
Therapeutic Potential of Curcumin as an Antimycobacterial Agent


Abstract
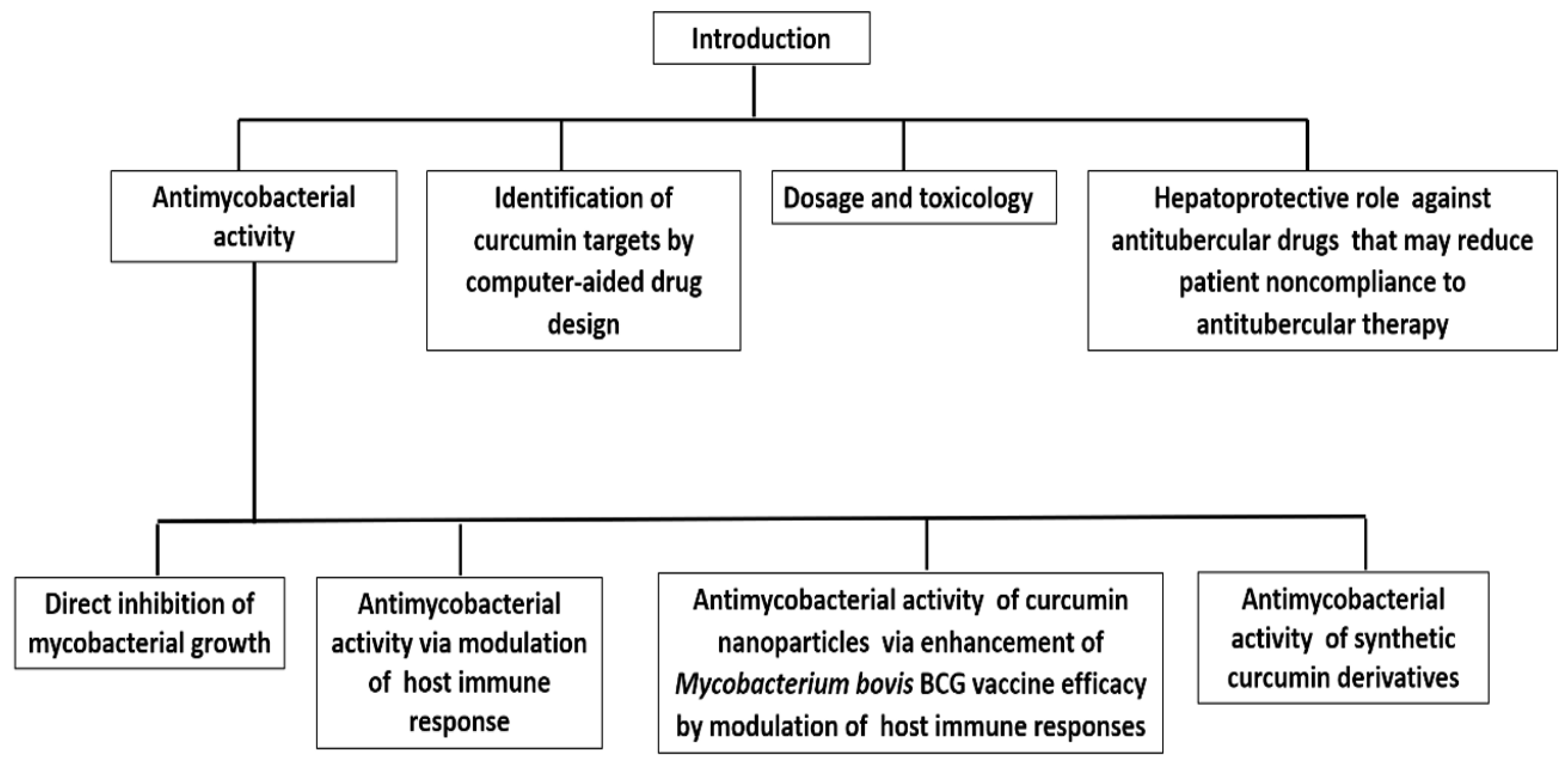
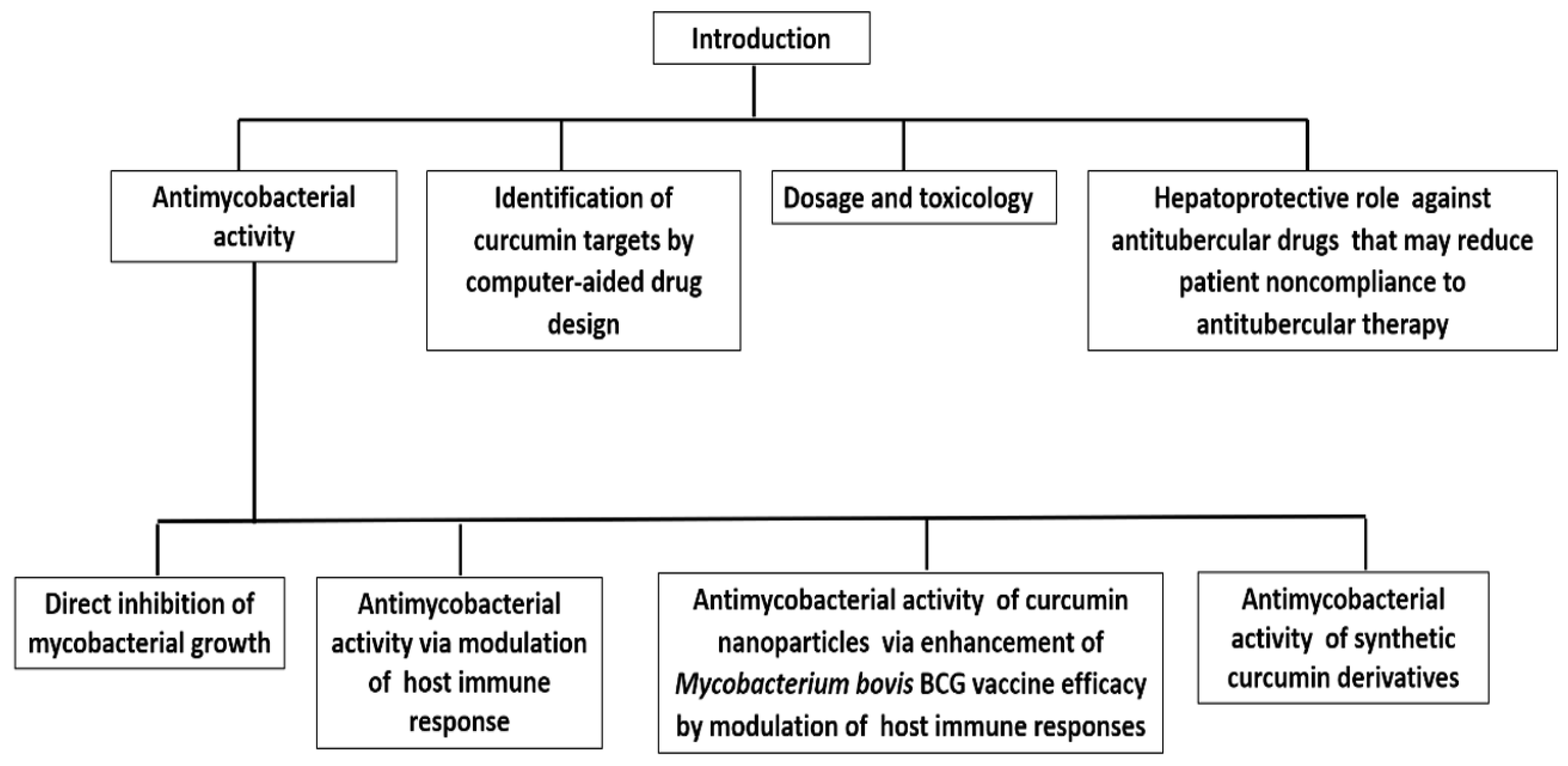
:1. Introduction
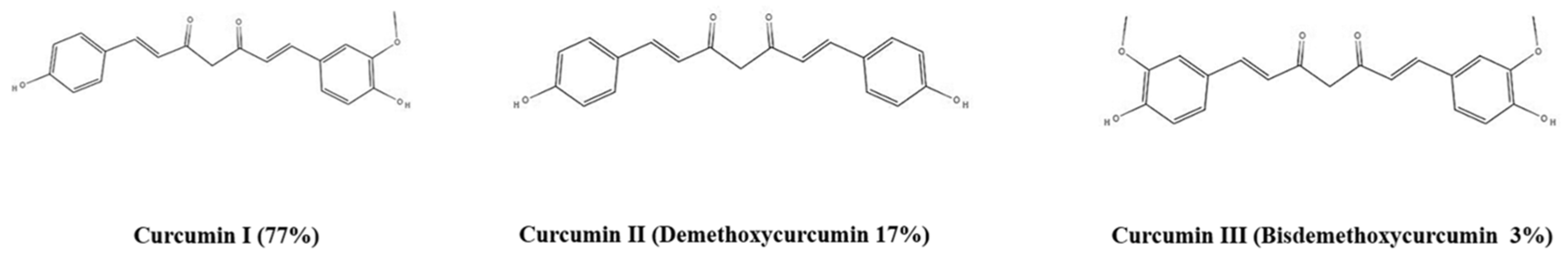
2. Physical, Chemical, and Biochemical Properties of Curcumin
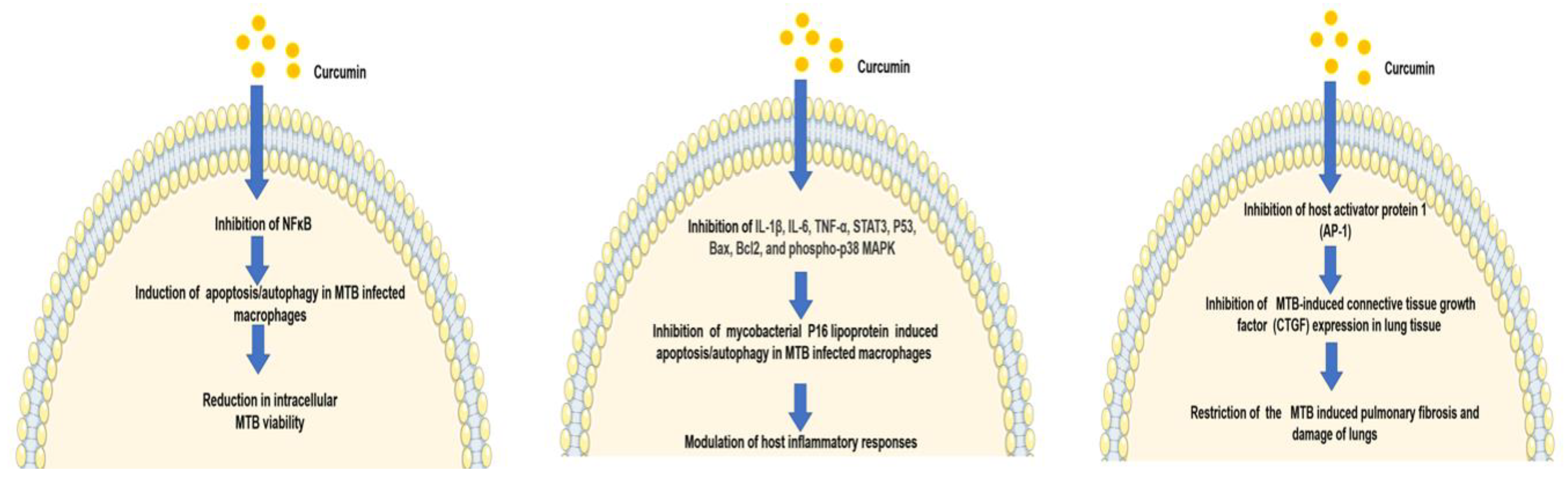
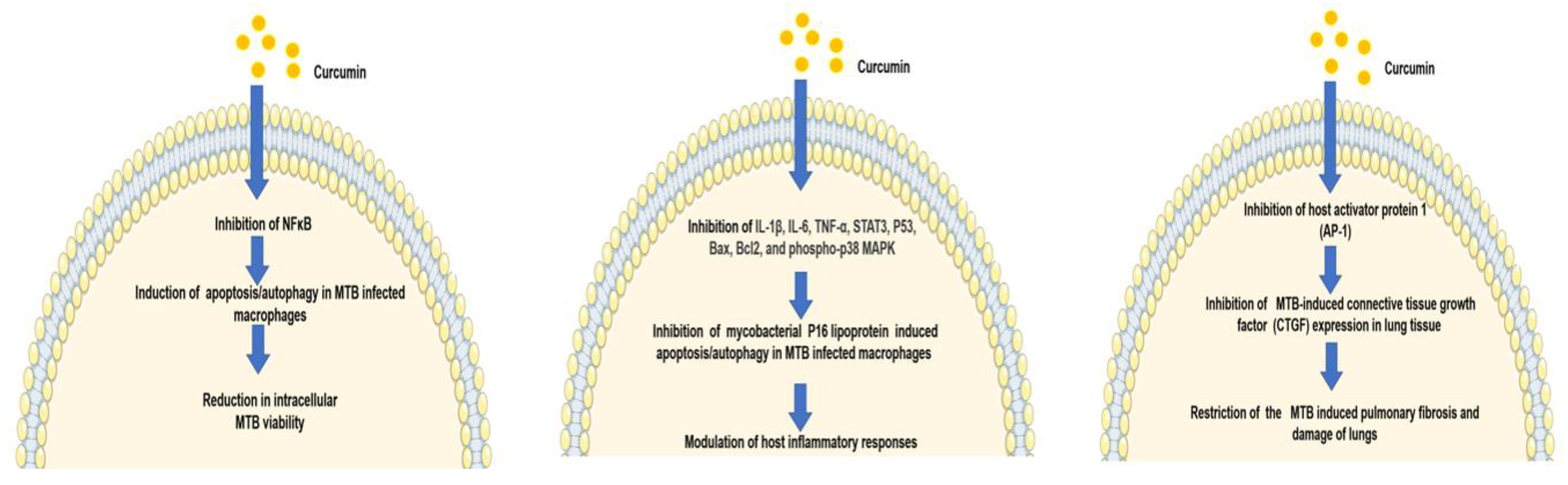
2.1. Curcumin as an Antimycobacterial Agent
2.2. Curcumin Exhibits Antitubercular Activity via Modulation of the Host Immune Response
2.3. Curcumin Nanoparticles Enhance Mycobacterium bovis BCG Vaccine Efficacy by Modulating Host Immune Responses
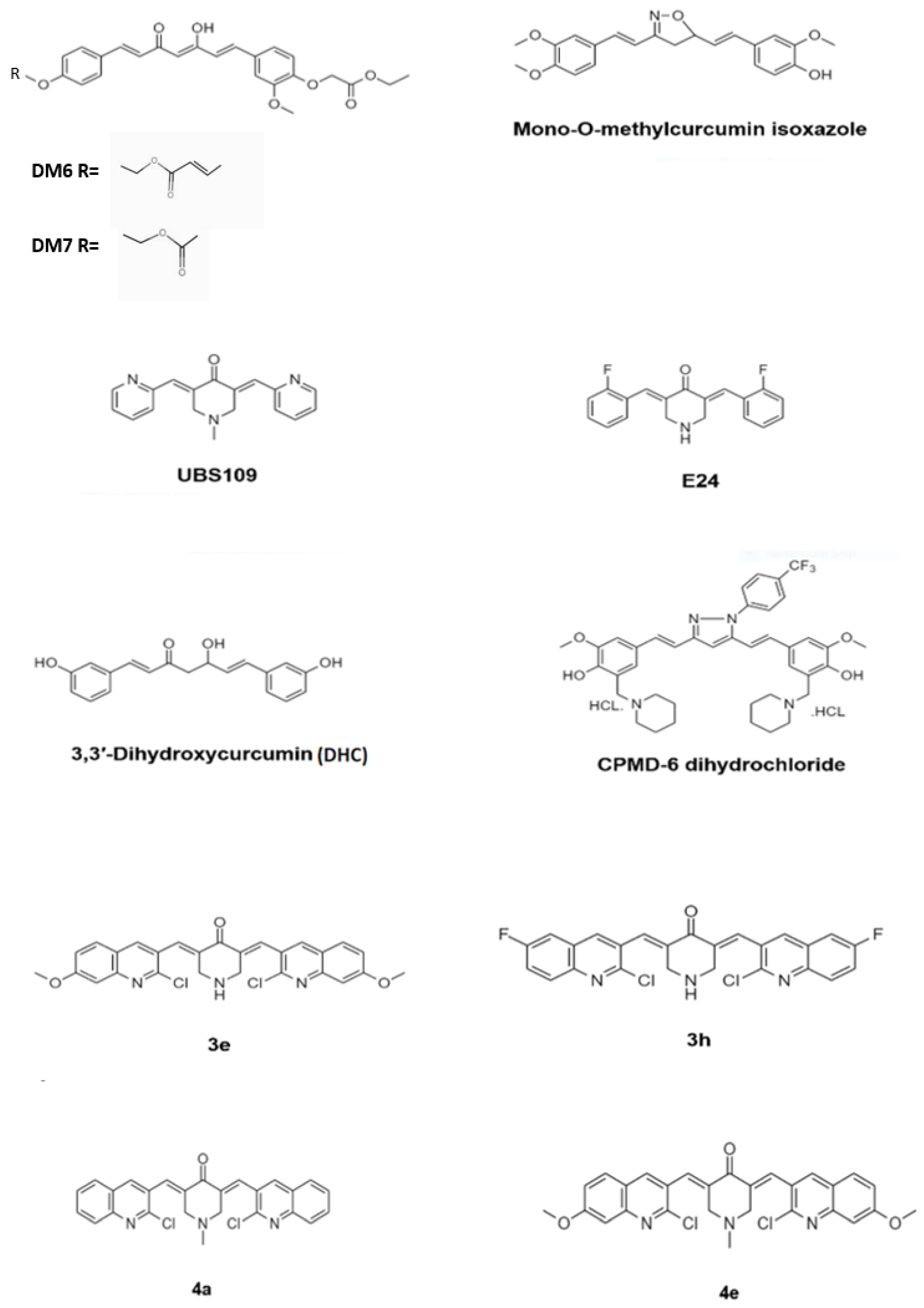
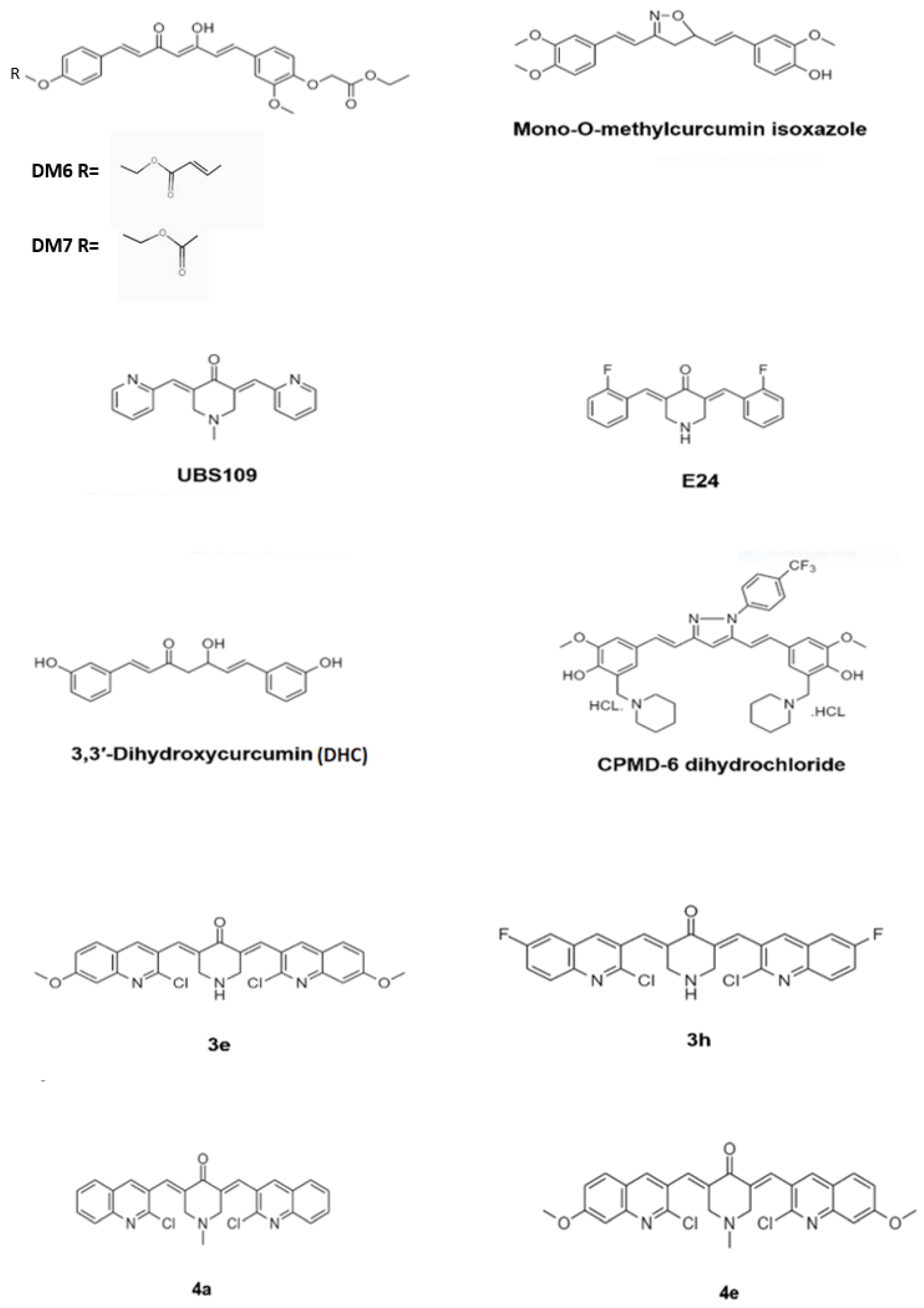
3. Antimycobacterial Activity of Curcumin Synthetic Derivatives
4. Identification of Curcumin Targets in Mycobacterium by Computer-Aided Drug Design
5. The Dosage and Toxicology of Curcumin
6. Hepatoprotective Role of Curcumin against Anti-TB Chemotherapy (ATT)
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Alipoor, S.D.; Adcock, I.M.; Tabarsi, P.; Folkerts, G.; Mortaz, E. MiRNAs in tuberculosis: Their decisive role in the fate of TB. Eur. J. Pharmacol. 2020, 886, 173529. [Google Scholar] [CrossRef]
- Adnan, M.; Ali, S.; Sheikh, K.; Amber, R. Review on antibacterial activity of Himalayan medicinal plants traditionally used to treat pneumonia and tuberculosis. J. Pharm. Pharmacol. 2019, 71, 1599–1625. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.B.; Odingo, J.; Bailey, M.A.; Sunde, B.; Korkegian, A.; O’Malley, T.; Ovechkina, Y.; Ioerger, T.R.; Sacchettini, J.C.; Young, K.; et al. Identification of cyclic hexapeptides natural products with inhibitory potency against Mycobacterium tuberculosis. BMC Res. Notes 2018, 11, 416. [Google Scholar] [CrossRef] [Green Version]
- Pires, C.T.; Scodro, R.B.; Cortez, D.A.; Brenzan, M.A.; Siqueira, V.L.; Caleffi-Ferracioli, K.R.; Vieira, L.C.; Monteiro, J.L.; Corrêa, A.G.; Cardoso, R.F. Structure–activity relationship of natural and synthetic coumarin derivatives against Mycobacterium tuberculosis. Future Med. Chem. 2020, 12, 1533–1546. [Google Scholar] [CrossRef] [PubMed]
- Nguta, J.M.; Appiah-Opong, R.; Nyarko, A.K.; Yeboah-Manu, D.; Addo, P.G. Current perspectives in drug discovery against tuberculosis from natural products. Int. J. Mycobacteriol. 2015, 4, 165–183. [Google Scholar] [CrossRef] [Green Version]
