumican Regulates Cancer Cell Growth, Invasion, and Metastasis
4. Lumican Modulates Cancer Cell Motility
5. Lumican at the Crossroad between Apoptosis and Autophagy
6. Implications of Lumican in Cancer-Associated Inflammation
7. Lumican as Prognosis Marker, Chemoresistance Regulator, and Cancer Target
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tzanakakis, G.; Neagu, M.; Tsatsakis, A.; Nikitovic, D. Proteoglycans and Immunobiology of Cancer—Therapeutic Implications. Front. Immunol. 2019, 10, 875. [Google Scholar] [CrossRef] [PubMed]
- Tzanakakis, G.; Giatagana, E.-M.; Kuskov, A.; Berdiaki, A.; Tsatsakis, A.; Neagu, M.; Nikitovic, D. Proteoglycans in the Pathogenesis of Hormone-Dependent Cancers: Mediators and Effectors. Cancers 2020, 12, 2401. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, K.C.; Chu, P.Y.; Chang, G.C.; Liu, K.J. Elevated Expression of Lumican in Lung Cancer Cells Promotes Bone Metastasis through an Autocrine Regulatory Mechanism. Cancers 2020, 12, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 1–19. [Google Scholar] [CrossRef]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. The Extracellular Matrix: Not Just Pretty Fibrils. Science 2009, 326, 1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikitovic, D.; Etzardi, M.; Eberdiaki, A.; Tsatsakis, A.; Tzanakakis, G.N. Cancer Microenvironment and Inflammation: Role of Hyaluronan. Front. Immunol. 2015, 6, 169. [Google Scholar] [CrossRef] [Green Version]
- Nikitovic, D.; Berdiaki, A.; Spyridaki, I.; Krasanakis, T.; Tsatsakis, A.; Tzanakakis, G.N. Proteoglycans—Biomarkers and Targets in Cancer Therapy. Front. Endocrinol. 2018, 9, 69. [Google Scholar] [CrossRef] [Green Version]
- Tampa, M.; Georgescu, S.; Mitran, M.; Mitran, C.; Matei, C.; Caruntu, A.; Scheau, C.; Nicolae, I.; Matei, A.; Caruntu, C.; et al. Current Perspectives on the Role of Matrix Metalloproteinases in the Pathogenesis of Basal Cell Carcinoma. Biomolecules 2021, 11, 903. [Google Scholar] [CrossRef]
- Ahmad, R.; Eubank, T.; Lukomski, S.; Boone, B. Immune Cell Modulation of the Extracellular Matrix Contributes to the Pathogenesis of Pancreatic Cancer. Biomolecules 2021, 11, 901. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.Z.; Roden, D.L.; Wang, C.; Holliday, H.; Harvey, K.; Cazet, A.S.; Murphy, K.J.; Pereira, B.; Al-Eryani, G.; Bartonicek, N.; et al. Stromal cell diversity associated with immune evasion in human triple-negative breast cancer. EMBO J. 2020, 39, e104063. [Google Scholar] [CrossRef]
- Zeltz, C.; Primac, I.; Erusappan, P.; Alam, J.; Noel, A.; Gullberg, D. Cancer-associated fibroblasts in desmoplastic tumors: Emerging role of integrins. Semin. Cancer Biol. 2019, 62, 166–181. [Google Scholar] [CrossRef]
- Thomas, D.; Radhakrishnan, P. Tumor-stromal crosstalk in pancreatic cancer and tissue fibrosis. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Onyeisi, J.; Lopes, C.; Götte, M. Syndecan-4 as a Pathogenesis Factor and Therapeutic Target in Cancer. Biomolecules 2021, 11, 503. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Pearson, C.S.; Geller, H.M. Flexible Roles for Proteoglycan Sulfation and Receptor Signaling. Trends Neurosci. 2017, 41, 47–61. [Google Scholar] [CrossRef]
- Hayes, A.; Sugahara, K.; Farrugia, B.; Whitelock, J.M.; Caterson, B.; Melrose, J. Biodiversity of CS-proteoglycan sulphation motifs: Chemical messenger recognition modules with roles in information transfer, control of cellular behaviour and tissue morphogenesis. Biochem. J. 2018, 475, 587–620. [Google Scholar] [CrossRef] [Green Version]
- Couchman, J.R.; Pataki, C.A. An Introduction to Proteoglycans and Their Localization. J. Histochem. Cytochem. 2012, 60, 885–897. [Google Scholar] [CrossRef]
- Mytilinaiou, M.; Nikitovic, D.; Berdiaki, A.; Kostouras, A.; Papoutsidakis, A.; Tsatsakis, A.; Tzanakakis, G.N. Emerging roles of syndecan 2 in epithelial and mesenchymal cancer progression. IUBMB Life 2017, 69, 824–833. [Google Scholar] [CrossRef] [Green Version]