- Tizabi, Y.; Hurley, L.L.; Qualls, Z.; Akinfiresoye, L. Relevance of the anti-inflammatory properties of curcumin in neurodegenerative diseases and depression. Molecules 2014, 19, 20864–20879. [Google Scholar] [CrossRef] [PubMed]
- Dragos, D.; Gilca, M.; Gaman, L.; Vlad, A.; Iosif, L.; Stoian, I.; Lupescu, O. Phytomedicine in joint disorders. Nutrients 2017, 9, 70. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Zarrabi, A.; Hashemi, F.; Moghadam, E.R.; Hashemi, F.; Entezari, M.; Hushmandi, K.; Mohammadinejad, R.; Najafi, M. Curcumin in cancer therapy: A novel adjunct for combination chemotherapy with paclitaxel and alleviation of its adverse effects. Life Sci. 2020, 256, 117984. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yue, Y.; Zheng, X.; Zhang, K.; Chen, S.; Du, Z. Curcumin, inflammation, and chronic diseases: How are they linked? Molecules 2015, 20, 9183–9213. [Google Scholar] [CrossRef]
- Vera-Ramirez, L.; Pérez-Lopez, P.; Varela-Lopez, A.; Ramirez-Tortosa, M.; Battino, M.; Quiles, J.L. Curcumin and liver disease. Biofactors 2013, 39, 88–100. [Google Scholar] [CrossRef]
- Panahi, Y.; Hosseini, M.S.; Khalili, N.; Naimi, E.; Simental-Mendía, L.E.; Majeed, M.; Sahebkar, A. Effects of curcumin on serum cytokine concentrations in subjects with metabolic syndrome: A post-hoc analysis of a randomized controlled trial. Biomed. Pharmacother. 2016, 82, 578–582. [Google Scholar] [CrossRef]
- Wang, Q.; Ye, C.; Sun, S.; Li, R.; Shi, X.; Wang, S.; Zeng, X.; Kuang, N.; Liu, Y.; Shi, Q. Curcumin attenuates collagen-induced rat arthritis via anti-inflammatory and apoptotic effects. Int. Immunopharmacol. 2019, 72, 292–300. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; Sánchez, C.R.; Rodriguez, J.; Cani, P.D.; Bindels, L.B.; Delzenne, N.M. Prebiotic Effect of Berberine and Curcumin Is Associated with the Improvement of Obesity in Mice. Nutrients 2021, 13, 1436. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, pharmaceutical, nutraceutical, and analytical aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef] [Green Version]
- Mangwani, N.; Singh, P.K.; Kumar, V. Medicinal plants: Adjunct treatment to tuberculosis chemotherapy to prevent hepatic damage. J. Ayurveda Integr. Med. 2020, 11, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Alsarhan, A.; Sultana, N.; Al-Khatib, A.; Kadir, M.R.A. Review on some Malaysian traditional medicinal plants with therapeutic properties. J. Basic Appl. 2014, 10, 149–159. [Google Scholar]