- McEwan, P.A.; Scott, P.G.; Bishop, P.; Bella, J. Structural correlations in the family of small leucine-rich repeat proteins and proteoglycans. J. Struct. Biol. 2006, 155, 294–305. [Google Scholar] [CrossRef]
- Kobe, B.; Kajava, A.V. The leucine-rich repeat as a protein recognition motif. Curr. Opin. Struct. Biol. 2001, 11, 725–732. [Google Scholar] [CrossRef]
- Schaefer, L.; Iozzo, R.V. Biological Functions of the Small Leucine-rich Proteoglycans: From Genetics to Signal Transduction. J. Biol. Chem. 2008, 283, 21305–21309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, S.; Li, J.; Ge, W. Multifaceted Roles of Asporin in Cancer: Current Understanding. Front. Oncol. 2019, 9, 948. [Google Scholar] [CrossRef] [PubMed]
- Sanders, E.J.; Walter, M.A.; Parker, E.; Ara’mburo, C.; Harvey, S. Opticin binds retinal growth hormone in the embryonic vitreous. Investig. Opthalmol. Vis. Sci. 2003, 44, 5404–5409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikitovic, D.; Papoutsidakis, A.; Karamanos, N.; Tzanakakis, G. Lumican affects tumor cell functions, tumor–ECM interactions, angiogenesis and inflammatory response. Matrix Biol. 2014, 35, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Kram, V.; Kilts, T.M.; Bhattacharyya, N.; Li, L.; Young, M.F. Small leucine rich proteoglycans, a novel link to osteoclastogenesis. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef]
- Tzanakakis, G.; Giatagana, E.-M.; Berdiaki, A.; Spyridaki, I.; Hida, K.; Neagu, M.; Tsatsakis, A.; Nikitovic, D. The Role of IGF/IGF-IR-Signaling and Extracellular Matrix Effectors in Bone Sarcoma Pathogenesis. Cancers 2021, 13, 2478. [Google Scholar] [CrossRef] [PubMed]
- Zafiropoulos, A.; Nikitovic, D.; Katonis, P.; Tsatsakis, A.; Karamanos, N.K.; Tzanakakis, G.N. Decorin-Induced Growth Inhibition Is Overcome through Protracted Expression and Activation of Epidermal Growth Factor Receptors in Osteosarcoma Cells. Mol. Cancer Res. 2008, 6, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Voudouri, K.; Nikitovic, D.; Berdiaki, A.; Kletsas, D.; Karamanos, N.; Tzanakakis, G.N. IGF-I/EGF and E2 signaling crosstalk through IGF-IR conduit point affects breast cancer cell adhesion. Matrix Biol. 2016, 56, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Aggelidakis, J.; Berdiaki, A.; Nikitovic, D.; Papoutsidakis, A.; Papachristou, D.J.; Tsatsakis, A.M.; Tzanakakis, G.N. Biglycan Regulates MG63 Osteosarcoma Cell Growth Through a LPR6/beta-Catenin/IGFR-IR Signaling Axis. Front. Oncol. 2018, 8, 470. [Google Scholar] [CrossRef] [Green Version]
- Papoutsidakis, A.; Giatagana, E.M.; Berdiaki, A.; Spyridaki, I.; Spandidos, D.A.; Tsatsakis, A.; Tzanakakis, G.N.; Nikitovic, D. Lumican mediates HTB94 chondrosarcoma cell growth via an IGFIR/Erk1/2 axis. Int. J. Oncol. 2020, 57, 791–803. [Google Scholar] [CrossRef]
- Chakravarti, S.; Stallings, R.L.; Sundarraj, N.; Cornuet, P.K.; Hassell, J.R. Primary Structure of Human Lumican (Keratan Sulfate Proteoglycan) and Localization of the Gene (LUM) to Chromosome 12q21.3–q22. Genomics 1995, 27, 481–488. [Google Scholar] [CrossRef]
- Dunlevy, J.R.; Neame, P.J.; Vergnes, J.-P.; Hassell, J.R. Identification of the N-Linked Oligosaccharide Sites in Chick Corneal Lumican and Keratocan That Receive Keratan Sulfate. J. Biol. Chem. 1998, 273, 9615–9621. [Google Scholar] [CrossRef] [Green Version]
- Roughley, P.J.; White, R.J.; Cs-Szabó, G.; Mort, J.S. Changes with age in the structure of fibromodulin in human articular cartilage. Osteoarthr. Cartil. 1996, 4, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Cornuet, P.K.; Blochberger, T.C.; Hassell, J.R. Molecular polymorphism of lumican during corneal development. Investig. Ophthalmol. Vis. Sci. 1994, 35, 870–877. [Google Scholar]
- Grover, J.; Chen, X.N.; Korenberg, J.R.; Roughley, P.J. The human lumican gene. Organization, chromosomal location, and expression in articular cartilage. J. Biol. Chem. 1995, 270, 21942–21949. [Google Scholar] [CrossRef] [Green Version]