- Deb, B.C.; Chakraborty, S. Evaluation of genetic variability and characterization of some elite turmeric genotypes in Terai Region in India. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 2357–2366. [Google Scholar] [CrossRef]
- Soleimani, V.; Sahebkar, A.; Hosseinzadeh, H. Turmeric (Curcuma longa) and its major constituent (curcumin) as nontoxic and safe substances. Phytother. Res. 2018, 32, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Eke-Okoro, U.J.; Raffa, R.B.; Pergolizzi, J.V., Jr.; Breve, F.; Taylor, R., Jr.; NEMA Research Group. Curcumin in turmeric: Basic and clinical evidence for a potential role in analgesia. J. Clin. Pharm. Ther. 2018, 43, 460–466. [Google Scholar] [CrossRef] [Green Version]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A review of curcumin and its derivatives as anticancer agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahams, S.; Haylett, W.L.; Johnson, G.; Carr, J.A.; Bardien, S. Antioxidant effects of curcumin in models of neurodegeneration, aging, oxidative and nitrosative stress: A review. Neuroscience 2019, 406, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Pivari, F.; Mingione, A.; Brasacchio, C.; Soldati, L. Curcumin and type 2 diabetes mellitus: Prevention and treatment. Nutrients 2019, 11, 1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.S.; Acharya, A.; Ray, R.S.; Agrawal, R.; Raghuwanshi, R.; Jain, P. Cellular and molecular mechanisms of curcumin in prevention and treatment of disease. Crit. Rev. Food. Sci. Nutr. 2020, 60, 887–939. [Google Scholar] [CrossRef] [PubMed]
- Marini, E.; Di Giulio, M.; Magi, G.; Di Lodovico, S.; Cimarelli, M.E.; Brenciani, A.; Nostro, A.; Cellini, L.; Facinelli, B. Curcumin, an antibiotic resistance breaker against a multiresistant clinical isolate of Mycobacterium abscessus. Phytother. Res. 2018, 32, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.; Oberley-Deegan, R.E.; Bai, A.; Ovrutsky, A.R.; Kinney, W.H.; Weaver, M.; Zhang, G.; Honda, J.R.; Chan, E.D. Curcumin enhances human macrophage control of Mycobacterium tuberculosis infection. Respirology 2016, 21, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Shariq, M.; Quadir, N.; Sharma, N.; Singh, J.; Sheikh, J.A.; Khubaib, M.; Hasnain, S.E.; Ehtesham, N.Z. Mycobacterium tuberculosis RipA Dampens TLR4-Mediated Host Protective Response Using a Multi-Pronged Approach Involving Autophagy, Apoptosis, Metabolic Repurposing, and Immune Modulation. Front. Immunol. 2021, 12, 434. [Google Scholar] [CrossRef]