- Chakravarti, S.; Magnuson, T.; Lass, J.H.; Jepsen, K.J.; LaMantia, C.; Carroll, H. Lumican Regulates Collagen Fibril Assembly: Skin Fragility and Corneal Opacity in the Absence of Lumican. J. Cell Biol. 1998, 141, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; Shiraishi, A.; Kao, C.W.-C.; Converse, R.L.; Funderburgh, J.L.; Swiergiel, J.; Roth, M.R.; Conrad, G.W.; Kao, W.W.-Y. Characterization and Expression of the Mouse Lumican Gene. J. Biol. Chem. 1997, 272, 30306–30313. [Google Scholar] [CrossRef] [Green Version]
- Raouf, A.; Ganss, B.; McMahon, C.; Vary, C.; Roughley, P.J.; Seth, A. Lumican is a major proteoglycan component of the bone matrix. Matrix Biol. 2002, 21, 361–367. [Google Scholar] [CrossRef]
- Quantock, A.J.; Meek, K.; Chakravarti, S. An x-ray diffraction investigation of corneal structure in lumican-deficient mice. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1750–1756. [Google Scholar]
- Chakravarti, S.; Petroll, W.M.; Hassell, J.R.; Jester, J.; Lass, J.H.; Paul, J.; Birk, D.E. Corneal opacity in lumican-null mice: Defects in collagen fibril structure and packing in the posterior stroma. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3365–3373. [Google Scholar]
- Mao, W.; Luo, M.; Huang, X.; Wang, Q.; Fan, J.; Gao, L.; Zhang, Y.; Geng, J. Knockdown of Lumican Inhibits Proliferation and Migration of Bladder Cancer. Transl. Oncol. 2019, 12, 1072–1078. [Google Scholar] [CrossRef]
- De Wit, M.; Carvalho, B.; Diemen, P.M.D.-V.; Van Alphen, C.; Beliën, J.A.M.; Meijer, G.A.; Fijneman, R.J.A. Lumican and versican protein expression are associated with colorectal adenoma-to-carcinoma progression. PLoS ONE 2017, 12, e0174768. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, K.J.; Wu, F.; Peragallo, J.H.; Paul, J.; Roberts, L.; Ezura, Y.; Oldberg, A.; Birk, D.E.; Chakravarti, S. A Syndrome of Joint Laxity and Impaired Tendon Integrity in Lumican- and Fibromodulin-deficient Mice. J. Biol. Chem. 2002, 277, 35532–35540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikitovic, D.; Berdiaki, A.; Zafiropoulos, A.; Katonis, P.; Tsatsakis, A.; Karamanos, N.; Tzanakakis, G. Lumican expression is positively correlated with the differentiation and negatively with the growth of human osteosarcoma cells. FEBS J. 2007, 275, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Karamanou, K.; Franchi, M.; Vynios, D.; Brézillon, S. Epithelial-to-mesenchymal transition and invadopodia markers in breast cancer: Lumican a key regulator. Semin. Cancer Biol. 2019, 62, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Y.; Zuo, Y.; Ma, F.; Song, H. Lumican expression in gastric cancer and its association with biological behavior and prognosis. Oncol. Lett. 2017, 14, 5235–5240. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, X.; Hu, X.; Jiang, F.; Shen, Y.; Xu, R.; Wu, L.; Wei, P.; Shen, X. LUM Expression and Its Prognostic Significance in Gastric Cancer. Front. Oncol. 2020, 10, 605. [Google Scholar] [CrossRef]
- Yamauchi, N.; Kanke, Y.; Saito, K.; Okayama, H.; Yamada, S.; Nakajima, S.; Endo, E.; Kase, K.; Yamada, L.; Nakano, H.; et al. Stromal expression of cancer-associated fibroblast-related molecules, versican and lumican, is strongly associated with worse relapse-free and overall survival times in patients with esophageal squamous cell carcinoma. Oncol. Lett. 2021, 21, 445. [Google Scholar] [CrossRef]
- Seya, T.; Tanaka, N.; Shinji, S.; Yokoi, K.; Koizumi, M.; Teranishi, N.; Yamashita, K.; Tajiri, T.; Ishiwata, T.; Naito, Z. Lumican expression in advanced colorectal cancer with nodal metastasis correlates with poor prognosis. Oncol. Rep. 2006, 16, 1225–1230. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Dong, Q.; Lu, Y.; Dong, K.; Wang, R.; Liang, Z. Lumican inhibits immune escape and carcinogenic pathways in colorectal adenocarcinoma. Aging 2021, 13, 4388–4408. [Google Scholar] [CrossRef]