- Arnett, E.; Weaver, A.M.; Woodyard, K.C.; Montoya, M.J.; Li, M.; Hoang, K.V.; Hayhurst, A.; Azad, A.K.; Schlesinger, L.S. PPARγ is critical for Mycobacterium tuberculosis induction of Mcl-1 and limitation of human macrophage apoptosis. PLoS Pathog. 2018, 14, e1007100. [Google Scholar] [CrossRef]
- Bah, A.; Sanicas, M.; Nigou, J.; Guilhot, C.; Astarie-Dequeker, C.; Vergne, I. The lipid virulence factors of Mycobacterium tuberculosis exert multilayered control over autophagy-related pathways in infected human macrophages. Cells 2020, 9, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.Y.; Wang, H.L.; Huang, J.; Shi, G.C.; Wan, Y.G.; Wang, J.X.; Xi, X.E. Curcumin inhibits 19-kDa lipoprotein of Mycobacterium tuberculosis induced macrophage apoptosis via regulation of the JNK pathway. Biochem. Bioph. Res. Commun. 2014, 446, 626–632. [Google Scholar]
- Liu, L.; Liu, J.; Niu, G.; Wu, Q.; Li, W.; Zhou, Y. The effects of curcumin on the 19 000 Mycobacterium tuberculosis protein-induced inflammatory and apoptotic reaction and the expression of p38 mitogen-activated protein kinases in WBC264-9C macrophages. Chin. J. Tuberc. Respir. Dis. 2014, 37, 421–426. [Google Scholar]
- Marzouki, S.; Bini Dhouib, I.; Benabdessalem, C.; Rekik, R.; Doghri, R.; Maroueni, A.; Bellasfar, Z.; Fazaa, S.; Bettaieb, J.; Barbouche, M.R.; et al. Specific immune responses in mice following subchronic exposure to acetamiprid. Life Sci. 2017, 188, 10–16. [Google Scholar] [CrossRef]
- Erokhina, M.V.; Lepekha, L.N.; Voronezhskaya, E.E.; Nezlin, L.P.; Avdienko, V.G.; Ergeshov, A.E. Application of Laser Scanning Confocal Microscopy for the Visualization of M. tuberculosis in Lung Tissue Samples with Weak Ziehl–Neelsen Staining. J. Clin. Med. 2019, 8, 1185. [Google Scholar] [CrossRef] [Green Version]
- Urbanowski, M.E.; Ordonez, A.A.; Ruiz-Bedoya, C.A.; Jain, S.K.; Bishai, W.R. Cavitary tuberculosis: The gateway of disease transmission. Lancet Infect. Dis. 2020, 20, e117–e128. [Google Scholar] [CrossRef]
- Odashima, K.; Kagiyama, N.; Kanauchi, T.; Ishiguro, T.; Takayanagi, N. Incidence and etiology of chronic pulmonary infections in patients with idiopathic pulmonary fibrosis. PLoS ONE 2020, 15, e0230746. [Google Scholar] [CrossRef]
- Lee, H.; Hua, H.; Wang, C.; Yu, M.; Chen, B.; Lin, A.C. Mycobacterium tuberculosis induces connective tissue growth factor expression through the TLR2-JNK-AP-1 pathway in human lung fibroblasts. FASEB J. 2019, 33, 12554–12564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machlaurin, A.; Dolk, F.C.K.; Setiawan, D.; Werf, T.S.; Postma, M.J. Cost-Effectiveness Analysis of BCG Vaccination against Tuberculosis in Indonesia: A Model-Based Study. Vaccines 2020, 8, 707. [Google Scholar] [CrossRef] [PubMed]