- De Wit, M.; Belt, E.J.T.; Diemen, P.M.D.-V.; Carvalho, B.; Coupé, V.M.H.; Stockmann, H.B.A.C.; Bril, H.; Beliën, J.A.M.; Fijneman, R.J.A.; Meijer, G.A. Lumican and Versican Are Associated with Good Outcome in Stage II and III Colon Cancer. Ann. Surg. Oncol. 2013, 20 (Suppl. 3), S348–S359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deyali, C.; Truty, M.A.; Kang, Y.; Chopin-Laly, X.; Zhang, R.; Roife, D.J.; Chatterjee, D.; Lin, E.; Thomas, R.M.; Wang, H.; et al. Extracellular Lumican Inhibits Pancreatic Cancer Cell Growth and Is Associated with Prolonged Survival after Surgery. Clin. Cancer Res. 2014, 20, 6529–6540. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Roife, D.; Lee, Y.; Lv, H.; Suzuki, R.; Ling, J.; Rios Perez, M.V.; Li, X.; Dai, B.; Pratt, M.; et al. Transforming Growth Factor-beta Limits Secretion of Lumican by Activated Stellate Cells within Primary Pancreatic Adenocarcinoma Tumors. Clin. Cancer Res. 2016, 22, 4934–4946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Brézillon, S.; Ventéo, L.; Ramont, L.; D’Onofrio, M.-F.; Perreau, C.; Pluot, M.; Maquart, F.-X.; Wegrowski, Y. Expression of lumican, a small leucine-rich proteoglycan with antitumour activity, in human malignant melanoma. Clin. Exp. Dermatol. 2007, 32, 405–416. [Google Scholar] [CrossRef]
- Nikitovic, D.; Chalkiadaki, G.; Berdiaki, A.; Aggelidakis, J.; Katonis, P.; Karamanos, N.K.; Tzanakakis, G.N. Lumican regulates osteosarcoma cell adhesion by modulating TGFbeta2 activity. Int. J. Biochem. Cell Biol. 2011, 43, 928–935. [Google Scholar] [CrossRef]
- Appunni, S.; Rubens, M.; Ramamoorthy, V.; Anand, V.; Khandelwal, M.; Saxena, A.; McGranaghan, P.; Odia, Y.; Kotecha, R.; Sharma, A. Lumican, pro-tumorigenic or anti-tumorigenic: A conundrum. Clin. Chim. Acta 2020, 514, 1–7. [Google Scholar] [CrossRef]
- Ho, T.H.; Serie, D.J.; Parasramka, M.; Cheville, J.C.; Bot, B.M.; Tan, W.; Wang, L.; Joseph, R.W.; Hilton, T.; Leibovich, B.C.; et al. Differential gene expression profiling of matched primary renal cell carcinoma and metastases reveals upregulation of extracellular matrix genes. Ann. Oncol. 2016, 28, 604–610. [Google Scholar] [CrossRef]
- Radwanska, A.; Litwin, M.; Nowak, D.; Baczynska, D.; Wegrowski, Y.; Maquart, F.-X.; Malicka-Blaszkiewicz, M. Overexpression of lumican affects the migration of human colon cancer cells through up-regulation of gelsolin and filamentous actin reorganization. Exp. Cell Res. 2012, 318, 2312–2323. [Google Scholar] [CrossRef]
- Salcher, S.; Spoden, G.; Huber, J.M.; Golderer, G.; Lindner, H.; Ausserlechner, M.J.; Kiechl-Kohlendorfer, U.; Geiger, K.; Obexer, P. Repaglinide Silences the FOXO3/Lumican Axis and Represses the Associated Metastatic Potential of Neuronal Cancer Cells. Cells 2019, 9, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhou, Q.; Yu, Z.; Wu, X.; Chen, X.; Li, J.; Li, C.; Yan, M.; Zhu, Z.; Liu, B.; et al. Cancer-associated fibroblast-derived Lumican promotes gastric cancer progression via the integrin beta1-FAK signaling pathway. Int. J. Cancer 2017, 141, 998–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, D.; Krawczenko, A.; Duś, D.; Malicka-Błaszkiewicz, M. Actin in human colon adenocarcinoma cells with different metastatic potential. Acta Biochim. Pol. 2002, 49, 823–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, Q.-M.; He, W.; Hou, G.-M.; Liang, Y.; Wang, G.; Li, C.-L.; Liao, B.; Liu, X.; Ye, Z.; Lu, J.-L.; et al. Interference of Lumican Regulates the Invasion and Migration of Liver Cancer Cells. Sichuan Da Xue Xue Bao Yi Xue Ban 2018, 49, 358–363. [Google Scholar] [PubMed]
- Li, X.; Kang, Y.; Roife, D.; Lee, Y.; Pratt, M.; Perez, M.R.; Dai, B.; Koay, E.J.; Fleming, J.B. Prolonged exposure to extracellular lumican restrains pancreatic adenocarcinoma growth. Oncogene 2017, 36, 5432–5438. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Lee, Y.; Kang, Y.; Dai, B.; Perez, M.R.; Pratt, M.; Koay, E.J.; Kim, M.; Brekken, R.A.; Fleming, J.B. Hypoxia-induced autophagy of stellate cells inhibits expression and secretion of lumican into microenvironment of pancreatic ductal adenocarcinoma. Cell Death Differ. 2018, 26, 382–393. [Google Scholar] [CrossRef] [Green Version]