- Whitlow, E.; Mustafa, A.S.; Hanif, S.N.M. An Overview of the Development of New Vaccines for Tuberculosis. Vaccines 2020, 8, 586. [Google Scholar] [CrossRef]
- Ahmad, S.; Bhattacharya, D.; Kar, S.; Ranganathan, A.; Van Kaer, L.; Das, G. Curcumin Nanoparticles Enhance Mycobacterium bovis BCG Vaccine Efficacy by Modulating Host Immune Responses. Infect. Immun. 2019, 87, e00291-19. [Google Scholar] [CrossRef] [Green Version]
- Tousif, S.; Singh, D.K.; Mukherjee, S.; Ahmad, S.; Arya, R.; Nanda, R.; Ranganathan, A.; Bhattacharyya, M.; Van Kaer, L.; Kar, S.K.; et al. Nanoparticle-Formulated Curcumin Prevents Posttherapeutic Disease Reactivation and Reinfection with Mycobacterium tuberculosis following Isoniazid Therapy. Front. Immunol. 2017, 8, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahagirdar, P.S.; Gupta, P.K.; Kulkarni, S.P.; Devarajan, P.V. Intramacrophage Delivery of Dual Drug Loaded Nanoparticles for Effective Clearance of Mycobacterium tuberculosis. J. Pharm. Sci. 2020, 109, 2262–2270. [Google Scholar] [CrossRef] [PubMed]
- Heger, M.; van Golen, R.F.; Broekgaarden, M.; Michel, M.C. The Molecular Basis for the Pharmacokinetics and Pharmacodynamics of Curcumin and Its Metabolites in Relation to Cancer. Pharmacol. Rev. 2013, 66, 222–307. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.A.; Gescher, A.J.; Steward, W.P. Curcumin: The story so far. Eur. J. Cancer 2005, 41, 1955–1968. [Google Scholar] [CrossRef] [PubMed]
- Nasir Abbas Bukhari, S.; Franzblau, S.G.; Jantan, I.; Jasamai, M. Current prospects of synthetic curcumin anlogues and chalcone derivatives against Mycobacterium tuberculosis. Med. Chem. 2013, 9, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Yadav, P.; Karaulia, P.; Singh, V.K.; Gupta, P.; Puttrevu, S.K.; Chauhan, S.; Bhatta, R.S.; Tadigoppula, N.; Gupta, U.D.; et al. Biological evaluation of novel curcumin-pyrazole-mannich derivative active against drug-resistant Mycobacterium tuberculosis. Future Microbiol. 2017, 12, 1349–1362. [Google Scholar] [CrossRef]
- Agrawal, D.K.; Saikia, D.; Tiwari, R.; Ojha, S.; Shanker, K.; Kumar, J.K.; Gupta, A.K.; Tandon, S.; Negi, A.S.; Khanuja, S.P. Demethoxycurcumin and its Semisynthetic Analogues as Antitubercular Agents. Planta Med. 2008, 74, 1828–1831. [Google Scholar] [CrossRef] [Green Version]
- Changtam, C.; Hongmanee, P.; Suksamrarn, A. Isoxazole anlogues of curcuminoids with highly potent multidrug-resistant antimycobacterial activity. Eur. J. Med. Chem. 2010, 45, 4446–4457. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, P.R.; Reeves, A.Z.; Powell, K.R.; Napier, R.J.; Swimm, A.I.; Sun, A.; Giesler, K.; Bommarius, B.; Shinnick, T.M.; Snyder, J.P.; et al. Monocarbonyl anlogues of curcumin inhibit growth of antibiotic sensitive and resistant strains of Mycobacterium tuberculosis. Eur. J. Med. Chem. 2015, 92, 693–699. [Google Scholar] [CrossRef] [Green Version]
- Polaquini, C.R.; Morão, L.G.; Nazaré, A.C.; Torrezan, G.S.; Dilarri, G.; Cavalca, L.B.; Campos, D.L.; Silva, I.C.; Pereira, J.A.; Scheffers, D.; et al. Antibacterial activity of 3,3′-dihydroxycurcumin (DHC) is associated with membrane perturbation. Bioorg. Chem. 2019, 90, 103031. [Google Scholar] [CrossRef]
- Subhedar, D.D.; Shaikh, M.H.; Nawale, L.; Sarkar, D.; Khedkar, V.M.; Shingate, B.B. Quinolidene based monocarbonyl curcumin analogues as promising antimycobacterial agents: Synthesis and molecular docking study. Bioorg. Med. Chem. Lett. 2017, 27, 922–928. [Google Scholar] [CrossRef]
- Muegge, I.; Bergner, A.; Kriegl, J.M. Computer-aided drug design at Boehringer Ingelheim. J. Comput. Aided. 2017, 31, 275–285. [Google Scholar] [CrossRef]
- Bruch, E.M.; Petrella, S.; Bellinzoni, M. Structure-Based Drug Design for Tuberculosis: Challenges Still Ahead. Appl. Sci. 2020, 10, 4248. [Google Scholar] [CrossRef]
- Aanandhi, M.V.; Bhattacherjee, D.; George, P.S.G.; Ray, A. Natural polyphenols down-regulate universal stress protein in Mycobacterium tuberculosis: An in-silico approach. J. Adv. Pharm. Technol. Res. 2014, 5, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Somvanshi, P.; Bhardwaj, T.; Mandal, R.K.; Dar, S.A.; Wahid, M.; Jawed, A.; Lohani, M.; Khan, M.; Areeshi, M.Y.; et al. Aspartate-β-semialdeyhyde dehydrogenase as a potential therapeutic target of Mycobacterium tuberculosis H37Rv: Evidence from in silico elementary mode analysis of biological network model. J. Cell. Biochem. 2018, 119, 2832–2842. [Google Scholar] [CrossRef] [PubMed]
- Dube, D.; Kukshal, V.; Srivastava, S.K.; Tripathi, R.P.; Ramachandran, R. NAD+-dependent DNA ligase (Rv3014c) from M. tuberculosis: Strategies for inhibitor design. Med. Chem. Res. 2007, 17, 189–198. [Google Scholar] [CrossRef]