- Jeanne, A.; Untereiner, V.; Perreau, C.; Proult, I.; Gobinet, C.; Boulagnon-Rombi, C.; Terryn, C.; Martiny, L.; Brézillon, S.; Dedieu, S. Lumican delays melanoma growth in mice and drives tumor molecular assembly as well as response to matrix-targeted TAX2 therapeutic peptide. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Brézillon, S.; Radwanska, A.; Zeltz, C.; Malkowski, A.; Ploton, D.; Bobichon, H.; Perreau, C.; Malicka-Blaszkiewicz, M.; Maquart, F.-X.; Wegrowski, Y. Lumican core protein inhibits melanoma cell migration via alterations of focal adhesion complexes. Cancer Lett. 2009, 283, 92–100. [Google Scholar] [CrossRef]
- Karamanou, K.; Franchi, M.; Proult, I.; Rivet, R.; Vynios, D.; Brézillon, S. Lumican Inhibits In Vivo Melanoma Metastasis by Altering Matrix-Effectors and Invadopodia Markers. Cells 2021, 10, 841. [Google Scholar] [CrossRef]
- Brézillon, S.; Untereiner, V.; Mohamed, H.; Ahallal, E.; Proult, I.; Nizet, P.; Boulagnon-Rombi, C.; Sockalingum, G.D. Label-Free Infrared Spectral Histology of Skin Tissue Part II: Impact of a Lumican-Derived Peptide on Melanoma Growth. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Yang, C.-T.; Hsu, P.-C.; Chow, S.-E. Downregulation of lumican enhanced mitotic defects and aneuploidy in lung cancer cells. Cell Cycle 2019, 19, 97–108. [Google Scholar] [CrossRef]
- Yang, C.-T.; Li, J.-M.; Chu, W.-K.; Chow, S.-E. Downregulation of lumican accelerates lung cancer cell invasion through p120 catenin. Cell Death Dis. 2018, 9, 414. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, M. Involvement of partial EMT in cancer progression. J. Biochem. 2018, 164, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Karamanou, K.; Franchi, M.; Piperigkou, Z.; Perreau, C.; Maquart, F.-X.; Vynios, D.H.; Brézillon, S. Lumican effectively regulates the estrogen receptors-associated functional properties of breast cancer cells, expression of matrix effectors and epithelial-to-mesenchymal transition. Sci. Rep. 2017, 7, srep45138. [Google Scholar] [CrossRef] [PubMed]
- Karamanou, K.; Franchi, M.; Onisto, M.; Passi, A.; Vynios, D.H.; Brézillon, S. Evaluation of lumican effects on morphology of invading breast cancer cells, expression of integrins and downstream signaling. FEBS J. 2020, 287, 4862–4880. [Google Scholar] [CrossRef]
- Leygue, E.; Snell, L.; Dotzlaw, H.; Hole, K.; Hiller-Hitchcock, T.; Roughley, P.J.; Watson, P.H.; Murphy, L.C. Expression of lumican in human breast carcinoma. Cancer Res. 1998, 58, 1348–1352. [Google Scholar] [PubMed]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Pietraszek-Gremplewicz, K.; Chatron-Colliet, A.; Brézillon, S.; Perreau, C.; Jakubiak-Augustyn, A.; Krotkiewski, H.; Maquart, F.-X.; Wegrowski, Y. Lumican: A new inhibitor of matrix metalloproteinase-14 activity. FEBS Lett. 2014, 588, 4319–4324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietraszek-Gremplewicz, K.; Karamanou, K.; Niang, A.; Dauchez, M.; Belloy, N.; Maquart, F.X.; Baud, S.; Brezillon, S. Small leucine-rich proteoglycans and matrix metalloproteinase-14: Key partners? Matrix Biol. 2019, 75, 271–285. [Google Scholar] [CrossRef]
- Karamanou, K.; Perrot, G.; Maquart, F.-X.; Brézillon, S. Lumican as a multivalent effector in wound healing. Adv. Drug Deliv. Rev. 2018, 129, 344–351. [Google Scholar] [CrossRef]
- Malinowski, M.; Pietraszek, K.; Perreau, C.; Boguslawski, M.; Decot, V.; Stoltz, J.-F.; Vallar, L.; Niewiarowska, J.; Cierniewski, C.; Maquart, F.-X.; et al. Effect of Lumican on the Migration of Human Mesenchymal Stem Cells and Endothelial Progenitor Cells: Involvement of Matrix Metalloproteinase-14. PLoS ONE 2012, 7, e50709. [Google Scholar] [CrossRef] [Green Version]
- Seomun, Y.; Kim, J.-T.; Joo, C.-K. MMP-14 mediated MMP-9 expression is involved in TGF-beta1-induced keratinocyte migration. J. Cell. Biochem. 2008, 104, 934–941. [Google Scholar] [CrossRef]
- Takino, T.; Suzuki, T.; Seiki, M. Isolation of Highly Migratory and Invasive Cells in Three-Dimensional Gels. Curr. Protoc. Cell Biol. 2020, 86, e103. [Google Scholar] [CrossRef]
- Long, W.; Yi, P.; Amazit, L.; LaMarca, H.L.; Ashcroft, F.; Kumar, R.; Mancini, M.A.; Tsai, S.Y.; Tsai, M.J.; O’Malley, B.W. SRC-3Delta4 mediates the interaction of EGFR with FAK to promote cell migration. Mol. Cell 2010, 37, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brézillon, S.; Pietraszek-Gremplewicz, K.; Maquart, F.-X.; Wegrowski, Y. Lumican effects in the control of tumour progression and their links with metalloproteinases and integrins. FEBS J. 2013, 280, 2369–2381. [Google Scholar] [CrossRef]
- Zeltz, C.; Brézillon, S.; Perreau, C.; Ramont, L.; Maquart, F.-X.; Wegrowski, Y. Lumcorin: A leucine-rich repeat 9-derived peptide from human lumican inhibiting melanoma cell migration. FEBS Lett. 2009, 583, 3027–3032. [Google Scholar] [CrossRef] [Green Version]
- Pietraszek, K.; Brézillon, S.; Perreau, C.; Malicka-Błaszkiewicz, M.; Maquart, F.-X.; Wegrowski, Y. Lumican—Derived Peptides Inhibit Melanoma Cell Growth and Migration. PLoS ONE 2013, 8, e76232. [Google Scholar] [CrossRef] [PubMed]
- Kuphal, S.; Palm, H.G.; Poser, I.; Bosserhoff, A. Snail-regulated genes in malignant melanoma. Melanoma Res. 2005, 15, 305–313. [Google Scholar] [CrossRef]
- Coulson-Thomas, V.J.; Coulson-Thomas, Y.M.; Gesteira, T.F.; de Paula, C.A.A.; Carneiro, C.R.; Ortiz, V.; Toma, L.; Kao, W.W.-Y.; Nader, H.B. Lumican expression, localization and antitumor activity in prostate cancer. Exp. Cell Res. 2013, 319, 967–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stasiak, M.; Boncela, J.; Perreau, C.; Karamanou, K.; Chatron-Colliet, A.; Proult, I.; Przygodzka, P.; Chakravarti, S.; Maquart, F.-X.; Kowalska, M.A.; et al. Lumican Inhibits SNAIL-Induced Melanoma Cell Migration Specifically by Blocking MMP-14 Activity. PLoS ONE 2016, 11, e0150226. [Google Scholar] [CrossRef] [Green Version]
- Zeltz, C.; Brezillon, S.; Kapyla, J.; Eble, J.A.; Bobichon, H.; Terryn, C.; Perreau, C.; Franz, C.M.; Heino, J.; Maquart, F.X.; et al. Lumican inhibits cell migration through alpha2beta1 integrin. Exp. Cell Res. 2010, 316, 2922–2931. [Google Scholar] [CrossRef]
- Zhou, K.; Ge, M. Effect of lumican gene over-expression on proliferation of lung adenocarcinoma cell A549 and its mechanism. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2013, 29, 492–495. [Google Scholar] [PubMed]
- Grzesiak, J.J.; Bouvet, M. The alpha2beta1 integrin mediates the malignant phenotype on type I collagen in pancreatic cancer cell lines. Br. J. Cancer 2006, 94, 1311–1319. [Google Scholar] [CrossRef]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Vij, N.; Roberts, L.; Joyce, S.; Chakravarti, S. Lumican suppresses cell proliferation and aids Fas–Fas ligand mediated apoptosis: Implications in the cornea. Exp. Eye Res. 2004, 78, 957–971. [Google Scholar] [CrossRef]
- Vuillermoz, B.; Khoruzhenko, A.; D’Onofrio, M.F.; Ramont, L.; Venteo, L.; Perreau, C.; Antonicelli, F.; Maquart, F.X.; Wegrowski, Y. The small leucine-rich proteoglycan lumican inhibits melanoma progression. Exp. Cell Res. 2004, 296, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.E.; Fulford, L.A.; Albig, A.R. Lumican Reduces Tumor Growth Via Induction of Fas-Mediated Endothelial Cell Apoptosis. Cancer Microenviron. 2010, 4, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Yoshimori, T.; Ohsumi, Y. The Role of Atg Proteins in Autophagosome Formation. Annu. Rev. Cell Dev. Biol. 2011, 27, 107–132. [Google Scholar] [CrossRef]
- Levy, J.M.M.; Towers, C.G.; Thorburn, A. Targeting autophagy in cancer. Nat. Rev. Cancer 2017, 17, 528–542. [Google Scholar] [CrossRef]
- Zhong, Z.; Sanchez-Lopez, E.; Karin, M. Autophagy, Inflammation, and Immunity: A Troika Governing Cancer and Its Treatment. Cell 2016, 166, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Li, P.; Peng, F.; Zhang, M.; Zhang, Y.; Liang, H.; Zhao, W.; Qi, L.; Wang, H.; Wang, C.; et al. Autophagy-related prognostic signature for breast cancer. Mol. Carcinog. 2015, 55, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Neill, T.; Kapoor, A.; Xie, C.; Buraschi, S.; Iozzo, R.V. A functional outside-in signaling network of proteoglycans and matrix molecules regulating autophagy. Matrix Biol. 2021, 100, 118–149. [Google Scholar] [CrossRef]