- Takeuchi, T.; Ishidoh, T.; Iijima, H.; Kuriyama, I.; Shimazaki, N.; Koiwai, O.; Kuramochi, K.; Kobayashi, S.; Sugawara, F.; Sakaguchi, K.; et al. Structural relationship of curcumin derivatives binding to the BRCT domain of human DNA polymerase λ. Genes Cells 2006, 11, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Kocaadam, B.; Şanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2015, 57, 2889–2895. [Google Scholar] [CrossRef] [PubMed]
- Lao, C.D.; Ruffin, M.T.; Normolle, D.; Heath, D.D.; Murray, S.I.; Bailey, J.M.; Boggs, M.E.; Crowell, J.; Rock, C.L.; Brenner, D.E. Dose escalation of a curcuminoid formulation. BMC Complem. Altern. Med. 2006, 6, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotwal, P.; Dogra, A.; Sharma, A.; Bhatt, S.; Gour, A.; Sharma, S.; Wazir, P.; Singh, P.P.; Kumar, A.; Nandi, U. Effect of Natural Phenolics on Pharmacokinetic Modulation of Bedaquiline in Rat to Assess the Likelihood of Potential Food–Drug Interaction. J. Agric. Food Chem. 2020, 68, 1257–1265. [Google Scholar] [CrossRef]
- Ramappa, V.; Aithal, G.P. Hepatotoxicity Related to Anti-tuberculosis Drugs: Mechanisms and Management. J. Clin. Exp. Hepatol. 2013, 3, 37–49. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Guo, Y.; Deng, Y.; Li, C.; Zuo, C.; Peng, W. Involvement of protoporphyrin IX accumulation in the pathogenesis of isoniazid/rifampicin-induced liver injury: The prevention of curcumin. Xenobiotica 2017, 47, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Coelho, T.S.; Perdigão, J.; Pereira, C.; Couto, I.; Portugal, I.; Maschmann, R.D.A.; Ramos, D.F.; von Groll, A.; Rossetti, M.L.; et al. Interplay between mutations and efflux in drug resistant clinical isolates of Mycobacterium tuberculosis. Front. Microbiol. 2017, 8, 711. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Mycobacterial Strains | MIC (μg/mL) | Reference |
|---|---|---|---|
| Curcumin | MTB H37Rv | 16 | [45] |
| Demethoxycurcumin (DM) | MTB H37Rv | 200 | [46] |
| DM6 | MTB H37Rv | 7.8 | |
| DM7 | MTB H37Rv | 125 | |
| Mono-O-methylcurcumin- isoxazole | MTB H37Ra | 0.09 | [47] |
| a INH, RIF, STM-resistant MTB (clinical isolate M3) | 0.195 | ||
| b INH, RIF, EMB, STM-resistant MTB (clinical isolate M4) | 1.56 | ||
| INH, RIF, STM-resistant MTB (clinical isolate M5) | 3.125 | ||
| INH, RIF-resistant MTB (clinical isolate M6) | 0.39 | ||
| INH, RIF, EMB, STM-resistant MTB (clinical isolate M8) | 3.125 | ||
| INH, RIF-resistant MTB (clinical isolate M11) | 3.125 | ||
| INH, RIF, EMB-resistant MTB (clinical isolate M16) | 1.56 | ||