- Li, X.; Roife, D.; Kang, Y.; Dai, B.; Pratt, M.; Fleming, J.B. Extracellular lumican augments cytotoxicity of chemotherapy in pancreatic ductal adenocarcinoma cells via autophagy inhibition. Oncogene 2016, 35, 4881–4890. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, S.; Zeng, S.; Shen, H. The critical roles of activated stellate cells-mediated paracrine signaling, metabolism and onco-immunology in pancreatic ductal adenocarcinoma. Mol. Cancer 2018, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B.; Vijayalekshmi, R.V.; Sung, B. Targeting Inflammatory Pathways for Prevention and Therapy of Cancer: Short-Term Friend, Long-Term Foe. Clin. Cancer Res. 2009, 15, 425–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Avgustinova, A.; Iravani, M.; Robertson, D.; Fearns, A.; Gao, Q.; Klingbeil, P.; Hanby, A.M.; Speirs, V.; Sahai, E.; Calvo, F.; et al. Tumour cell-derived Wnt7a recruits and activates fibroblasts to promote tumour aggressiveness. Nat. Commun. 2016, 7, 10305. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, L.; Tredup, C.; Gubbiotti, M.A.; Iozzo, R.V. Proteoglycan neofunctions: Regulation of inflammation and autophagy in cancer biology. FEBS J. 2016, 284, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Zhang, W.; Yang, H.; Bi, J.; Xie, Y.; Cheng, B.; Wang, Y.; Chen, S. Celastrol Induces Necroptosis and Ameliorates Inflammation via Targeting Biglycan in Human Gastric Carcinoma. Int. J. Mol. Sci. 2019, 20, 5716. [Google Scholar] [CrossRef] [Green Version]
- Lohr, K.; Sardana, H.; Lee, S.; Wu, F.; Huso, D.L.; Hamad, A.R.; Chakravarti, S. Extracellular matrix protein lumican regulates inflammation in a mouse model of colitis. Inflamm. Bowel Dis. 2012, 18, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.-M.; Ma, L.; Jin, Y.-N.; Yu, Y.-Q. Lumican overexpression exacerbates lipopolysaccharide-induced renal injury in mice. Mol. Med. Rep. 2015, 12, 4089–4094. [Google Scholar] [CrossRef] [Green Version]
- Skjesol, A.; Yurchenko, M.; Bösl, K.; Gravastrand, C.; Nilsen, K.E.; Grøvdal, L.M.; Agliano, F.; Patane, F.; Lentini, G.; Kim, H.; et al. The TLR4 adaptor TRAM controls the phagocytosis of Gram-negative bacteria by interacting with the Rab11-family interacting protein 2. PLOS Pathog. 2019, 15, e1007684. [Google Scholar] [CrossRef]
- Jiang, Z.; Georgel, P.; Du, X.; Shamel, L.; Sovath, S.; Mudd, S.; Huber, M.; Kalis, C.; Keck, S.; Galanos, C.; et al. CD14 is required for MyD88-independent LPS signaling. Nat. Immunol. 2005, 6, 565–570. [Google Scholar] [CrossRef]
- Maiti, G.; Frikeche, J.; Lam, C.Y.-M.; Biswas, A.; Shinde, V.; Samanovic, M.; Kagan, J.C.; Mulligan, M.J.; Chakravarti, S. Matrix lumican endocytosed by immune cells controls receptor ligand trafficking to promote TLR4 and restrict TLR9 in sepsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2100999118. [Google Scholar] [CrossRef]
- Wu, F.; Vij, N.; Roberts, L.; López-Briones, S.; Joyce, S.; Chakravarti, S. A Novel Role of the Lumican Core Protein in Bacterial Lipopolysaccharide-induced Innate Immune Response. J. Biol. Chem. 2007, 282, 26409–26417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Komuro, Y.; Kiyomatsu, T.; Kanazawa, T.; Kazama, Y.; Tanaka, J.; Tanaka, T.; Yamamoto, Y.; Shirane, M.; Muto, T.; et al. Prediction of Sensitivity of Rectal Cancer Cells in Response to Preoperative Radiotherapy by DNA Microarray Analysis of Gene Expression Profiles. Cancer Res. 2006, 66, 3370–3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Liu, L.; Shu, Q.; Li, D.; Wang, R. Leukemia stem cells promote chemoresistance by inducing downregulation of lumican in mesenchymal stem cells. Oncol. Lett. 2019, 18, 4317–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dick, J.E.; Bhatia, M.; Gan, O.; Kapp, U.; Wang, J.C.Y. Assay of human stem cells by repopulation of NOD/SCID mice. STEM CELLS 1997, 15 (Suppl. 1), 199–207. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Cancer Type | Detected Expression (Protein/mRNA) | Level of Expression | Clinical Correlation | Ref. |
|---|---|---|---|---|
| Gastric cancer | Protein | Overexpressed in cancerous gastric tissues compared to normal tissues | Cancer dissemination to secondary sites and lymphatic metastasis | [48] |
| Gastric cancer | mRNA | Higher expression of lumican in the gastric cancer tissues than neighboring non-tumor tissues | Poor overall survival | [49] |
| Colon cancer | Protein | Overexpressed by cancer cells | Lymph node metastasis and a lower survival rate | [51] |
| Colon cancer | mRNA | Overexpressed | Poor prognosis | [52] |
| Adenoma to colon cancer transition | Protein | Increased expression during the transition process | Cancer stage | [44] |
| Colon cancer | Protein | Overexpressed | Positively correlated to a longer disease-specific and disease-free survival in stage II colon cancer patients and a more prolonged disease-specific survival in microsatellite-stable stage II colon cancer patients | [53] |
| Pancreatic ductal adenocarcinoma (PDAC) | Protein | Overexpressed | Associated with prolonged survival after surgery | [54] |
| Melanoma | Protein | Not expressed by tumor cells, expressed at peritumoral stroma | Negatively associated with melanoma growth | [57] |
| Cancer Type | Model | Alterations in Signaling Pathways | Effect on Cell Function | Ref. | |
|---|---|---|---|---|---|
| Tumorigenic action | Chondrosarcoma | HTB94 human cell line (in vitro) | IGF-I/IGF-IR/ERK1/2 | Cell growth | [32] |
| Osteosarcoma | Saos-2 human cell line (in vitro) | TGF-β2/Smad2 | Migration and adhesion to fibronectin substrate | [46,58] | |
| Gastric cancer | MKN45 human cell line, primary cell cultures, tissue biopsies (in vitro), and ice model (in vivo) | Integrin-β1/FAK | Cell growth, migration, and invasion | [63] | |
| Liver cancer | HepG2 and MHCC97H human cell lines (in vitro) | ERK1/JNK | Migration and invasion | [65] | |
| Neuroblastoma | SH-EP, SK-N-SH, and ZMR32 human cell lines (in vitro) | FoxO | Migration | [62] | |
| Anti-tumorigenic action | Lung cancer | A549, H460, H1975, H157, and H838 human cell lines (in vitro) | p120 catenin | Cadherin-mediated invasion | [73] |
| Pancreatic ductal adenocarcinoma (PDAC) | PANC-1 human cell line, PancO2 murine cell line, primary PDAC cells from PDX models (in vitro), and mice model (in vivo) | EGFR and TGF- β/p38/Smads | Cell growth | [66] | |
| Pancreatic ductal adenocarcinoma (PDAC) | PANC-1 human cell line, primary cell cultures (in vitro), and tissue biopsies from PDX model (ex vivo) | HIF-1a and AMPK | Cell growth | [67] | |
| Melanoma | A375 human cell line (in vitro) | Integrin-β1/FAK/vinculin | Migration | [69] | |
| Melanoma | B16F1 human cell line (in vitro) and mice model (in vivo) | Snail1 | Metastasis and invasion | [66] | |
| Breast cancer | MCF-7/c and MDA-MB-231 human cell lines (in vitro) | CD44/Hyaluronan synthase and Integrin-α1 and -β1/FAK/ERK1/2/MAPK 42/44/Akt | EMT metastasis | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giatagana, E.-M.; Berdiaki, A.; Tsatsakis, A.; Tzanakakis, G.N.; Nikitovic, D. Lumican in Carcinogenesis—Revisited. Biomolecules 2021, 11, 1319. https://doi.org/10.3390/biom11091319
Giatagana E-M, Berdiaki A, Tsatsakis A, Tzanakakis GN, Nikitovic D. Lumican in Carcinogenesis—Revisited. Biomolecules. 2021; 11(9):1319. https://doi.org/10.3390/biom11091319
Chicago/Turabian StyleGiatagana, Eirini-Maria, Aikaterini Berdiaki, Aristidis Tsatsakis, George N. Tzanakakis, and Dragana Nikitovic. 2021. "Lumican in Carcinogenesis—Revisited" Biomolecules 11, no. 9: 1319. https://doi.org/10.3390/biom11091319
APA StyleGiatagana, E.-M., Berdiaki, A., Tsatsakis, A., Tzanakakis, G. N., & Nikitovic, D. (2021). Lumican in Carcinogenesis—Revisited. Biomolecules, 11(9), 1319. https://doi.org/10.3390/biom11091319