| INH, RIF, EMB, STM-resistant MTB (clinical isolate M21) | 3.125 | ||
| INH, RIF-resistant MTB (clinical isolate M22) | 0.39 | ||
| INH, RIF-resistant MTB (clinical isolate M27) | 0.195 | ||
| c INH, RIF, STM, OFX, CIP-resistant MTB (clinical isolate M46) | 1.56 | ||
| INH, RIF, STM, OFX, CIP-resistant MTB (clinical isolate M48) | 1.56 | ||
| INH, RIF, STM, OFX, CIP-resistant MTB (clinical isolate M53) | 3.125 | ||
| UBS-109 | M. marinum | 10 mM * | [48] |
| MTB H37Rv | ~10 μM * | ||
| EF-24 | MTB Beijing F2 | 20 μM * | |
| M. marinum | 25 mM * | ||
| CPMD-6- dihydrochloride | MTB H37R | 2 | [45] |
| INH-resistant MTB ATCC 35822 | 2 | ||
| RIF-resistant MTB ATCC 35838 | 2 | ||
| STM-resistant MTB ATCC 35820 | 2 | ||
| ETB-resistant MTB ATCC 35837 | 2 | ||
| M. fortuitum ATCC 6841 | 16 | ||
| M.abscessus ATCC 19977 | 16 | ||
| 3,3′-Dihydroxycurcumin | MTB | 156 | [49] |
| Quinolidene based monocarbonyl curcumin analogue 3e | MTB | >30 # | [50] |
| M. bovis BCG | 2.7 # | ||
| Quinolidene based monocarbonyl curcumin analogue 3h | MTB | >30 # | |
| M. bovis BCG | 9.2 # | ||
| Quinolidene based monocarbonyl curcumin analogue 4a | MTB | 26.5 # | |
| M. bovis BCG | 7.3 # | ||
| Quinolidene based monocarbonyl curcumin analogue 4e | MTB | 7.8 # | |
| M. bovis BCG | 9.4 # |
| Compound | Predicted Mycobacterial Target | Reference |
|---|---|---|
| Curcumin | Universal stress protein (USP) | [53] |
| Aspartate-β-semialdehyde dehydrogenase (ASD) | [54] | |
| Dihydrodipicolinate reductase | ||
| Monoacetylcurcumin | M. tuberculosis NAD+-dependent DNA ligases (MtuLigA) | [55] |
| BRCT domain-containing DNA polymerase λ | [56] | |
| Quinolidene based monocarbonyl curcumin analogues 3e, 3h, 4a and 4e | Pantothenate synthetase (MTB PS) | [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barua, N.; Buragohain, A.K. Therapeutic Potential of Curcumin as an Antimycobacterial Agent. Biomolecules 2021, 11, 1278. https://doi.org/10.3390/biom11091278
Barua N, Buragohain AK. Therapeutic Potential of Curcumin as an Antimycobacterial Agent. Biomolecules. 2021; 11(9):1278. https://doi.org/10.3390/biom11091278
Chicago/Turabian StyleBarua, Nilakshi, and Alak Kumar Buragohain. 2021. "Therapeutic Potential of Curcumin as an Antimycobacterial Agent" Biomolecules 11, no. 9: 1278. https://doi.org/10.3390/biom11091278
APA StyleBarua, N., & Buragohain, A. K. (2021). Therapeutic Potential of Curcumin as an Antimycobacterial Agent. Biomolecules, 11(9), 1278. https://doi.org/10.3390/biom11091278